1. Introduction
Calcium (Ca) demands are understandably greater in modern sow lines with larger litter sizes and greater milk production capabilities. It has been reported that dietary Ca plays an important role in the skeletal development of piglets during late pregnancy [
1]. Notably, the Ca level in the diet also affects milk Ca and other mineral element concentrations [
2]. Thus, higher mineral requirements, particularly for Ca, during later pregnancy may be a limiting factor in colostrum synthesis in mammary glands [
3]. High prolificacy and prolonged farrowing in sows have been linked to hypocalcemic responses during farrowing; therefore, low levels or a low utilization rate of Ca in the diet fed to gestating sows can reduce litter size, prolong delivery time, increase the number of stillbirths, and result in a higher occurrence of skeletal problems in piglets [
4]. Consequently, improving the utilization of Ca and other mineral elements is important for the productive performance of sows.
The feeding of nutrients to animals must be coordinated with endogenous physiological rhythms to optimize mammalian production [
5], signifying the importance of feeding time. On the one hand, recent studies show that circadian clocks and energy metabolism interact, and nutrient utilization is affected by the time of feed intake even when the same type of feed and the same number of calories are consumed [
6,
7]. One study shows that feeding with a high energy diet in the evening increases the risk of obesity more than feeding with a high energy diet before 12:00, and that evening or night feeding increases milk fat synthesis and energy production [
8]. In addition, feeding pregnant sows at 15:30 will alter their energy and nutrient metabolism and improve their backfat (BF) thickness gain compared with conventional feeding regimes [
9], and crude protein feeding time modulates the lipid profiles in sow milk and plasma [
10]. On the other hand, the mRNA expression of circadian clocks in livestock is also affected by feeding time [
11], and one study also reported that Ca feeding time in laying hens affected the expression of genes related to the circadian clock and Ca transporters and led to circadian variations in serum Ca [
12]. Ca feeding time during late pregnancy affects fatty acid transport and circadian clock-related changes in the placenta of pigs [
13]. Overall, it was speculated that daily Ca feeding time may affect productive performance and mineral element metabolism in sows. However, there are few reports on such effects. Therefore, this study focuses on the productive performance of sows, the mineral element profiles of their milk and serum, and their metal transporters under conditions of daily dietary Ca feeding at 06:00 and 15:00.
4. Discussion
Metabolic chronophysiology is of high importance with regards to eating time effects on the health of livestock [
5], while the effect of Ca feeding time during late pregnancy and lactation on the performance of sows has never been studied. The study showed that under isocaloric conditions per kilogram of live metabolic weight, feeding time did not influence sow body weight (BW) and BW gain either during gestation or lactation [
9], because the nutritional needs of the sows for energy was not compromised at the different feeding times. Similar results also appeared in this experiment; the fact that BF gain and loss did not differ with reference to feeding time in this study may be due to the similar feed intake observed.
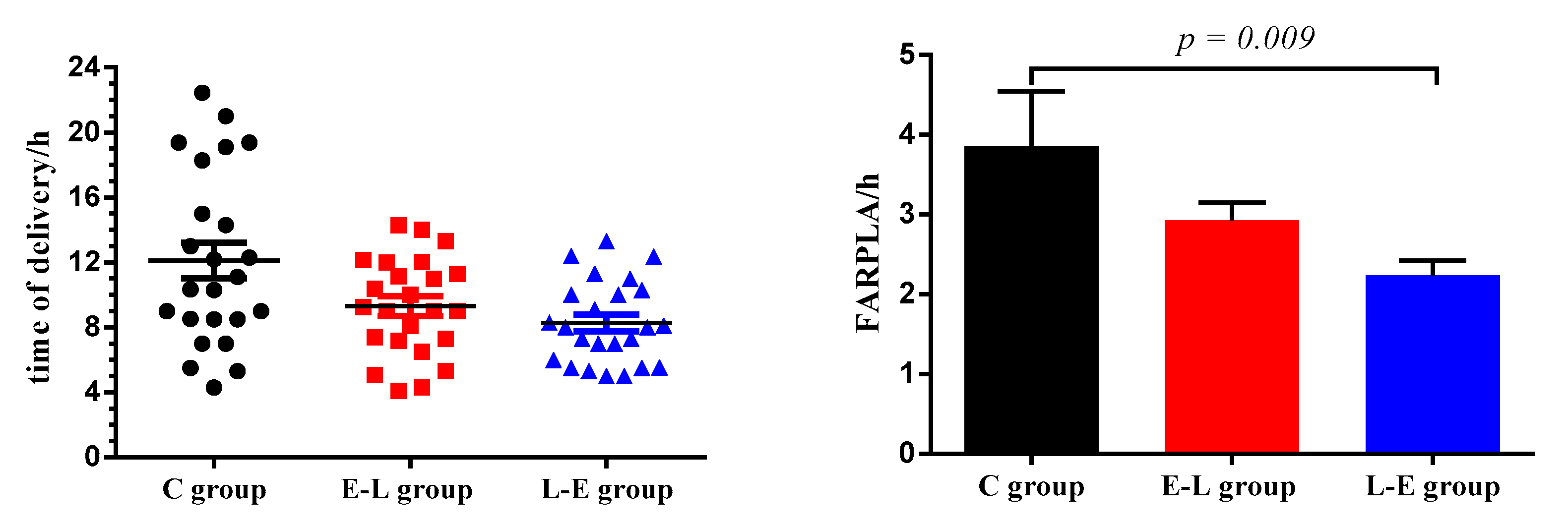
The Ca requirements of sows increase during late gestation and lactation [
4]. On the one hand, lower Ca utilization or insufficient Ca in the diet will lead to Ca deficiency, and inadequate mineral supplies may prolong delivery time, increase the number of stillbirths, and decrease the vigor of piglets [
4]. It has been demonstrated that feeding with different amounts of Ca during late pregnancy does not affect newborn birth weight or litter size [
21]. In the present study, feeding Ca to sows at 15:00 during late pregnancy significantly decreased the FARPLA of sows. Furthermore, the time of sow delivery in the L-E group was concentrated between 04:00−12:00 daily. Calcium is involved in muscle contraction during the delivery of sows. In addition, evening feeding improved nutrient digestibility [
5]. Therefore, we speculated that feeding Ca at 15:00 may improve the digestibility of Ca, which is beneficial for muscle contraction during sow delivery, but the specific mechanism needs further study. In addition, the study showed that there was a significant increase in the rate of stillbirths as the delivery time increased [
22]. This result is consistent with our experimental results, thus, it was believed that the shorter FARPLA of sows in the L-E group was the main reason for the lower rate of stillbirths. On the other hand, the average daily weight gain (ADG) of suckling piglets from sows fed low Ca diets (0.38% of Ca) or fed high Ca diets (1.12% of Ca) was greater than that of those from sows fed standard concentrations of Ca [
2]. Additionally, the study showed that the time of feeding was a determining factor in the weight gain of the mammal [
7], while feeding sows a daily meal at 15:30 had no difference in the ADG of suckling piglets compared with the control [
9]. Therefore, it was speculated that Ca feeding time may affect the Ca content in the blood and milk of sows, and that the utilization of Ca in suckling piglets was improved, which may be the main reason for increasing the ADG of piglets.
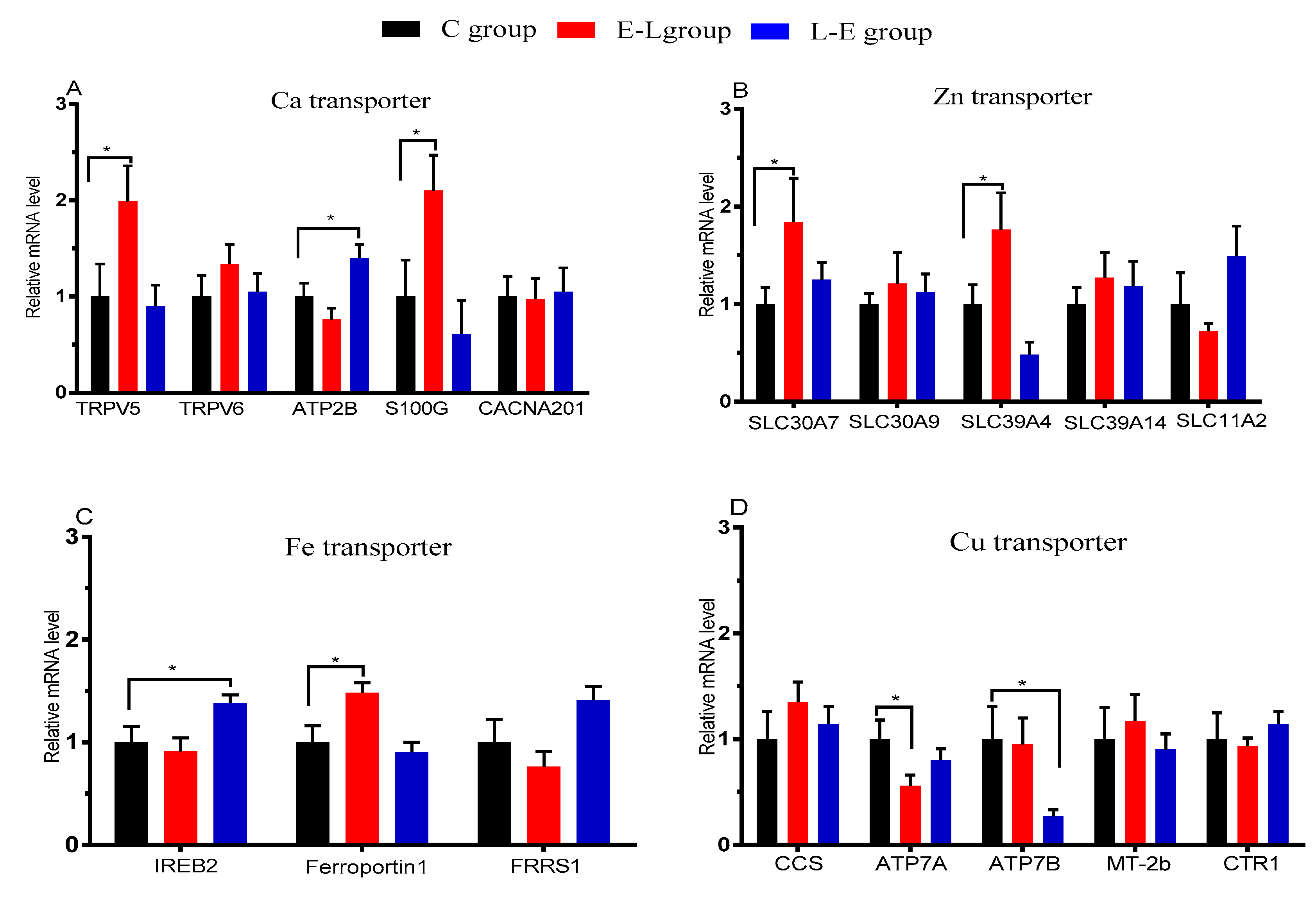
The placenta is an extremely important organ that is responsible for the exchange of nutrients, gases, wastes, and biologically active substances between the maternal and fetal system [
23]. Umbilical serum mineral element concentrations have been used to monitor the fetal mineral element status. Additionally, the transfer of minerals through the placental barrier occurs via several different transporters [
24]. Therefore, it can be inferred that the concentration of minerals in the sow serum, placenta, and umbilical serum are affected directly by a placental mineral transporter expression. In the present study, placental Ca, Fe, Zn, and Cu transporters were affected by Ca feeding time during late pregnancy.
Ca plays an important role in embryo implantation and development, and the functioning of the placenta [
25]. Ca passes through the placental barrier via several different Ca channels and transporters. To date, studies have shown that TRPV6, TRPV5, S100G, and ATP2B1 are expressed in placental trophoblasts in pigs [
26]. One paper reports that serum Ca is correlated with dietary Ca at farrowing [
27], and sows fed the low Ca diet in late gestation will reduce the serum Ca of sows, and the highest serum Ca level is seen in the neonatal piglets from sows fed a high Ca diet at birth [
21]. In addition, feeding Ca at 06:00 and 15:00 during late pregnancy affects mRNA expression of the placental circadian clock in pigs [
13]. Consistent with these observations, the present study shows that the umbilical serum Ca increased significantly in both the E-L and L-E groups. Similarly, sow serum Ca concentrations increased significantly. In contrast, placental Ca concentrations decreased significantly. Consistent with these observations, the mRNA expression levels of Ca transporters in the placenta, including TRPV5, S100G, and ATP2B, were up-regulated significantly in the E-L and L-E groups. Therefore, it is speculated that Ca feeding time to sows increased the utilization of calcium for sows, and Ca feeding time may affect the mRNA expression of placental transporters of Ca by regulating the placental mRNA expression of Period1 (PER1), Period2 (PER2), and Clock gene (CLOCK). Higher umbilical serum Ca might be beneficial to fetal growth and development [
28], and lower placental Ca concentrations may reduce the possibility of placental calcification. Additionally, higher sow serum Ca during delivery may contribute to reducing the risk of hypocalcemic responses. These results may account for the reduction of stillbirths in this study.
Zn is an essential trace mineral and plays a significant role in the metabolism, growth, and development of the body. Two protein families are involved in placental Zn transport, the ZnT proteins (Slc30), and the ZIP proteins (Slc39) [
29]. The present study showed that feeding Ca to sows at 06:00 significantly increased umbilical serum Zn. In contrast, the concentration of Zn in the sow serum and placenta in the E-L group decreased. Consistent with these observations, the mRNA expression of the Zn transporters in the placenta, including Slc39A4 and Slc30A7, were significantly upregulated in the E-L group. Therefore, it was believed that the time of feeding Ca to sows regulated placental Zn transport by regulating the mRNA expression level of transporters. Zn is important to fetal neurological development and is related to the developmental status during infancy. Therefore, increased umbilical serum Zn may contribute to fetal development.
As an essential micronutrient, Fe plays an important role in the development of the placenta and fetus during the fetal and neonatal period. A number of transporters, including transferrin receptor1 (TFR1), DMT1, ZIP14, ferroportin 1 (FPN1), Fe
3+ che/ate reductase 1 (FRRS1), and Fe responsive element binding protein (IRE-BP), are involved in placental Fe transport [
30,
31]. The present study shows that feeding Ca to sows at 06:00 or 15:00 significantly increased the umbilical serum Fe concentration and decreased placental Fe. Consistent with these observations, the mRNA expression of REB2 and FPN1 increased significantly in the L-E and E-L groups, respectively. It was believed that the time of feeding Ca to sows affected umbilical serum Fe by regulating placental mRNA expression of REB2 and FPN1. It is reported that anemia and Fe deficiency during pregnancy are associated with high placental weights and high ratios of placental weight to birth weight [
32]. Therefore, a higher concentration of Fe in the umbilical cord blood may contribute to fetal development.
A series of enzymes and carrier proteins are involved in placental Cu transport. The main role of ATP7A/ATP7B is the catalyzation of adenosine triphosphate (ATP) decomposition to supply energy and promote the excretion of Cu [
33]. This study shows that umbilical serum Cu decreased significantly in both the E-L and L-E groups. Consistent with these observations, placental mRNA expression of Cu transporters, including ATP7A and ATP7B, were significantly down-regulated in the E-L and L-E groups. These results demonstrated that the time of feeding Ca to sows during late pregnancy significantly decreased umbilical serum Cu by down-regulating the placental mRNA expression of ATP7A/ATP7B.
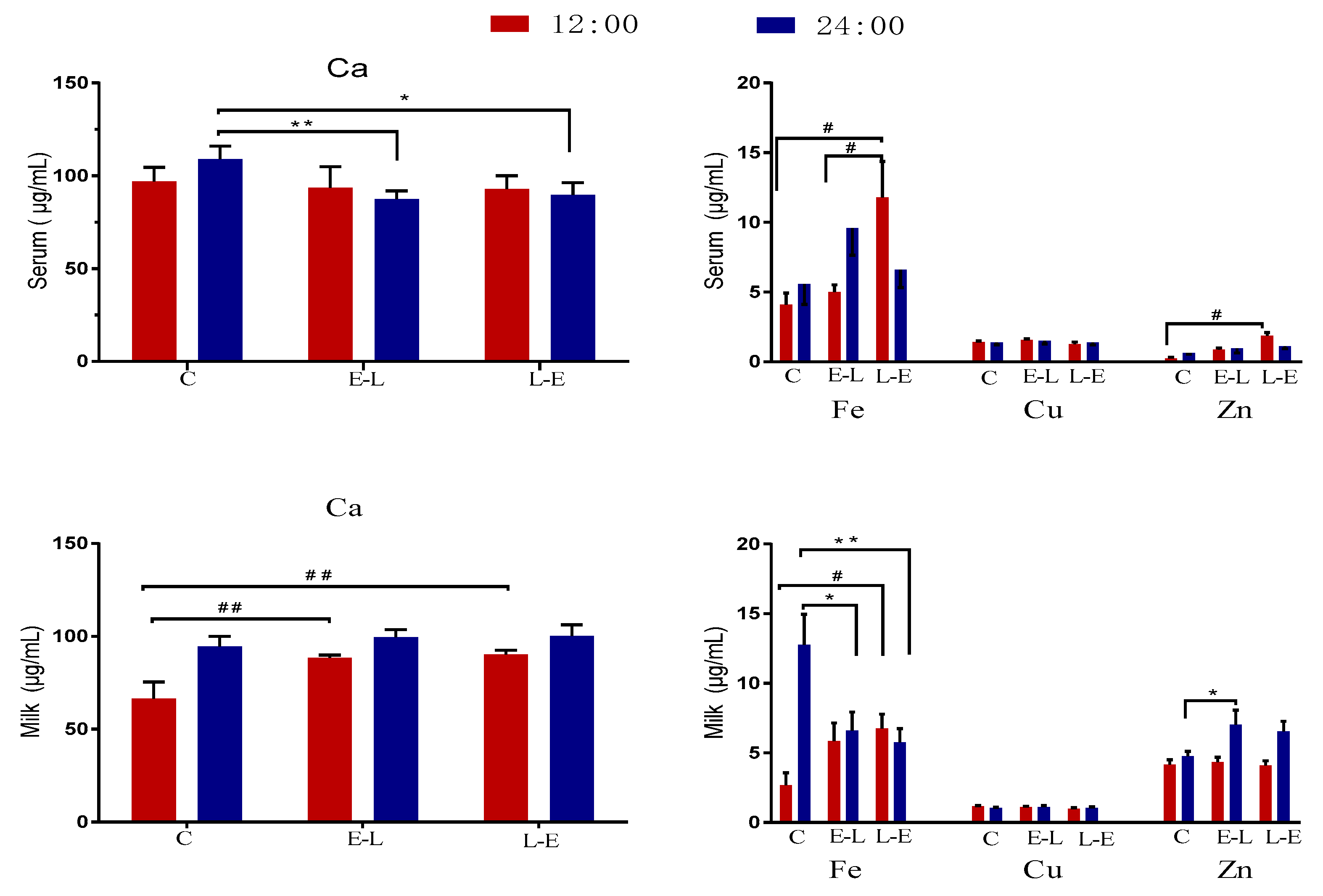
Colostrum synthesis is largely initiated before parturition, and mineral composition of colostrum can be influenced by the mineral needs of the pregnant sow and by dietary mineral supplementation [
3]. In addition, the interrelationships between mineral elements are complex. Research has indicated that increasing Ca intake can reduce the bioavailability of Zn, Cu, and Fe in animals, but that these effects may be dose-dependent [
21]. In the present study, an interesting observation was that colostral Ca decreased significantly in the E-L and L-E groups, although sow serum Ca increased. In contrast, colostral Cu and Zn increased significantly in both the E-L and L-E groups. It is believed that feeding Ca to sows at 06:00 or 15:00 during late pregnancy mainly promotes the utilization of Ca by fetus; however, the specific mechanism needs further research.
There were some changes in the Ca, Fe, and Zn contents of serum and milk observed between the day and night on day 7 of lactation, in the present study. On the one hand, in the present study, milk Ca increased significantly at 12:00 in both the E-L and L-E groups. In contrast, serum Ca decreased significantly at 24:00 in the E-L and L-E groups. In addition, one study shows that there was an upwards trend in the milk Ca of sows fed a high Ca diet, compared to that from other groups [
2]. Therefore, it is believed that maternal Ca feeding at 06:00 or 15:00 may promote the utilization of Ca during the night. On the other hand, in the present study, maternal Ca feeding at 15:00 significantly increased milk Fe during the day and decreased milk Fe during the night. In addition, maternal feeing Ca at 15:00 significantly increased milk Zn during the day and night. While it has been reported that a high Ca diet for two weeks does not inhibit the Fe absorption or alter the Fe status of young piglets [
34], another study found that mRNA expression of genes involved in Fe absorption and metabolism in pigs can be regulated by the time of feeding Fe [
35]. In addition, placental mRNA expression of PER1, PER2, and CLOCK can be affected by Ca feeding time [
13]. Therefore, it was speculated that the gene expression of Ca, Fe, and Zn transporters in mammary gland tissue may be related to PER1, PER2, and CLOCK, and the mutual antagonism between Ca and Zn for their absorption at the mammary epithelial cell may be weakened by changing the time of feeding. For technical reasons, the mRNA expressions of mineral element transporters and of PER1, PER2, and CLOCK in the mammary glands were not measured in the present study and these specific mechanisms need to be further studied.







{kind=link}
{kind=link}
{kind=link}