An Evaluation of Systematic Versus Strategically-Placed Camera Traps for Monitoring Feral Cats in New Zealand


Simple Summary
Abstract
1. Introduction
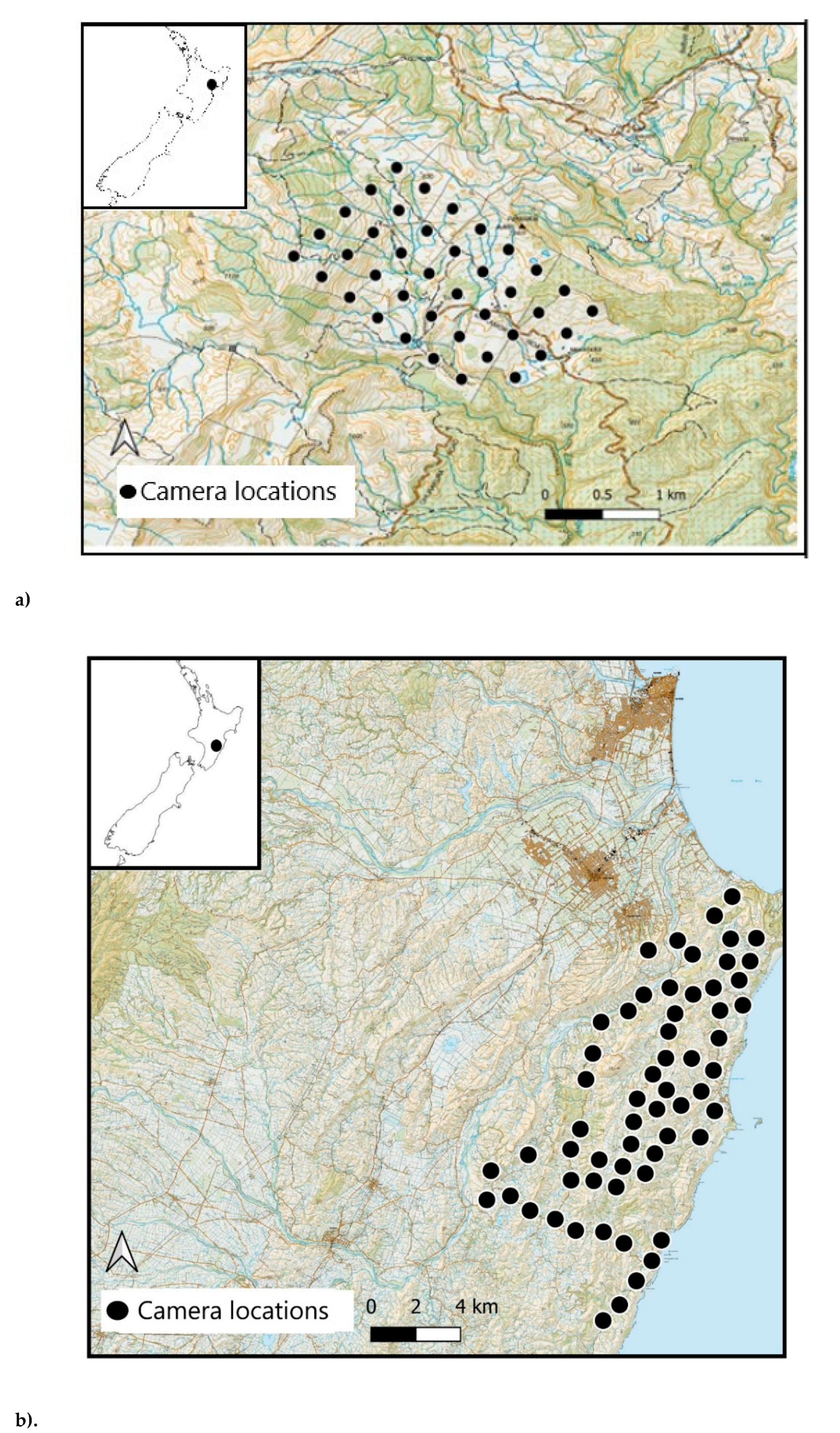
1.1. Camera Trap Deployment
1.2. Feral Cats as a Target Species
2. Materials and Methods
Occupancy Modelling
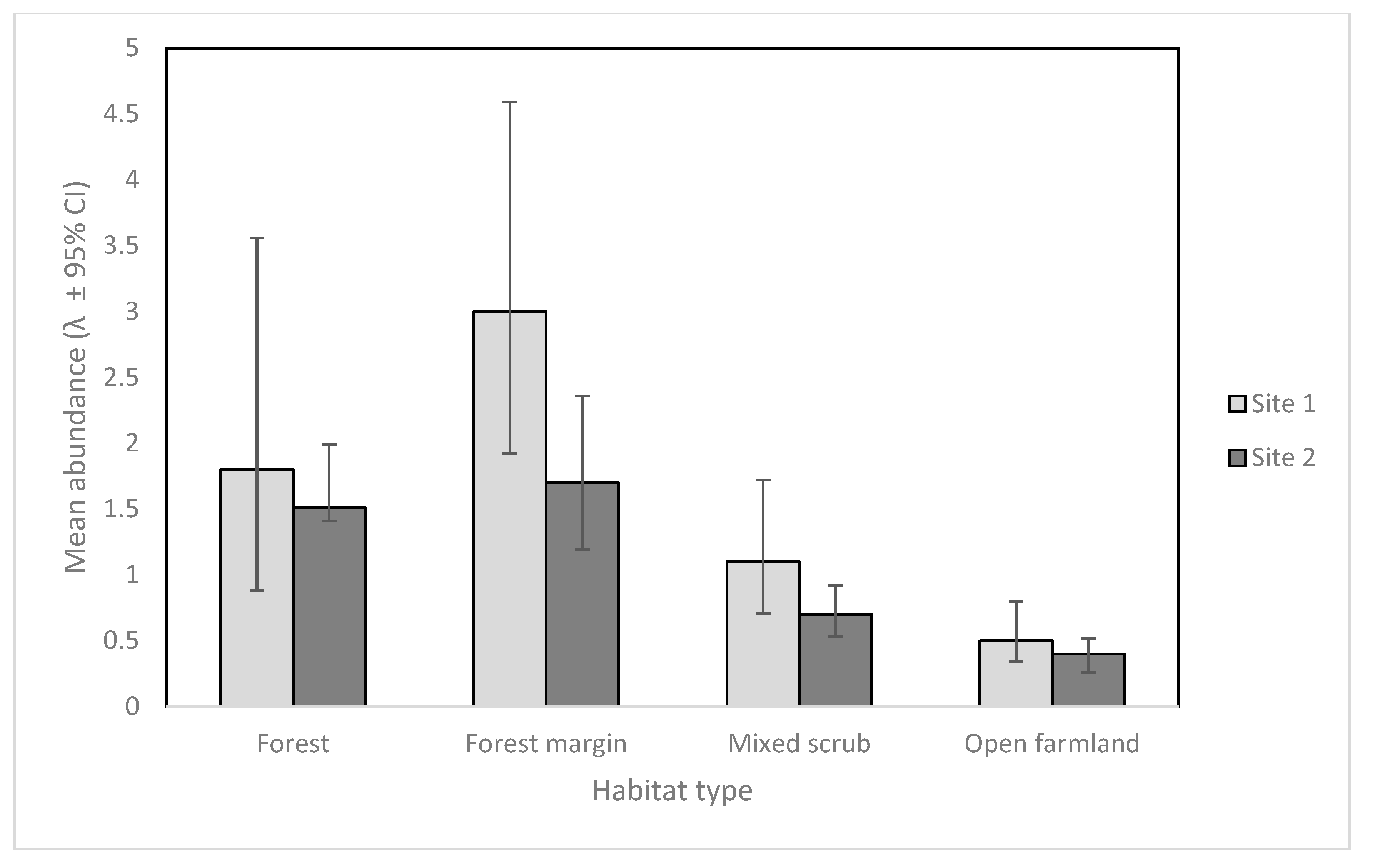
3. Results
4. Discussion
5. Conclusions
Future Research
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Royle, J.A.; Nichols, J.D. Estimating abundance from repeated presence–absence data or point counts. Ecology 2003, 84, 777–790. [Google Scholar] [CrossRef]
- Stanley, T.R.; Royle, J.A. Estimating site occupancy and abundance using indirect detection indices. J. Wildl. Manag. 2005, 69, 874–883. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Andrew Royle, J.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- MacKenzie, D.I. What are the issues with presence-absence data for wildlife managers? J. Wildl. Manag. 2005, 69, 849–860. [Google Scholar] [CrossRef]
- Dorazio, R.M.; Royle, J.A. Estimating size and composition of biological communities by modeling the occurrence of species. J. Am. Stat. Assoc. 2005, 100, 389–398. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Royle, J.A.; Brown, J.A.; Nichols, J.D. Occupancy estimation and modeling for rare and elusive populations. In Sampling Rare or Elusive Species; Thompson, W.L., Ed.; Island Press: Washington, DC, USA, 2004; Volume 8, pp. 149–171. [Google Scholar]
- Tobler, M.W.; Zúñiga Hartley, A.; Carrillo-Percastegui, S.E.; Powell, G.V. Spatiotemporal hierarchical modelling of species richness and occupancy using camera trap data. J. Appl. Ecol. 2015, 52, 413–421. [Google Scholar] [CrossRef]
- Fancourt, B.A. Avoiding the subject: The implications of avoidance behaviour for detecting predators. Behav. Ecol. Sociobiol. 2016, 70, 1535–1546. [Google Scholar] [CrossRef]
- Allen, M.L.; Evans, B.E.; Wheeler, M.E.; Mueller, M.A.; Pemble, K.; Olson, E.R.; Van Deelen, T.R. Survey techniques, detection probabilities, and the relative abundance of the carnivore guild on the Apostle Islands (2014–2016). arXiv 2017, arXiv:1703.10726v3. [Google Scholar]
- Royle, J.A. Site occupancy models with heterogeneous detection probabilities. Biometrics 2006, 62, 97–102. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.; Hines, J.E. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence, 2nd ed.; Academic Press: London, UK, 2017. [Google Scholar]
- Rich, L.N.; Davis, C.L.; Farris, Z.J.; Miller, D.A.; Tucker, J.M.; Hamel, S.; Kane, M.D. Assessing global patterns in mammalian carnivore occupancy and richness by integrating local camera trap surveys. Global Ecol. Biogeogr. 2017, 26, 918–929. [Google Scholar] [CrossRef]
- Linden, D.W.; Fuller, A.K.; Royle, J.A.; Hare, M.P. Examining the occupancy–density relationship for a low–density carnivore. J. Appl. Ecol. 2017, 54, 2043–2052. [Google Scholar] [CrossRef]
- Sarmento, P.; Cruz, J.; Eira, C.; Fonseca, C. Evaluation of camera trapping for estimating red fox abundance. J. Wildl. Manag. 2009, 73, 1207–1212. [Google Scholar] [CrossRef]
- Robley, A.; Gormley, A.; Woodford, L.; Lindeman, M.; Whitehead, B.; Albert, R.; Smith, A. Evaluation of Camera Trap Sampling Designs Used to Determine Change in Occupancy Rate and Abundance of Feral Cats; PRINTROOM 77 St Georges Rd, Preston 3072: Melbourne, Australia, April 2010. [Google Scholar]
- Bengsen, A.; Butler, J.; Masters, P. Estimating and indexing feral cat population abundances using camera traps. Wildl. Res. 2012, 38, 732–739. [Google Scholar] [CrossRef]
- Glen, A.S.; Anderson, D.; Veltman, C.J.; Garvey, P.M.; Nichols, M. Wildlife detector dogs and camera traps: A comparison of techniques for detecting feral cats. N. Z. J. Zool. 2016, 43, 127–137. [Google Scholar] [CrossRef]
- Pebsworth, P.A.; LaFleur, M. Advancing primate research and conservation through the use of camera traps: Introduction to the special issue. Int. J. Primatol. 2014, 35, 825–840. [Google Scholar] [CrossRef]
- Colyn, R.B.; Radloff, F.G.T.; O’Riain, M.J. Camera trapping mammals in the scrubland’s of the Cape Floristic Kingdom–the importance of effort, spacing and trap placement. Biodivers. Conserv. 2018, 27, 503–520. [Google Scholar] [CrossRef]
- Smith, J.K.; Coulson, G. A comparison of vertical and horizontal camera trap orientations for detection of potoroos and bandicoots. Aust. Mammal. 2012, 34, 196–201. [Google Scholar] [CrossRef]
- Nichols, M.; Glen, A.S.; Garvey, P.; Ross, J. A comparison of horizontal versus vertical camera placement to detect feral cats and mustelids. N. Z. J. Ecol. 2017, 41, 145–150. [Google Scholar] [CrossRef]
- Du Preez, B.D.; Loveridge, A.J.; Macdonald, D.W. To bait or not to bait: A comparison of camera-trapping methods for estimating leopard Panthera pardus density. Biol. Conserv. 2014, 176, 153–161. [Google Scholar] [CrossRef]
- Thorn, M.; Scott, D.M.; Green, M.; Bateman, P.W.; Cameron, E.Z. Estimating brown hyaena occupancy using baited camera traps. Afr. J. Wildl. Res. 2009, 39, 1–11. [Google Scholar] [CrossRef]
- Meek, P.D.; Ballard, G.; Claridge, A.; Kays, R.; Moseby, K.; O’brien, T.; Townsend, S. Recommended guiding principles for reporting on camera trapping research. Biodivers. Conserv. 2014, 23, 2321–2343. [Google Scholar] [CrossRef]
- Gerber, B.D.; Karpanty, S.M.; Kelly, M.J. Evaluating the potential biases in carnivore capture-recapture studies associated with the use of lure and varying density estimation techniques using photographic-sampling data of the Malagasy civet. Popul. Ecol. 2012, 54, 43–54. [Google Scholar] [CrossRef]
- Nichols, M.; Glen, A. Camera trapping to monitor the results of predator removal on Waitere Station. 2015. Available online: https://www.capetocity.co.nz/assets/Reports/Nichols-and-Glen-2015-Camera-trapping-to-monitor-the-results-of-predator-removal-on-Waitere-Station.pdf (accessed on 17 July 2018).
- Mann, G.K.; O’Riain, M.J.; Parker, D.M. The road less travelled: Assessing variation in mammal detection probabilities with camera traps in a semi-arid biodiversity hotspot. Biodivers. Conserv. 2015, 24, 531–545. [Google Scholar] [CrossRef]
- Medina, F.M.; Bonnaud, E.; Vidal, E.; Tershy, B.R.; Zavaleta, E.S.; Josh Donlan, C.; Nogales, M. A global review of the impacts of invasive cats on island endangered vertebrates. Glob. Chang. Biol. 2011, 17, 3503–3510. [Google Scholar] [CrossRef]
- Doherty, T.S.; Dickman, C.R.; Johnson, C.N.; Legge, S.M.; Ritchie, E.G.; Woinarski, J.C. Impacts and management of feral cats Felis catus in Australia. Mamm. Rev. 2017, 47, 83–97. [Google Scholar] [CrossRef]
- Rouco, C.; de Torre-Ceijas, R.; Martín-Collado, D.; Byrom, A.E. New Zealand Shouldn’t Ignore Feral Cats. BioScience 2017, 67, 686. [Google Scholar] [CrossRef]
- Glen, A.S.; Norbury, D.; Garvey, P.; Dickson, R. Effectiveness of feral cat control using para-aminopropiophenon (PAPP) on Toronui Station. 2017. Available online: https://www.capetocity.co.nz/assets/Reports/Glen-et-al-2017-Toronui-PAPP-trial.pdf (accessed on 17 July 2018).
- Gillies, C.A.; Fitzgerald, B.M. Feral cat. In The Handbook of New Zealand Mammals; King, C.M., Ed.; Oxford University Press: Melbourne, Australia, 2005; pp. 308–326. [Google Scholar]
- Langham, N.P.E.; Porter, R.E.R. Feral cats (Felis catus L.) on New Zealand farmland. I. Home range. Wildl. Res. 1991, 18, 741–760. [Google Scholar] [CrossRef]
- Fitzgerald, B.M.; Karl, B.J. Home range of feral house cats (Felis catus L.) in forest of the Orongorongo Valley, Wellington, New Zealand. N. Z. J. Ecol. 1986, 9, 71–82. [Google Scholar]
- Glen, A.S.; Byrom, A.E. Implications of Landholder Buy-in for the Success of Regional-scale Predator Control: Part 1: Review of Predator Movements. 2014. Available online: https://www.capetocity.co.nz/assets/Reports/UR-Glen-and-Byrom-Implications-of-landholder-buy-in...-2015.pdf (accessed on 17 July 2018).
- Kelly, M.J.; Holub, E.L. Camera trapping of carnivores: Trap success among camera types and across species, and habitat selection by species, on Salt Pond Mountain, Giles County, Virginia. North East. Nat. 2008, 15, 249–263. [Google Scholar] [CrossRef]
- Garvey, P.M.; Glen, A.S.; Clout, M.N.; Wyse, S.V.; Nichols, M.; Pech, R.P. Exploiting interspecific olfactory communication to monitor predators. Ecol. Appl. 2017, 27, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Fiske, I.; Chandler, R. Unmarked: An R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef]
- The R Development Core Team. R: A language and environment for statistical computing. 2016. Available online: https://www.r-project.org/ (accessed on 3 March 2019).
- Ramsey, D.S.; Caley, P.A.; Robley, A. Estimating population density from presence–absence data using a spatially explicit model. J. Wildl. Manag. 2015, 79, 491–499. [Google Scholar] [CrossRef]
- Russell, J.C.; Innes, J.G.; Brown, P.H.; Byrom, A.E. Predator-free New Zealand: Conservation country. BioScience 2015, 65, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Comer, S.; Speldewinde, P.; Tiller, C.; Clausen, L.; Pinder, J.; Cowen, S.; Algar, D. Evaluating the efficacy of a landscape scale feral cat control program using camera traps and occupancy models. Sci. Rep. 2018, 8, 5335. [Google Scholar] [CrossRef] [PubMed]
- Bengsen, A.J.; Algar, D.; Ballard, G.; Buckmaster, T.; Comer, S.; Fleming, P.J.; Zewe, F. Feral cat home-range size varies predictably with landscape productivity and population density. J. Zool. 2016, 298, 112–120. [Google Scholar] [CrossRef]
- Doherty, T.S.; Bengsen, A.J.; Davis, R.A. A critical review of habitat use by feral cats and key directions for future research and management. Wildl. Res. 2015, 41, 435–446. [Google Scholar] [CrossRef]
- Molsher, R.; Newsome, A.E.; Newsome, T.M.; Dickman, C.R. Mesopredator management: Effects of red fox control on the abundance, diet and use of space by feral cats. PLoS ONE 2017, 12, e0168460. [Google Scholar] [CrossRef] [PubMed]
- Fisher, P.; Algar, D.; Murphy, E.; Johnston, M.; Eason, C. How does cat behaviour influence the development and implementation of monitoring techniques and lethal control methods for feral cats? Appl. Anim. Behav. Sci. 2015, 173, 88–96. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Habitat Type | Number of Cameras (Site 1) | Number of Cameras (Site 2) | Mean Detections by Night (Site 1) | Mean Detections by Night (Site 2) |
|---|---|---|---|---|
| Forest | 2 | 23 | 0.2 | 2.5 |
| Forest Margin | 5 | 12 | 1 | 1.5 |
| Mixed scrub | 9 | 15 | 0.6 | 0.6 |
| Open farmland | 22 | 7 | 0.9 | 0.14 |
| Site 1 | |||||
|---|---|---|---|---|---|
| Model selection based on AICc: | AICc | Delta AICc | AICc Wt | Cum.Wt | LL |
| Abundance varies | 361.79 | 0.0 | 0.79 | 0.79 | −174.96 |
| Null model | 365.19 | 3.4 | 0.14 | 0.93 | −180.42 |
| Abundance and detection probability vary | 366.69 | 4.9 | 0.07 | 1.00 | −172.86 |
| Site 2 | |||||
| Model selection based on AICc: | AICc | Delta AICc | AICc Wt | Cum.Wt | LL |
| Abundance varies | 622.73 | 0.00 | 0.64 | 0.64 | −305.78 |
| Null model | 624.35 | 1.62 | 0.28 | 0.92 | −310.06 |
| Abundance and detection probability vary | 626.84 | 4.11 | 0.08 | 1.00 | −303.92 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nichols, M.; Ross, J.; Glen, A.S.; Paterson, A.M. An Evaluation of Systematic Versus Strategically-Placed Camera Traps for Monitoring Feral Cats in New Zealand. Animals 2019, 9, 687. https://doi.org/10.3390/ani9090687
Nichols M, Ross J, Glen AS, Paterson AM. An Evaluation of Systematic Versus Strategically-Placed Camera Traps for Monitoring Feral Cats in New Zealand. Animals. 2019; 9(9):687. https://doi.org/10.3390/ani9090687
Chicago/Turabian StyleNichols, Margaret, James Ross, Alistair S. Glen, and Adrian M. Paterson. 2019. "An Evaluation of Systematic Versus Strategically-Placed Camera Traps for Monitoring Feral Cats in New Zealand" Animals 9, no. 9: 687. https://doi.org/10.3390/ani9090687
APA StyleNichols, M., Ross, J., Glen, A. S., & Paterson, A. M. (2019). An Evaluation of Systematic Versus Strategically-Placed Camera Traps for Monitoring Feral Cats in New Zealand. Animals, 9(9), 687. https://doi.org/10.3390/ani9090687