Are Behavioral Tests Capable of Measuring Positive Affective States in Growing Pigs?

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing Conditions
2.2. Experimental Procedure
2.3. Statistical Analysis
3. Results
3.1. Human Approach Test (HAT)
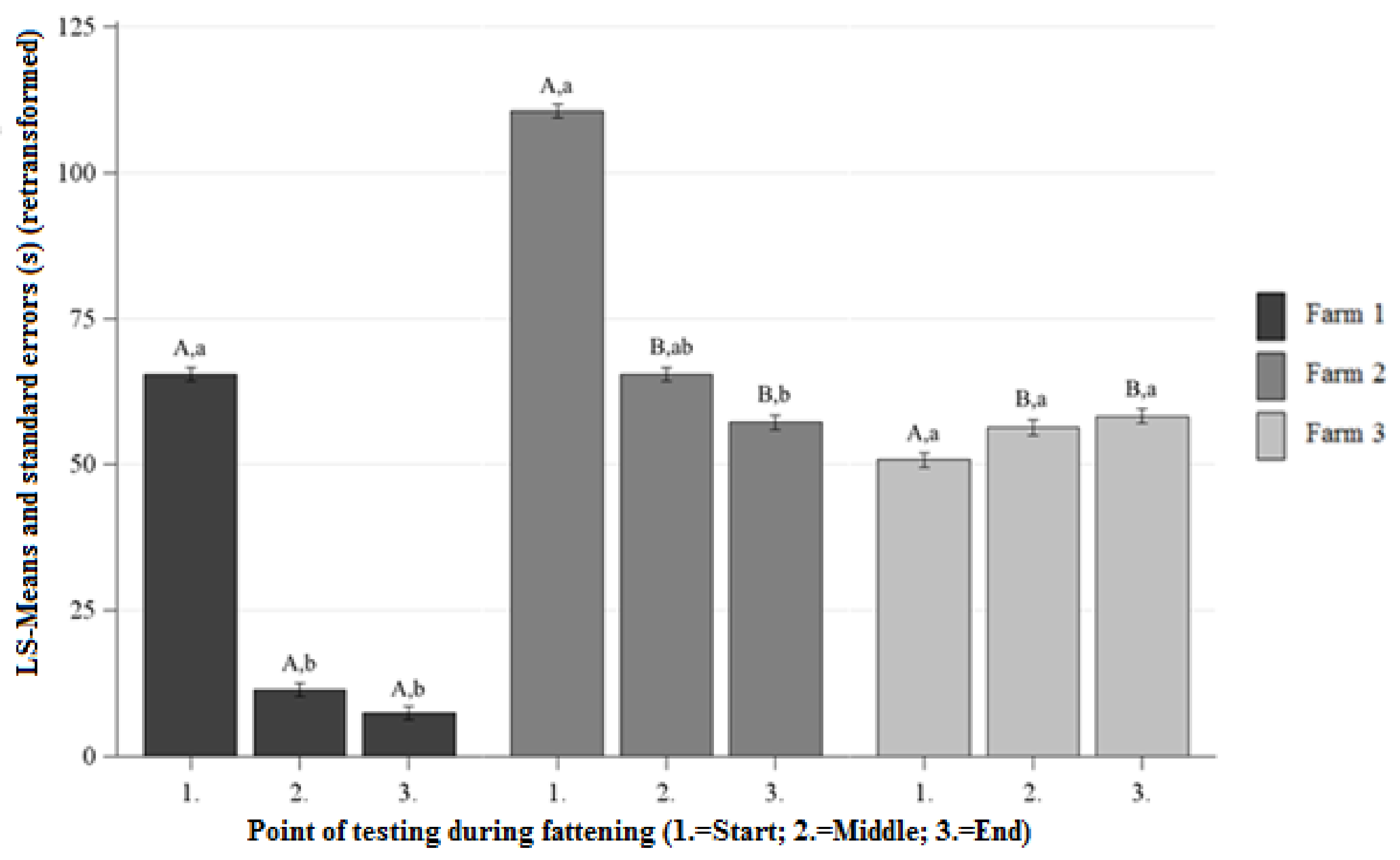
3.1.1. Approach Latency (AL) HAT
3.1.2. Duration of Contact (DC) HAT
Differences Between the Farms at Each Point of Testing
Differences Between the Points of Testing Within Each Farm
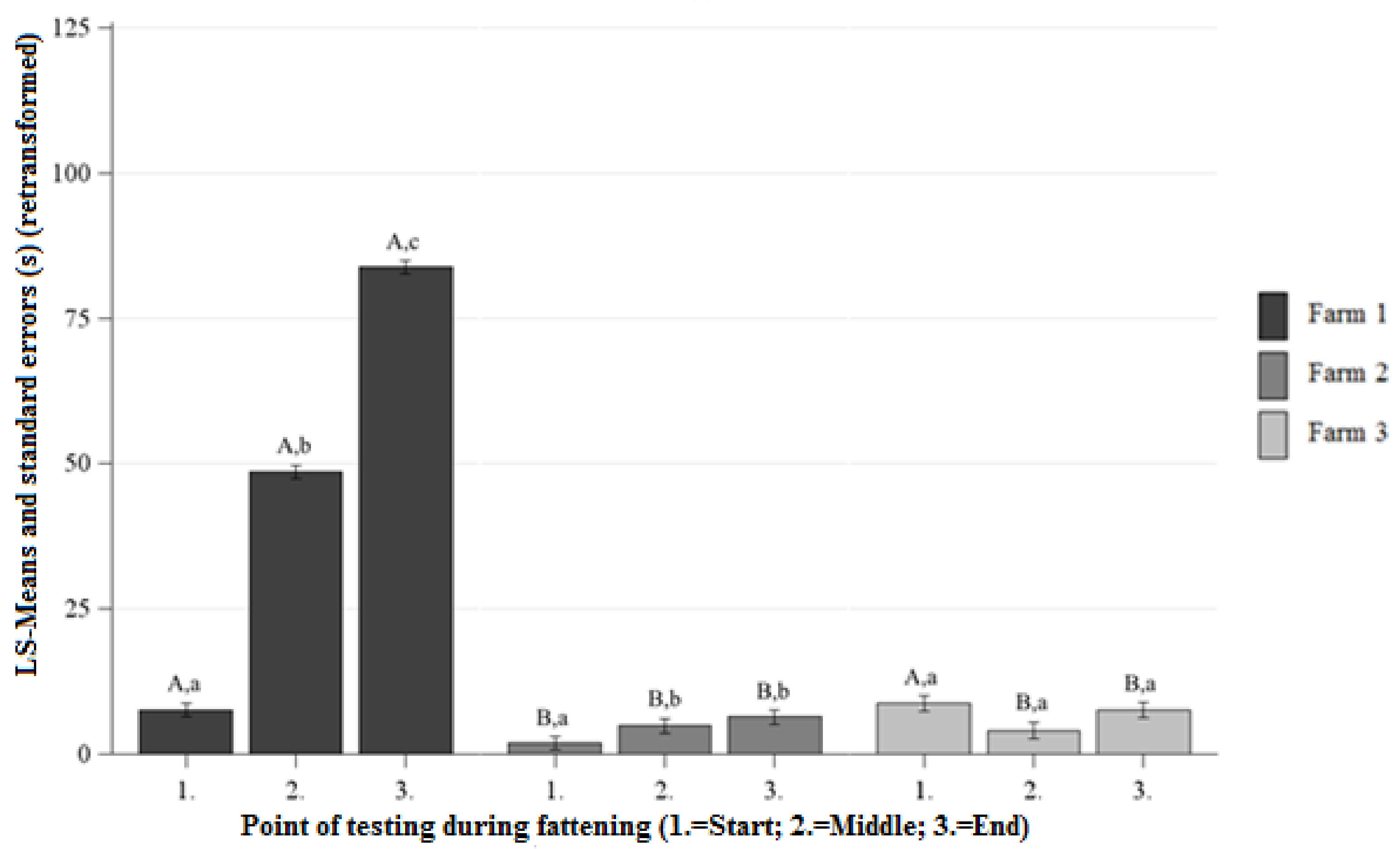
3.1.3. Number of Contacts (NC) HAT
Differences Between the Farms at Each Point of Testing
Differences Between the Points of Testing Within Each Farm
3.1.4. Gender HAT
3.2. Novel Object Test (NOT)
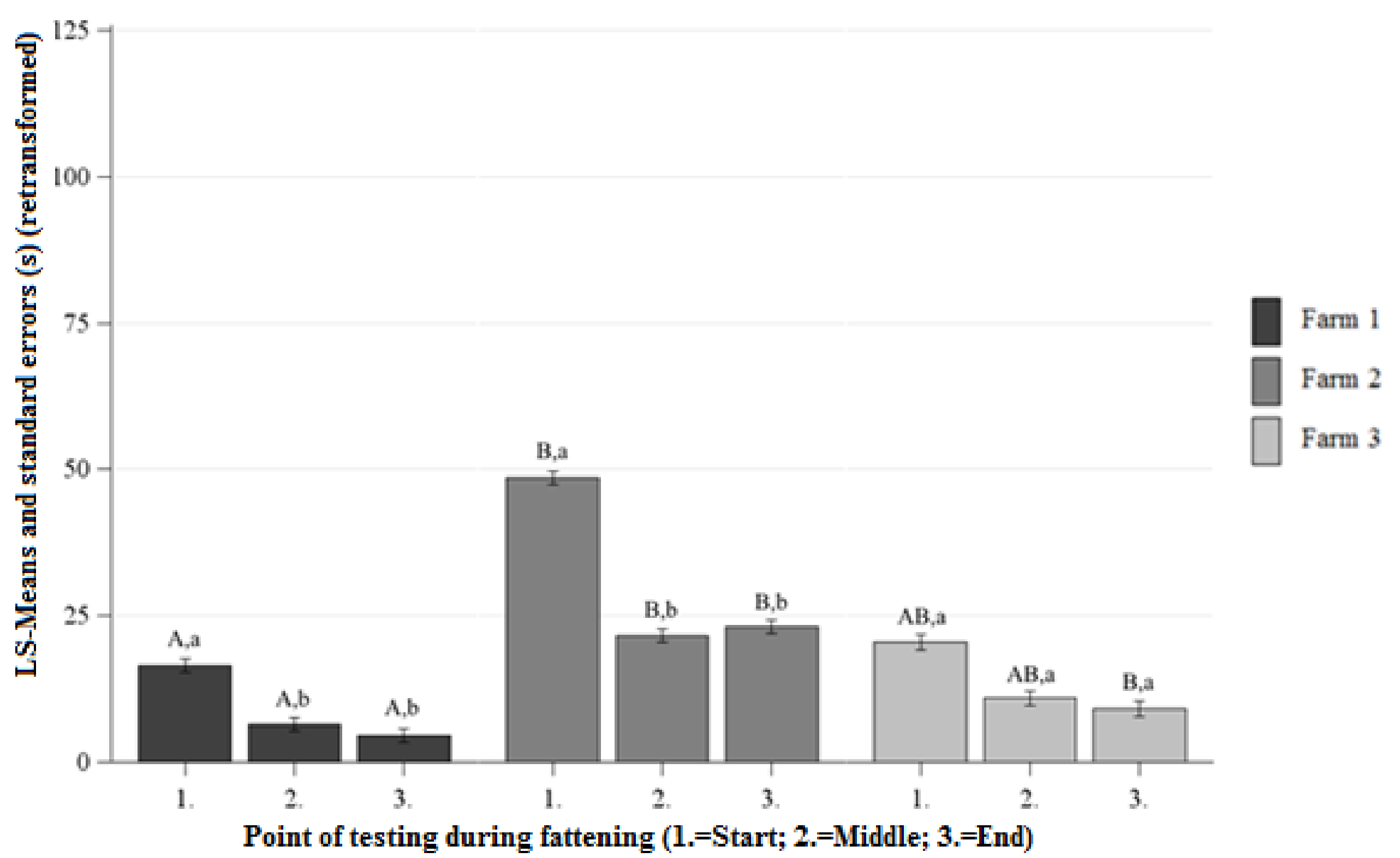
3.2.1. Approach Latency (AL) NOT
Differences between the Farms at Each Point of Testing
Differences between the Points of Testing Within Each Farm
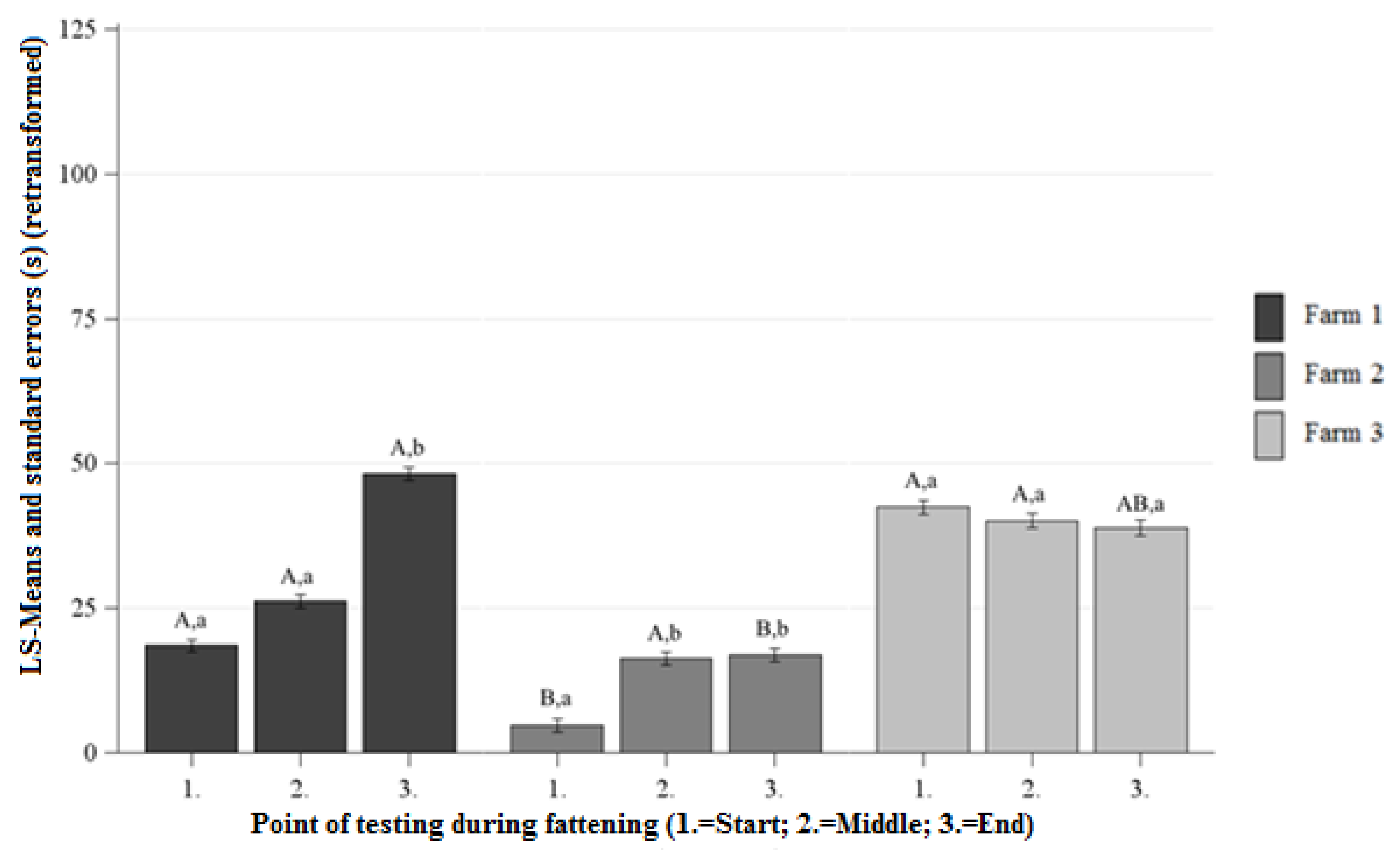
3.2.2. Duration of Contact (DC) NOT
Differences Between the Farms at Each Point of Testing
Differences Between the Points of Testing Within Each Farm
3.2.3. Number of Contacts (NC) NOT
Differences Between the Farms at Each Point of Testing
Differences Between the Points of Testing Within Each Farm
3.2.4. Gender NOT
3.3. Relationship Between the Behavioral Variables
4. Discussion
4.1. Approach Latency (AL)
4.2. Duration of Contacts (DC)
4.3. Number of Contacts (NC)
4.4. Gender
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Forkman, B.; Boissy, A.; Meunier-Salaün, M.-C.; Canali, E.; Jones, R.B. A criticalreview of fear tests used on cattle, pigs, sheep, poultry and horses. Physiol. Behav. 2007, 92, 340–374. [Google Scholar] [CrossRef] [PubMed]
- Wilsson, E.; Sundgren, P.E. The use of a behaviour test for selection of dogs for service and breeding. II. Heritability for tested parameters and effect of selection based on service dog characteristics. Appl. Anim. Behav. Sci. 1997, 54, 235–241. [Google Scholar] [CrossRef]
- Carreras, R.; Arroyo, L.; Mainau, E.; Valent, D.; Bassols, A.; Dalmau, A. Can theway pigs are handled alter behavioural and physiological measures of affective state? Behav. Process. 2017, 142, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.; Nordquist, R.E.; van der Staay, F.J. A review of behavioural methods to study emotion and mood in pigs, Sus scrofa. Appl. Anim. Behav. Sci. 2014, 9–28. [Google Scholar] [CrossRef]
- Hemsworth, P.H.; Coleman, G.J. Human–Livestock Interactions: The Stockperson and the Productivity of Intensively Farmed Animals; CAB International: Wallingford, UK, 1998. [Google Scholar]
- Dalmau, A.; Fabrega, E.; Velarde, A. Fear assessment in pigs exposed to a novelobject test. Appl. Anim. Behav. Sci. 2009, 117, 173–180. [Google Scholar] [CrossRef]
- Boissy, A. Fear and fearfulness in animals. Q. Rev. Biol. 1995, 70, 165–191. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D. Animal behaviour, animal welfare and the scientific study of affect. Appl. Anim. Behav. Sci. 2009, 118, 108–117. [Google Scholar] [CrossRef]
- Czycholl, I.; Kniese, C.; Schrader, L.; Krieter, J. Assessment of the multi-criteriaevaluation system of the Welfare Quality® protocol for growing pigs. Animal 2017, 11, 1573–1580. [Google Scholar] [CrossRef]
- Temple, D.; Dalmau, A.; Ruiz de la Torre, J.; Manteca, X.; Velarde, A. Application of the Welfare Quality protocol to assess growing pigs kept under intensive conditions in Spain. J. Vet. Behav. 2011, 6, 138–149. [Google Scholar] [CrossRef]
- Bokkers, E.A.M.; de Vries, M.; Antonissen, I.; de Boer, I.J.M. Inter- and intra-observer reliability of experienced and inexperienced observers for the Qualitative Behaviour Assessment in dairy cattle. Anim. Welf. 2012, 21, 307–318. [Google Scholar] [CrossRef]
- Tuyttens, F.A.M.; de Graaf, S.; Heerkens, J.L.T.; Jacobs, L.; Nalon, E.; Ott, S.; Stadig, L.; Van Laer, E.; Ampe, B. Oberserver bias in animal behaviour research: Can we believe what we score, if we score what we believe? Anim. Behav. 2014, 90, 273–280. [Google Scholar] [CrossRef]
- Marcet Rius, M.; Cozzi, A.; Bienboire-Frosini, C.; Teruel, E.; Chabaud, C.; Monneret, P. Selection of putative indicators of positive emotions triggered by object and social play in mini-pigs. Appl. Anim. Behav. Sci. 2018, 202, 13–19. [Google Scholar] [CrossRef]
- Carter, A.J.; Marshall, H.H.; Heinsohn, R.; Cowlishaw, G. How not to measure boldness. Novel object and antipredator responses are not the same in wild baboons. Anim. Behav. 2012, 84, 603–609. [Google Scholar] [CrossRef]
- van der Staay, F.J.; van Zutphen, J.A.; de Ridder, M.M.; Nordquist, R.E. Effects of environmental enrichment on decision-making behavior in pigs. Appl. Anim. Behav. Sci. 2017, 194, 14–23. [Google Scholar] [CrossRef]
- Bracke, M.; Spoolder, H.A.M. Novel object test can detect marginal differences in environmental enrichment in pigs. Appl. Anim. Behav. Sci. 2008, 109, 39–48. [Google Scholar] [CrossRef]
- SAS institute Inc.; SAS Institute Inc.: Cary, NC, USA, 2017.
- Brown, J.A.; Dewey, C.; Delange, C.F.M.; Mandell, I.B.; Purslow, P.P.; Robinson, J.A.; Squires, E.J.; Widowski, T.M. Reliability of temperament tests on finishingpigs in group-housing and comparison to social tests. Appl. Anim. Behav. Sci. 2009, 118, 28–35. [Google Scholar] [CrossRef]
- Stolba, A.; Wood-Gush, D.G.M. Arousal and exploration in growing pigs in different environments. Appl. Anim. Behav. Sci. 1980, 6, 382–383. [Google Scholar] [CrossRef]
- Wemelsfelder, F.; Haskell, M.; Mendl, M.T.; Calvert, S.; Lawrence, A.B. Diversity of behaviour during novel object tests is reduced in pigs housed in substrate-impoverished conditions. Anim. Behav. 2000, 60, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Beattie, V.E.; Walker, N.; Sneddon, I.A. Effects of environmental enrichment on behaviour and productivity of growing pigs. Anim. Welf. 1995, 4, 207–220. [Google Scholar]
- Pedersen, L.J.; Damm, B.I.; Kongsted, A.G. The influence of adverse or gentle handling procedures on sexual behaviour in fearful and confident sows. Appl. Anim. Behav. Sci. 2003, 83, 277–290. [Google Scholar] [CrossRef]
- Waiblinger, S.; Boivin, S.; Pedersen, V.; Tosi, M.; Janczak, A.M.; Visser, E.K. Assessing the human animal relationship in farmed species: A critical review. Appl. Anim. Behav. Sci. 2006, 101, 185–242. [Google Scholar] [CrossRef]
- Stolba, A.; Woodgush, D.G.M. Verhaltensgliederung und Reaktion auf Neureize als ethologische Kriterien zur Beurteilung von Haltungsbedingungen bei Hausschweinen. KTBL-Schrift 1981, 264, 110–128. [Google Scholar]
- Boivin, X.; Le Neindre, P.; Chupin, J.M. Establishment of cattle–human relationships. Appl. Anim. Behav. Sci. 1992, 32, 325–335. [Google Scholar] [CrossRef]
- Reimert, I.; Rodenburg, T.B.; Ursinus, W.W.; Kemp, B.; Bolhuis, J.E. Responses to novel situations of female and castrated male pigs with divergent social breedingvalues and different backtest classifications in barren and straw-enriched housing. Appl. Anim. Behav. Sci. 2014, 151, 24–35. [Google Scholar] [CrossRef]
- Jarvis, S.; D’Eath, R.B.; Robson, S.K.; Lawrence, A.B. The effect of confinement during lactation on the hypothalamic-pituitary-adrenal axis and behaviour of primiparous sows. Physiol. Behav. 2006, 87, 345–352. [Google Scholar] [CrossRef]
- Reimert, I.; Rodenburg, T.B.; Ursinus, W.W.; Duijvesteijn, N.; Camerlink, I.; Kemp, B.; Bolhuis, J.E. Backtest and novelty behavior of female and castrated male piglets with diverging social breeding values for growth. J. Anim. Sci. 2013, 91, 4589–4597. [Google Scholar] [CrossRef] [PubMed]
- Prunier, A.; Bonneau, M.; von Borell, E.H.; Cinotti, S.; Gunn, M.; Frederiksen, B.; Giersing, M.; Morton, D.B.; Tuyttens, F.A.M.; Velarde, A. A review of the welfare consequences of surgical castration in piglets and the evaluation of non-surgical methods. Anim. Welf. 2006, 15, 277–289. [Google Scholar]
- Fleming, S.A.; Dilger, R.N. Young pigs exhibit differential exploratory behavior during novelty preference tasks in response to age, sex, and delay. Behav. Brain Res. 2017, 321, 50–60. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioral Variables | Female | Male | p-Value |
|---|---|---|---|
| HAT-AL | 33.36 ± 1.09 a | 52.05 ± 1.11 b | 0.04 |
| HAT-DC | 10.97 ± 1.10 a | 7.75 ± 1.12 b | 0.005 |
| HAT-NC | 1.96 ± 1.03 a | 1.82 ± 1.03 a | 0.05 |
| NOT-AL | 12.52 ± 1.08 a | 16.14 ± 1.09 b | 0.01 |
| NOT-DC | 25.98 ± 1.08 a | 20.95 ± 1.10 b | 0.03 |
| NOT-NC | 3.23 ± 1.03 a | 3.04 ± 1.03 a | 0.1 |
| Behavioral Variables | HAT-DC | HAT-AL | NOT-NC | NOT-DC | NOT-AL |
|---|---|---|---|---|---|
| HAT-NC | 0.76 | −0.62 | 0.16 | 0.21 | −0.16 |
| HAT-DC | −0.68 | 0.16 | 0.24 | −0.19 | |
| HAT-AL | −0.13 | −0.18 | 0.18 | ||
| NOT-NC | 0.74 | −0.55 | |||
| NOT-DC | −0.61 | ||||
| NOT-AL |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krugmann, K.; Warnken, F.; Krieter, J.; Czycholl, I. Are Behavioral Tests Capable of Measuring Positive Affective States in Growing Pigs? Animals 2019, 9, 274. https://doi.org/10.3390/ani9050274
Krugmann K, Warnken F, Krieter J, Czycholl I. Are Behavioral Tests Capable of Measuring Positive Affective States in Growing Pigs? Animals. 2019; 9(5):274. https://doi.org/10.3390/ani9050274
Chicago/Turabian StyleKrugmann, Katja, Farina Warnken, Joachim Krieter, and Irena Czycholl. 2019. "Are Behavioral Tests Capable of Measuring Positive Affective States in Growing Pigs?" Animals 9, no. 5: 274. https://doi.org/10.3390/ani9050274
APA StyleKrugmann, K., Warnken, F., Krieter, J., & Czycholl, I. (2019). Are Behavioral Tests Capable of Measuring Positive Affective States in Growing Pigs? Animals, 9(5), 274. https://doi.org/10.3390/ani9050274