Investigating the Role of Prolactin as a Potential Biomarker of Stress in Castrated Male Domestic Dogs


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Place
2.2. Video Analysis
2.3. Presence of Fear in the Collection Room
2.4. Serum Samples Collection and Storing
2.5. ELISA Kit
2.6. Statistical Analysis
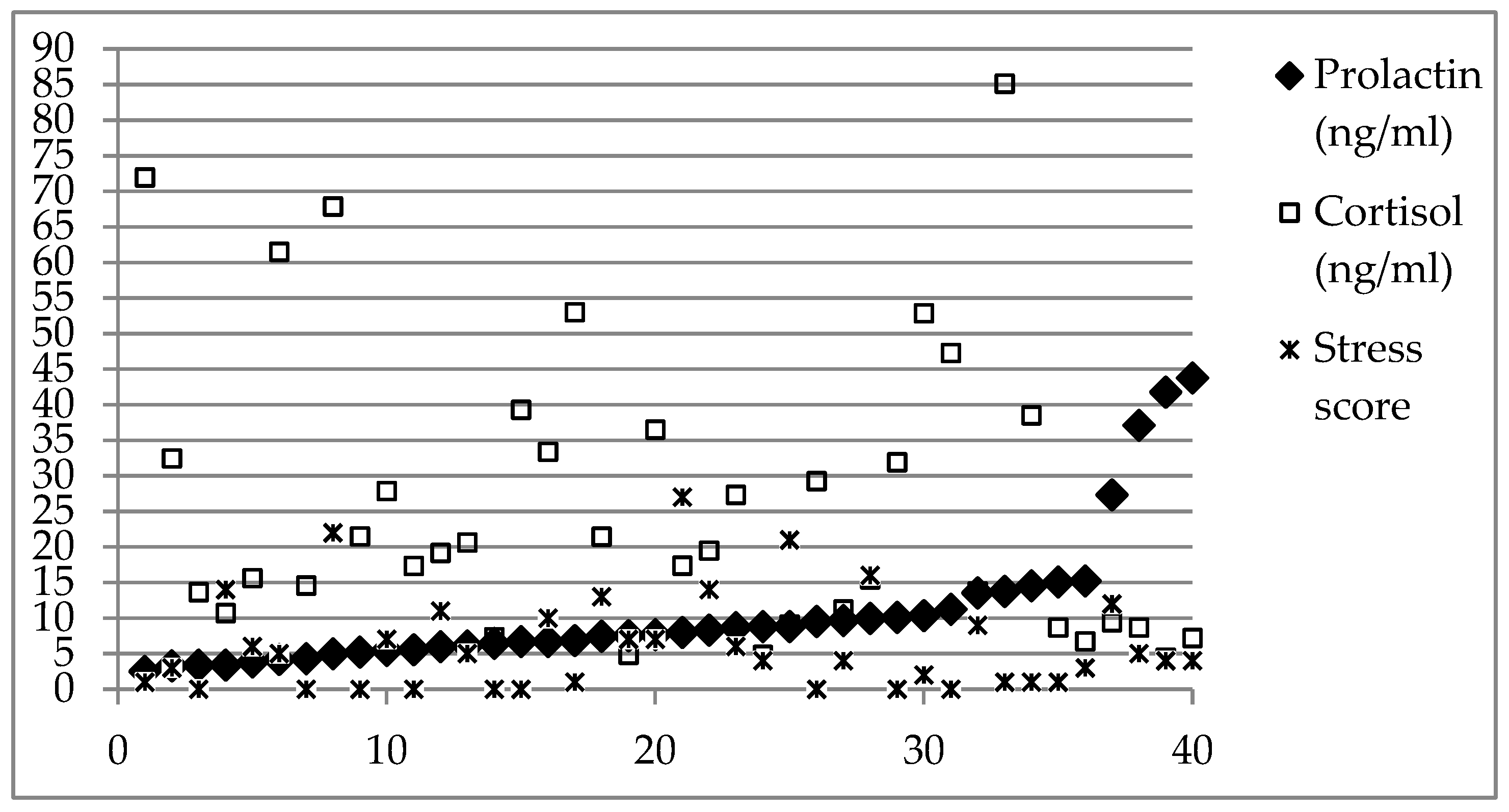
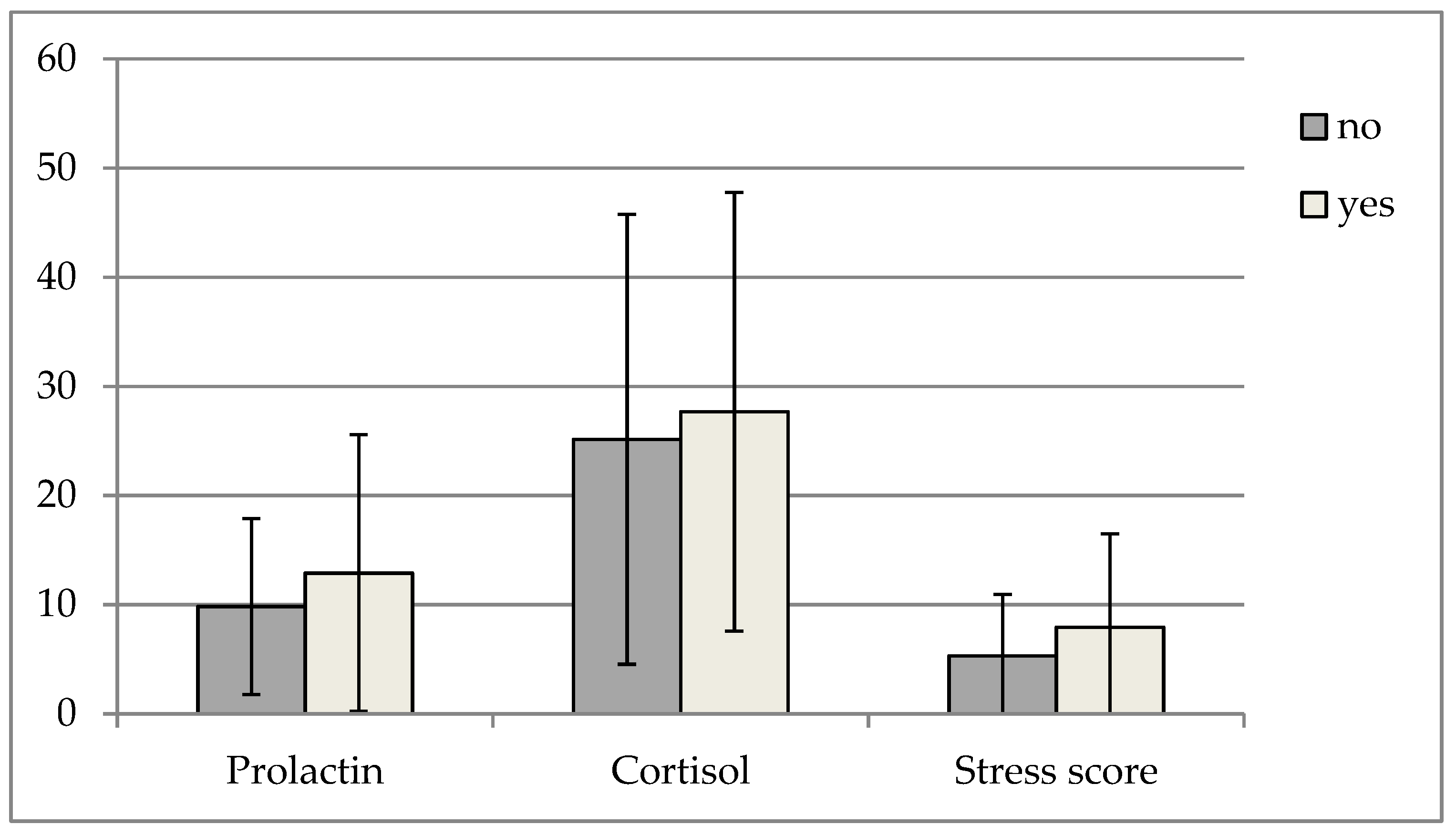
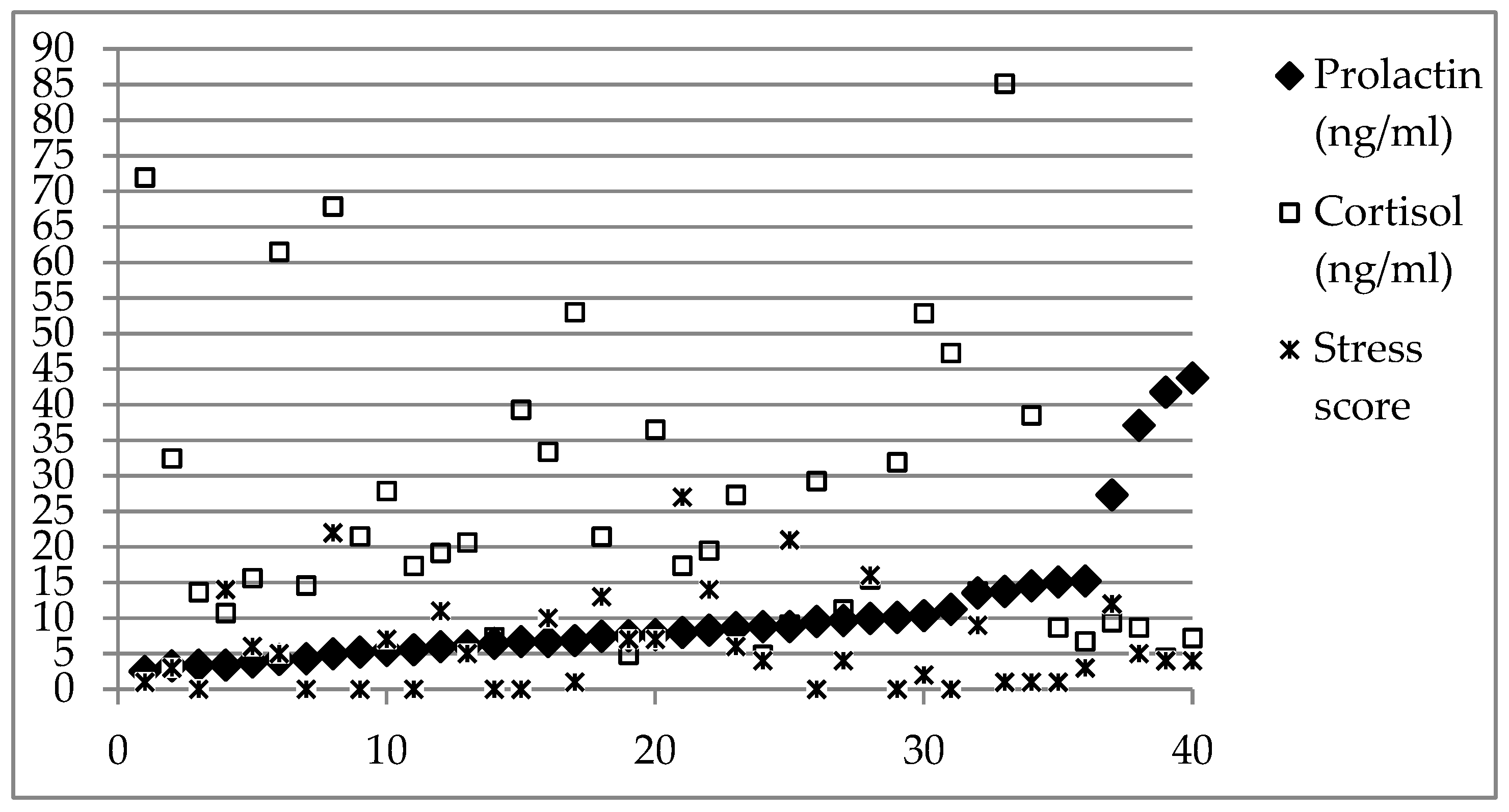
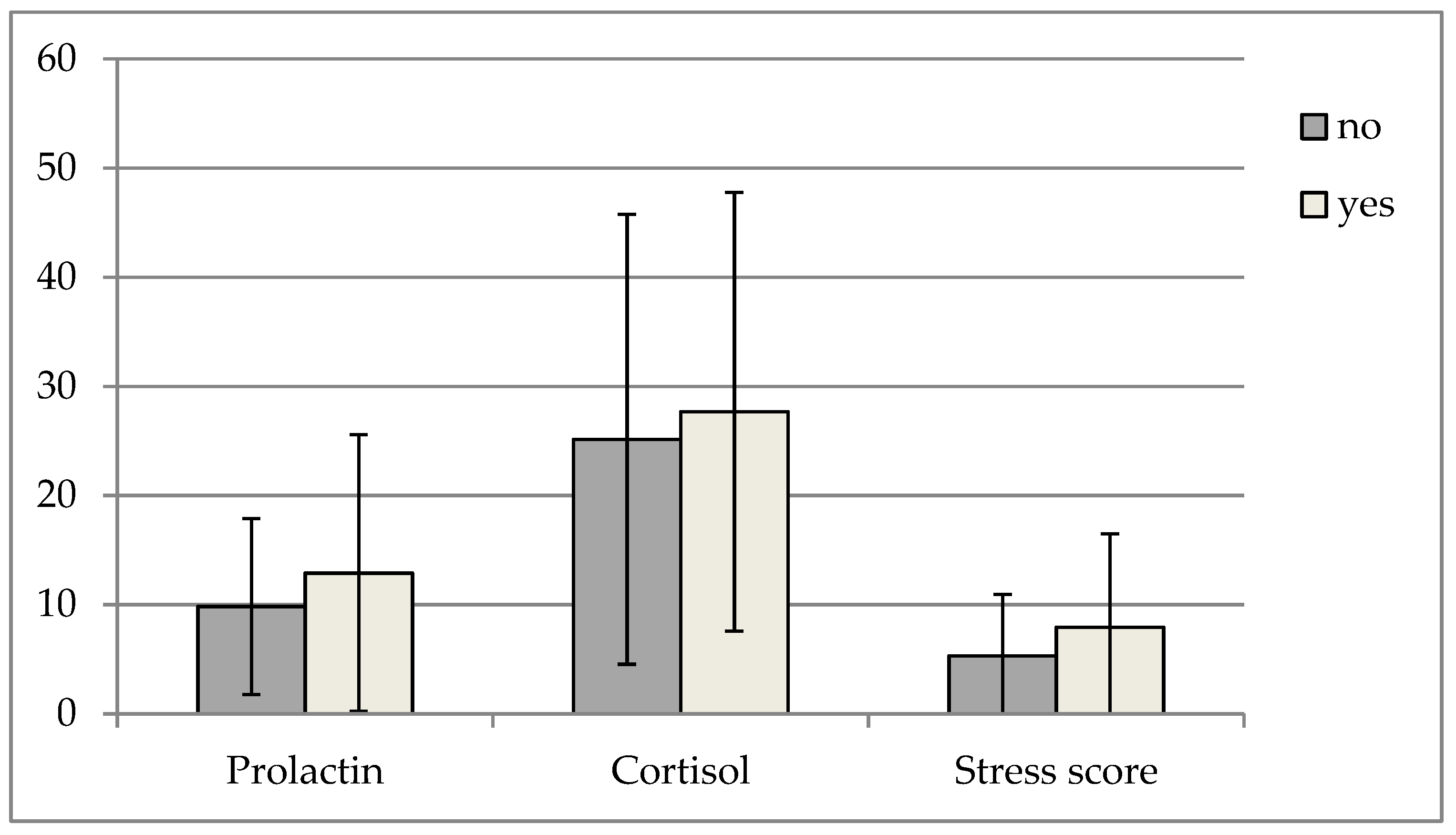
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bole-Feysot, C.; Goffin, V.; Edery, M.; Binart, N.; Kelly, P.A. Prolactin (PRL) and its receptor: Actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocr. Rev. 1998, 19, 225–268. [Google Scholar] [CrossRef] [PubMed]
- Bernard, V.; Young, J.; Binart, N. Prolactin—A pleiotropic factor in health and disease. Nat. Rev. Endocrinol. 2019, 15, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Euker, J.S.; Meites, J.; Riegle, G.D. Effects of acute stress on serum LH and prolactin in intact, castrate and dexamethasone-treated male rats. Endocrinology 1975, 96, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Döhler, K.-D.; Gärtner, K.; von zur Mühlen, A.; Döhler, U. Activation of anterior pituitary. thyroid and adrenal gland in rats after disturbance stress. Eur. J. Endocrinol. 1977, 86, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Van Vugt, D.A.; Bruni, J.F.; Meites, J. Naloxone inhibition of stress-induced increase in prolactin secretion. Life Sci. 1978, 22, 85–89. [Google Scholar] [CrossRef]
- Alamer, M. The role of prolactin in thermoregulation and water balance during heat stress in domestic ruminants. Asian J. Anim. Vet. Adv. 2011, 6, 1153–1169. [Google Scholar] [CrossRef]
- Kataria, N.; Kataria, A. Ambience-associated variation in serum biomarkers of oxidative stress in donkeys of arid tracts in India. Egypt. J. Biol. 2013, 15, 44–47. [Google Scholar] [CrossRef]
- Yayou, K.I.; Ito, S.; Yamamoto, N.; Kitagawa, S.; Okamura, H. Relationships of stress responses with plasma oxytocin and prolactin in heifer calves. Physiol. Behav. 2010, 99, 362–369. [Google Scholar] [CrossRef]
- Kataria, N.; Kataria, A.K. Use of prolactin as an indicator of stress in Marwari sheep from arid tracts in India. Comp. Clin. Path. 2011, 20, 333–336. [Google Scholar] [CrossRef]
- Manzon, L.A. The role of prolactin in fish osmoregulation: A review. Gen. Comp. Endocrinol. 2002, 125, 291–310. [Google Scholar] [CrossRef]
- Kirk, S.E.; Xie, T.Y.; Steyn, F.J.; Grattan, D.R.; Bunn, S.J. Restraint stress increases prolactin-mediated phosphorylation of signal transducer and activator of transcription 5 in the hypothalamus and adrenal cortex in the male mouse. J. Neuroendocrinol. 2017, 29, 1–9. [Google Scholar] [CrossRef]
- Rojas Vega, S.; Hollmann, W.; Strüder, H.K. Influences of Exercise and Training on the Circulating Concentration of Prolactin in Humans. J. Neuroendocrinol. 2012, 24, 395–402. [Google Scholar] [CrossRef]
- Lennartsson, A.-K.; Jonsdottir, I.H. Prolactin in response to acute psychosocial stress in healthy men and women. Psychoneuroendocrinology 2011, 36, 1530–1539. [Google Scholar] [CrossRef] [PubMed]
- Noel, G.L.; Suh, H.K.; Stone, J.G.; Frantz, A.G. Human prolactin and growth hormone release during surgery and other conditions of stress. J. Clin. Endocrinol. Metab. 1972, 35, 840–851. [Google Scholar] [CrossRef] [PubMed]
- Noel, G.L.; Dimond, R.C.; Earll, J.M.; Frantz, A.G. Prolactin, thyrotropin, and growth hormone release during stress associated with parachute jumping. Aviat. Sp. Environ. Med. 1976, 47, 543–547. [Google Scholar]
- Vaernes, R.; Ursin, H.; Darragh, A.; Lambe, R. Endocrine response patterns and psychological correlates. J. Psychosom. Res. 1982, 26, 123–131. [Google Scholar] [CrossRef]
- Jeffcoate, W.J.; Lincoln, N.B.; Selby, C.; Herbert, M. Correlation between anxiety and serum prolactin in humans. J. Psychosom. Res. 1986, 30, 217–222. [Google Scholar] [CrossRef]
- Baumgartner, A.; Gräf, K.J.; Kürten, I. Prolactin in patients with major depressive disorder and in healthy subjects. III. Investigation of basal and Post-TRH prolactin during different forms of acute and chronic psychological stress. Biol. Psychiatry 1988, 24, 286–298. [Google Scholar] [CrossRef]
- Modell, E.; Goldstein, D.; Reyes, F. Endocrine and behavioral responses to psychological stress in hyderandrogenic women. Int. J. Gynecol. Obstet. 1991, 34, 90. [Google Scholar] [CrossRef]
- Arnetz, B.B.; Fjellner, B.; Eneroth, P.; Kallner, A. Stress and psoriasis: Psychoendocrine and metabolic reactions in psoriatic patients during standardized stressor exposure. Psychosom. Med. 1985, 47, 528–541. [Google Scholar] [CrossRef]
- Gerra, G.; Zaimovic, A.; Mascetti, G.G.; Gardini, S.; Zambelli, U.; Timpano, M.; Raggi, M.A.; Brambilla, F. Neuroendocrine responses to experimentally-induced psychological stress in healthy humans. Psychoneuroendocrinology 2001, 26, 91–107. [Google Scholar] [CrossRef]
- Sobrinho, L.G.; Simões, M.; Barbosa, L.; Raposo, J.F.; Pratas, S.; Fernandes, P.L.; Santos, M.A. Cortisol, prolactin, growth hormone and neurovegetative responses to emotions elicited during an hypnoidal state. Psychoneuroendocrinology 2003, 28, 1–17. [Google Scholar] [CrossRef]
- Biondi, M.; Picardi, A. Psychological stress and neuroendocrine function in humans: The last two decades of research. Psychother. Psychosom. 1999, 68, 114–150. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.E.; Herbert, M.; Walder, C.P.; Selby, C.; Jeffcoate, W.J. Prolactin and stress: Some endocrine correlates of pre-operative anxiety. Clin. Endocrinol. (Oxf.) 1986, 24, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, C.; Manteca, X.; Cerón, J.; Martínez-Subiela, S.; Cuenca, R.; Lavin, S.; Garcia, F.; Pastor, J. Perioperative stress response in dogs undergoing elective surgery: Variations in behavioural, neuroendocrine, immune and acute phase responses. Anim. Welf. 2008, 17, 259–273. [Google Scholar]
- Marrocco-Trischitta, M.M.; Tiezzi, A.; Svampa, M.G.; Bandiera, G.; Camilli, S.; Stillo, F.; Petasecca, P.; Sampogna, F.; Abeni, D.; Guerrini, P. Perioperative stress response to carotid endarterectomy: The impact of anesthetic modality. J. Vasc. Surg. 2004, 39, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- Pageat, P.; Gaultier, E. Current research in canine and feline pheromones. Vet. Clin. North Am.-Small Anim. Pract. 2003, 33, 187–211. [Google Scholar] [CrossRef]
- Pageat, P.; Lafont, C.; Falewée, C.; Bonnafous, L.; Gaultier, E.; Silliart, B. An evaluation of serum prolactin in anxious dogs and response to treatment with selegiline or fluoxetine. Appl. Anim. Behav. Sci. 2007, 105, 342–350. [Google Scholar] [CrossRef]
- Berteselli, G.V.; Servida, F.; Dall’Ara, P.; Verga, M.; Piola, E.; Puricelli, M.; Palestrini, C. Evaluation of immunological, stress and behavioural parameters in dogs (Canis familiaris) with anxiety related disorders. In Current Issues and Research in Veterinary Behavioral Medicine; Mills, D., Levine, E., Landsberg, G., Horwitz, D., Duxbury, M., Meyer, K., Huntley, L.R., Reich, M., Willard, J., Eds.; Purdue University Press: West Lafayette, IN, USA, 2005; pp. 18–22. ISBN 1557534098. [Google Scholar]
- Odendaal, J.S.J.; Meintjes, R.A. Neurophysiological correlates of affiliative behaviour between humans and dogs. Vet. J. 2003, 165, 296–301. [Google Scholar] [CrossRef]
- Cook, N.J. Review: Minimally invasive sampling media and the measurement of corticosteroids as biomarkers of stress in animals. Can. J. Anim. Sci. 2012, 92, 227–259. [Google Scholar] [CrossRef]
- Van Uum, S.H.M.; Sauvé, B.; Fraser, L.A.; Morley-Forster, P.; Paul, T.L.; Koren, G. Elevated content of cortisol in hair of patients with severe chronic pain: A novel biomarker for stress. Stress 2008, 11, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, C.; Tietze, A.; Skoluda, N.; Dettenborn, L. Hair as a retrospective calendar of cortisol production-Increased cortisol incorporation into hair in the third trimester of pregnancy. Psychoneuroendocrinology 2009, 34, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Heimbürge, S.; Kanitz, E.; Otten, W. The use of hair cortisol for the assessment of stress in animals. Gen. Comp. Endocrinol. 2019, 270, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Miranda-de la Lama, G.C.; Rivero, L.; Chacón, G.; Garcia-Belenguer, S.; Villarroel, M.; Maria, G.A. Effect of the pre-slaughter logistic chain on some indicators of welfare in lambs. Livest. Sci. 2010, 128, 52–59. [Google Scholar] [CrossRef]
- Part, C.E.; Kiddie, J.L.; Hayes, W.A.; Mills, D.S.; Neville, R.F.; Morton, D.B.; Collins, L.M. Physiological, physical and behavioural changes in dogs (Canis familiaris) when kennelled: Testing the validity of stress parameters. Physiol. Behav. 2014, 133, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Mason, G.; Mendl, M. Why is there no simple way of measuring animal welfare? Anim. Welf. 1993, 2, 301–319. [Google Scholar]
- Mormède, P.; Andanson, S.; Aupérin, B.; Beerda, B.; Guémené, D.; Malmkvist, J.; Manteca, X.; Manteuffel, G.; Prunet, P.; van Reenen, C.G.; et al. Exploration of the hypothalamic-pituitary-adrenal function as a tool to evaluate animal welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef] [PubMed]
- Angle, C.T.; Wakshlag, J.J.; Gillette, R.L.; Stokol, T.; Geske, S.; Adkins, T.O.; Gregor, C. Hematologic, serum biochemical, and cortisol changes associated with anticipation of exercise and short duration high-intensity exercise in sled dogs. Vet. Clin. Pathol. 2009, 38, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Ralph, C.R.; Tilbrook, A.J. Invited Review: The usefulness of measuring glucocorticoids for assessing animal welfare. J. Anim. Sci. 2016, 94, 457–470. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar]
- Trevisi, E.; Bertoni, G. Some physiological and biochemical methods for acute and chronic stress evaluationin dairy cows. Ital. J. Anim. Sci. 2009, 8, 265–286. [Google Scholar] [CrossRef]
- Bennett, A.; Hayssen, V. Measuring cortisol in hair and saliva from dogs: Coat color and pigment differences. Domest. Anim. Endocrinol. 2010, 39, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Kim, E.; Choi, M.H. Technical and clinical aspects of cortisol as a biochemical marker of chronic stress. BMB Rep. 2015, 48, 209. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Ebihara, S.; Nakamura, Y.; Takahashi, K. A model of human sleep-related growth hormone secretion in dogs: Effects of 3, 6, and 12 hours of forced wakefulness on plasma growth hormone, cortisol, and sleep stages. Endocrinology 1981, 109, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Kemppainen, R.J.; Sartin, J.L. Evidence for episodic but not circadian activity in plasma concentrations of adrenocorticotrophin, cortisol and thyroxine in dogs. J. Endocrinol. 1984, 103, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolezvská, J.; Brunclík, V.; Svoboda, M. Circadian Rhythm of Cortisol Secretion in Dogs of Different Daily Activities. Acta Vet. Brno. 2003, 72, 599–605. [Google Scholar] [CrossRef]
- Protopopova, A. Effects of sheltering on physiology, immune function, behavior, and the welfare of dogs. Physiol. Behav. 2016, 159, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Beerda, B.; Schilder, M.B.H.; Van Hooff, J.A.R.A.M.; De Vries, H.W.; Mol, J.A. Behavioural and hormonal indicators of enduring environmental stress in dogs. Anim. Welf. 2000, 9, 49–62. [Google Scholar]
- Stalder, T.; Kirschbaum, C.; Alexander, N.; Bornstein, S.R.; Gao, W.; Miller, R.; Stark, S.; Bosch, J.A.; Fischer, J.E. Cortisol in hair and the metabolic syndrome. J. Clin. Endocrinol. Metab. 2013, 98, 2573–2580. [Google Scholar] [CrossRef]
- Russell, E.; Koren, G.; Rieder, M.; Van Uum, S. Hair cortisol as a biological marker of chronic stress: Current status, future directions and unanswered questions. Psychoneuroendocrinology 2012, 37, 589–601. [Google Scholar] [CrossRef]
- Jaroenporn, S.; Nagaoka, K.; Kasahara, C.; Ohta, R.; Watanabe, G.; Taya, K. Physiological roles of prolactin in the adrenocortical response to acute restraint stress. Endocr. J. 2007, 54, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Beerda, B.; Schilder, M.B.H.; Van Hooff, J.A.R.A.M.; De Vries, H.W.; Mol, J.A. Behavioural, saliva cortisol and heart rate responses to different types of stimuli in dogs. Appl. Anim. Behav. Sci. 1998, 58, 365–381. [Google Scholar] [CrossRef]
- Mariti, C.; Gazzano, A.; Moore, J.L.; Baragli, P.; Chelli, L.; Sighieri, C. Perception of dogs’ stress by their owners. J. Vet. Behav. Clin. Appl. Res. 2012, 7, 213–219. [Google Scholar] [CrossRef]
- Beerda, B.; Schilder, M.B.H.; Van Hooff, J.A.R.A.M.; De Vries, H.W.; Mol, J.A. Chronic stress in dogs subjected to social and spatial restriction. I. Behavioral responses. Physiol. Behav. 1999, 66, 233–242. [Google Scholar] [CrossRef]
- Beerda, B.; Schilder, M.B.H.; Bernadina, W.; Van Hooff, J.A.R.A.M.; De Vries, H.W.; Mol, J.A. Chronic stress in dogs subjected to social and spatial restriction. II. Hormonal and immunological responses. Physiol. Behav. 1999, 66, 243–254. [Google Scholar] [CrossRef]
- Cozzi, A.; Mariti, C.; Ogi, A.; Sighieri, C.; Gazzano, A. Behavioral modification in sheltered dogs. Dog Behav. 2016, 2, 1–12. [Google Scholar]
- Siracusa, C.; Manteca, X.; Cuenca, R.; Del Mar Alcalá, M.; Alba, A.; Lavín, S.; Pastor, J. Effect of a synthetic appeasing pheromone on behavioral, neuroendocrine, immune, and acute-phase perioperative stress responses in dogs. J. Am. Vet. Med. Assoc. 2010, 237, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Friard, O.; Gamba, M. BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Gazzano, A.; Migoni, S.; Guardini, G.; Bowen, J.; Fatjò, J.; Mariti, C. Stress in aggressive dogs towards people: Behavioral analysis during consultation. Dog Behav. 2015, 1, 6–13. [Google Scholar]
- Stephen, J.M.; Ledger, R.A. An audit of behavioral indicators of poor welfare in kenneled dogs in the United Kingdom. J. Appl. Anim. Welf. Sci. 2005, 8, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Dreschel, N.A.; Granger, D.A. Physiological and behavioral reactivity to stress in thunderstorm-phobic dogs and their caregivers. Appl. Anim. Behav. Sci. 2005, 95, 153–168. [Google Scholar] [CrossRef]
- Rooney, N.; Gaines, S.; Hiby, E. A practitioner’s guide to working dog welfare. J. Vet. Behav. Clin. Appl. Res. 2009, 4, 127–134. [Google Scholar] [CrossRef]
- Polgár, Z.; Blackwell, E.J.; Rooney, N.J. Assessing the welfare of kennelled dogs—A review of animal-based measures. Appl. Anim. Behav. Sci. 2019, 213, 1–13. [Google Scholar] [CrossRef]
- Meyerhoff, J.L.; Oleshansky, M.A.; Mougey, E.H. Psychologic stress increases plasma levels of prolactin, cortisol, and POMC-derived peptides in man. Psychosom. Med. 1988, 50, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Richter, S.D.; Schürmeyer, T.H.; Schedlowski, M.; Hädicke, A.; Tewes, U.; Schmidt, R.E.; Wagner, T.O.F. Time kinetics of the endocrine response to acute psychological stress. J. Clin. Endocrinol. Metab. 1996, 81, 1956–1960. [Google Scholar] [PubMed]
- Jöchle, W. Prolactin in canine and feline reproduction. Reprod. Domest. Anim. 1997, 32, 183–193. [Google Scholar] [CrossRef]
- Gobello, C.; Colombani, M.; Scaglia, H.; De La Sota, R.L.; Goya, R.G. Heterogeneity of circulating prolactin in the bitch. Reprod. Nutr. Dev. 2001, 41, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Graf, K.J.; El Etreby, M.F. Endocrinology of reproduction in the female beagle dog and its significance in mammary gland tumorigenesis. Acta Endocrinol. Suppl. 1979, 222, 1–34. [Google Scholar]
- Graef, K.J.; Friedreich, E.; Matthes, S.; Hasan, S.H. Homologous radioimmunoassay for canine prolactin and its application in various physiological states. J. Endocrinol. 1977, 75, 93–103. [Google Scholar] [CrossRef]
- Corrada, Y.; Rimoldi, I.; Arreseigor, S.; Marecco, G.; Gobello, C. Prolactin reference range and pulsatility in male dogs. Theriogenology 2006, 66, 1599–1602. [Google Scholar] [CrossRef]
- Urhausen, C.; Seefeldt, A.; Eschricht, F.; Koch, A.; Hoppen, H.; Beyerbach, M.; Möhrke, C.; Dieleman, S.; Günzel-Apel, A. Concentrations of Prolactin, LH, Testosterone, TSH and Thyroxine in Normospermic Dogs of Different Breeds. Reprod. Domest. Anim. 2009, 44, 279–282. [Google Scholar] [CrossRef]
- Lane, M.B.; Flatland, B.; Olin, S.J.; Fecteau, K.A.; Rick, M.; Giori, L. Analytic performance evaluation of a veterinary-specific ELISA for measurement of serum cortisol concentrations of dogs. J. Am. Vet. Med. Assoc. 2018, 253, 1580–1588. [Google Scholar] [CrossRef] [PubMed]
- Cafazzo, S.; Maragliano, L.; Bonanni, R.; Scholl, F.; Guarducci, M.; Scarcella, R.; Di Paolo, M.; Pontier, D.; Lai, O.; Carlevaro, F.; et al. Behavioural and physiological indicators of shelter dogs’ welfare: Reflections on the no-kill policy on free-ranging dogs in Italy revisited on the basis of 15years of implementation. Physiol. Behav. 2014, 133, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Ishioka, K.; Hatai, H.; Komabayashi, K.; Soliman, M.M.; Shibata, H.; Honjoh, T.; Kimura, K.; Saito, M. Diurnal variations of serum leptin in dogs: Effects of fasting and re-feeding. Vet. J. 2005, 169, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.L.; Gelatt, K.N.; Gum, G.G. Serum hydrocortisone (cortisol) values in glaucomatous and normotensive Beagles. Am. J. Vet. Res. 1980, 41, 1516–1518. [Google Scholar]
- Garnier, F.; Benoit, E.; Virat, M.; Ochoa, R.; Delatour, P. Adrenal cortical response in clinically normal dogs before and after adaptation to a housing environment. Lab. Anim. 1990, 24, 40–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avella, M.; Schreck, C.B.; Prunet, P. Plasma prolactin and cortisol concentrations of stressed coho salmon, Oncorhynchus kisutch, in fresh water or salt water. Gen. Comp. Endocrinol. 1991, 81, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Kant, G.J.; Leu, J.R.; Anderson, S.M.; Mougey, E.H. Effects of chronic stress on plasma corticosterone, ACTH and prolactin. Physiol. Behav. 1987, 40, 775–779. [Google Scholar] [CrossRef]
- Cook, C.J. Oxytocin and prolactin suppress cortisol responses to acute stress in both lactating and non-lactating sheep. J. Dairy Res. 1997, 64, 327–339. [Google Scholar] [CrossRef]
- Bazan, J.F.; Bole-Feysot, C.; Goffin, V.; Edery, M.; Binart, N.; Kelly, P.A.; Carter, D.A.; Lightman, S.L.; Andrews, Z.B.; Kokay, I.C.; et al. Oxytocin responses to stress in lactating and hyperprolactinaemic rats. Endocrinology 2001, 46, 532–537. [Google Scholar]
- Van De Kar, L.D.; Blair, M.L. Forebrain pathways mediating stress-induced hormone secretion. Front. Neuroendocrinol. 1999, 20, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Ratner, A.; Yelvington, D.B.; Rosenthal, M. Prolactin and corticosterone response to repeated footshock stress in male rats. Psychoneuroendocrinology 1989, 14, 393–396. [Google Scholar] [CrossRef]
- Neumann, I.D.; Torner, L.; Wigger, A. Brain oxytocin: Differential inhibition of neuroendocrine stress responses and anxiety-related behaviour in virgin, pregnant and lactating rats. Neuroscience 1999, 95, 565–575. [Google Scholar] [CrossRef]
- Grattan, D.R. Behavioural significance of prolactin signalling in the central nervous system during pregnancy and lactation. Reproduction 2002, 123, 497–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuadra, G.; Zurita, A.; Lacerra, C.; Molina, V. Chronic stress sensitizes frontal cortex dopamine release in response to a subsequent novel stressor: Reversal by naloxone. Brain Res. Bull. 1999, 95, 567–575. [Google Scholar] [CrossRef]
- Coppola, C.L.; Grandin, T.; Enns, R.M. Human interaction and cortisol: Can human contact reduce stress for shelter dogs? Physiol. Behav. 2006, 87, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, M.B. Hypothalamic-pituitary-adrenal responses to brief social separation. Neurosci. Biobehav. Rev. 1997, 21, 11–29. [Google Scholar] [CrossRef]
- Hennessy, M.B.; Voith, V.L.; Mazzei, S.J.; Buttram, J.; Miller, D.D.; Linden, F. Behavior and cortisol levels of dogs in a public animal shelter, and an exploration of the ability of these measures to predict problem behavior after adoption. Appl. Anim. Behav. Sci. 2001, 73, 217–233. [Google Scholar] [CrossRef]
- Stephen, J.M.; Ledger, R.A. A longitudinal evaluation of urinary cortisol in kennelled dogs, Canis familiaris. Physiol. Behav. 2006, 87, 911–916. [Google Scholar] [CrossRef]
- Rooney, N.J.; Gaines, S.A.; Bradshaw, J.W.S. Behavioural and glucocorticoid responses of dogs (Canis familiaris) to kennelling: Investigating mitigation of stress by prior habituation. Physiol. Behav. 2007, 92, 847–854. [Google Scholar] [CrossRef]
- Righi, C.; Menchetti, L.; Orlandi, R.; Moscati, L.; Mancini, S.; Diverio, S. Welfare Assessment in Shelter Dogs by Using Physiological and Immunological Parameters. Animals 2019, 9, 340. [Google Scholar]
- Gutiérrez, J.; Gazzano, A.; Torracca, B.; Meucci, V.; Mariti, C. Determination of prolactin in canine saliva: Is it possible to use a commercial ELISA kit? Animals 2019, 9, 418. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Behaviour | Number and % of Dogs Displaying the Behaviour | Number of Occurrences | Behaviour | Number and % of Dogs Displaying the Behaviour | Number of Occurrences |
|---|---|---|---|---|---|
| Tongue out | 20; 50.0% | 101 | Circling | 3; 7.5% | 3 |
| Panting | 4; 10.0% | 38 | Eliminating | 3; 7.5% | 3 |
| Yawning | 15; 37.5% | 34 | Howling | 2; 5.0% | 3 |
| Paw lifting | 1; 2.5% | 21 | Turn head | 2; 5.0% | 2 |
| Behaviour against the fence | 3; 7.5% | 12 | Tuck tail | 1; 2.5% | 1 |
| Shaking | 9; 22.5% | 10 | Growling | 0; 0.0% | 0 |
| Pacing | 3; 7.5% | 7 | Hiding | 0; 0.0% | 0 |
| Cowering | 4; 10.0% | 6 | Salivation | 0; 0.0% | 0 |
| Whining | 4; 10.0% | 5 | Trembling | 0; 0.0% | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez, J.; Gazzano, A.; Pirrone, F.; Sighieri, C.; Mariti, C. Investigating the Role of Prolactin as a Potential Biomarker of Stress in Castrated Male Domestic Dogs. Animals 2019, 9, 676. https://doi.org/10.3390/ani9090676
Gutiérrez J, Gazzano A, Pirrone F, Sighieri C, Mariti C. Investigating the Role of Prolactin as a Potential Biomarker of Stress in Castrated Male Domestic Dogs. Animals. 2019; 9(9):676. https://doi.org/10.3390/ani9090676
Chicago/Turabian StyleGutiérrez, Jara, Angelo Gazzano, Federica Pirrone, Claudio Sighieri, and Chiara Mariti. 2019. "Investigating the Role of Prolactin as a Potential Biomarker of Stress in Castrated Male Domestic Dogs" Animals 9, no. 9: 676. https://doi.org/10.3390/ani9090676
APA StyleGutiérrez, J., Gazzano, A., Pirrone, F., Sighieri, C., & Mariti, C. (2019). Investigating the Role of Prolactin as a Potential Biomarker of Stress in Castrated Male Domestic Dogs. Animals, 9(9), 676. https://doi.org/10.3390/ani9090676