Genome-Wide Association Study of H/L Traits in Chicken

Simple Summary
Abstract
1. Introduction
2. Materials and Method
2.1. Experimental Animals
2.2. Phenotypic Measurement
2.3. Genotyping and Quality Control
2.4. Genome-Wide Association Analysis
2.5. Gene Identification and Annotation
3. Results
3.1. Phenotypic Description and Genetic Parameters
3.2. Population Structure
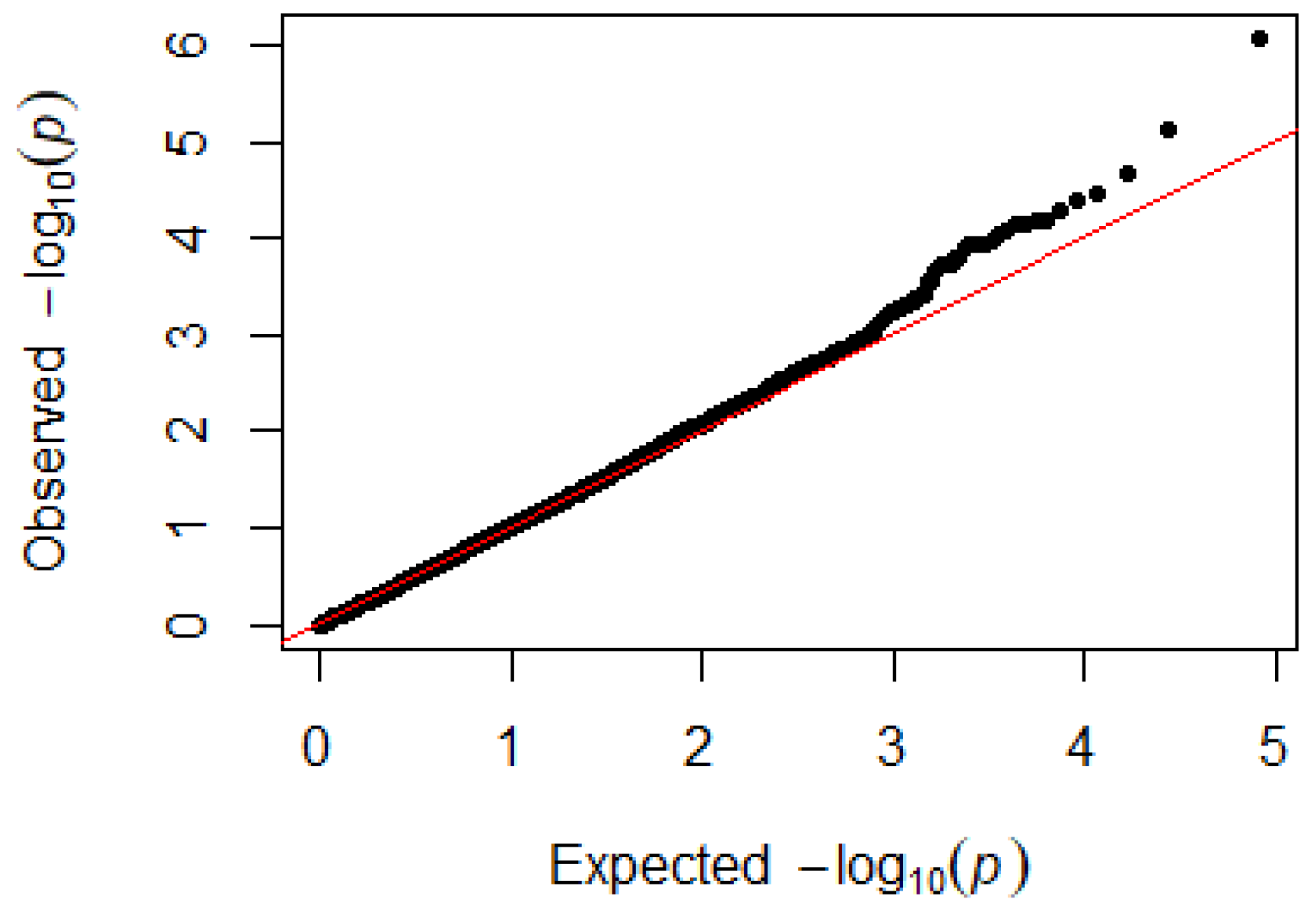
3.3. H/L Ratio
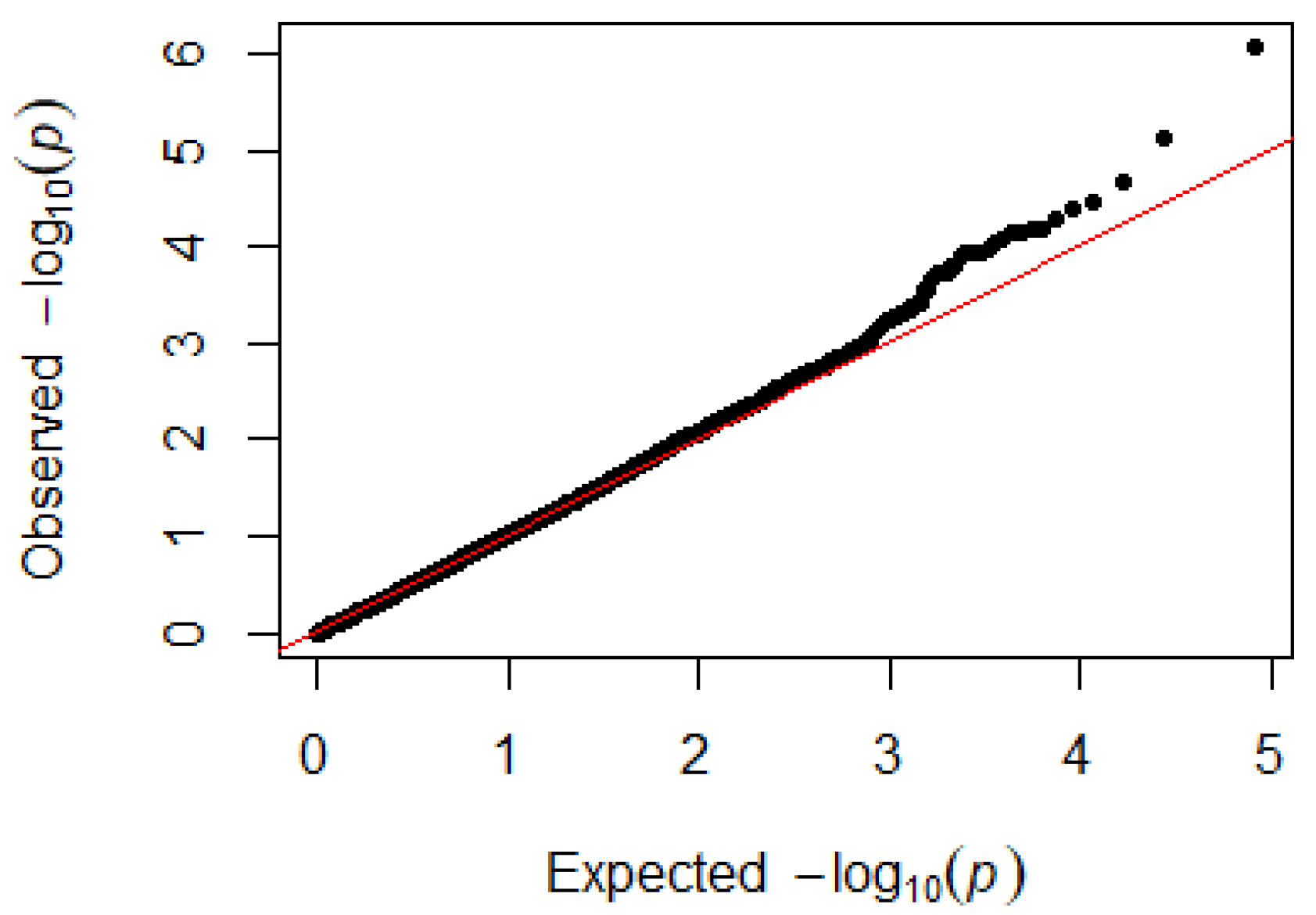
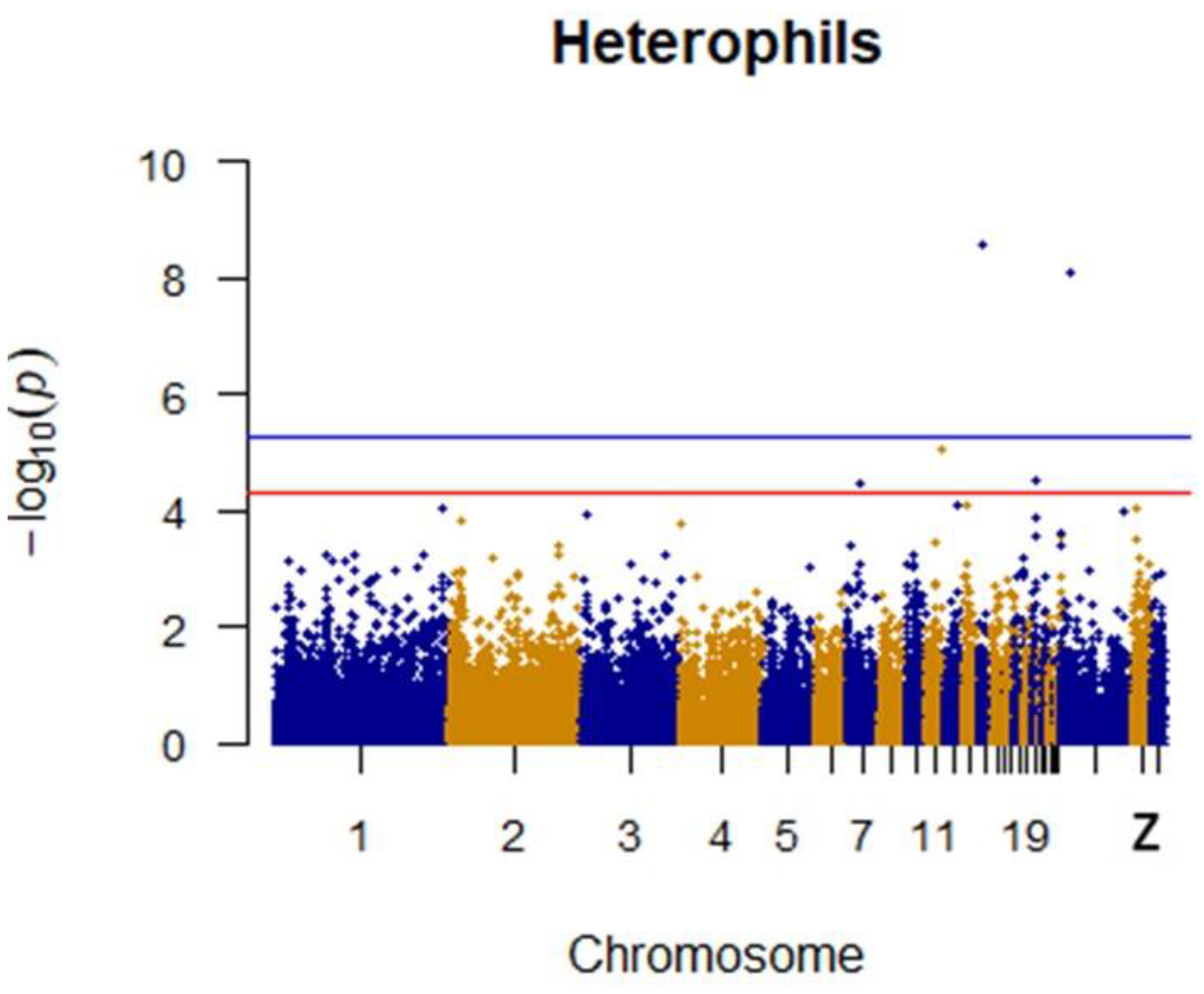
3.4. Heterophils and LYMPHOCYTES
3.5. Monocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sarker, N.; Tsudzuki, M.; Nishibori, M.; Yamamoto, Y. Direct and correlated response to divergent selection for serum immunoglobulin M and G levels in chickens. Poult. Sci. 1999, 78, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gross, W.B.; Siegel, H.S. Evaluation of the heterophil/lymphocyte ratio as a measure of stress in chickens. Avian Dis. 1983, 27, 972–979. [Google Scholar] [CrossRef]
- Stabler, J.G.; McCormick, T.W.; Powell, K.C.; Kogut, M.H. Avian heterophils and monocytes: Phagocytic and bactericidal activities against Salmonella enteritidis. Vet. Microbiol. 1994, 38, 293–305. [Google Scholar] [CrossRef]
- Kogut, M.H.; Hsin-I, C.; Swaggerty, C.L.; Pevzner, I.Y.; Huaijun, Z. Gene expression analysis of toll-like receptor pathways in heterophils from genetic chicken lines that differ in their susceptibility to salmonella enteritidis. Front. Genet. 2012, 3, 121. [Google Scholar] [CrossRef]
- Genovese, K.J.; He, H.; Swaggerty, C.L.; Kogut, M.H. The avian heterophil. Dev. Comp. Immunol. 2013, 41, 334–340. [Google Scholar] [CrossRef]
- Al-Murrani, W.K.; Al-Rawi, I.K.; Raof, N.M. Genetic resistance to Salmonella typhimurium in two lines of chickens selected as resistant and sensitive on the basis of heterophil/lymphocyte ratio. Br. Poult. Sci. 2002, 43, 501–507. [Google Scholar] [CrossRef]
- Dijk, A.V. Chicken heterophils are recruited to the site of Salmonella infection and release antibacterial mature Cathelicidin-2 upon stimulation with LPS. Mol. Immunol. 2009, 46, 1517–1526. [Google Scholar] [CrossRef] [PubMed]
- Kogut, M.H. Expression and function of Toll-like receptors in chicken heterophils. Dev. Comp. Immunol. 2005, 29, 791–807. [Google Scholar] [CrossRef]
- Maxwell, M.H.; Robertson, G.W. The avian heterophil leucocyte: A review. World’s Poult. Sci. J. 1998, 54, 155–178. [Google Scholar] [CrossRef]
- Dehnhard, N.; Quillfeldt, P.; Hennicke, J.C. Leucocyte profiles and H/L ratios in chicks of Red-tailed Tropicbirds reflect the ontogeny of the immune system. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2011, 181, 641–648. [Google Scholar] [CrossRef]
- Lentfer, T.L.; Pendl, H.; Gebhardt-Henrich, S.G.; FroHlich, E.K.F.; Von Borell, E. H/L ratio as a measurement of stress in laying hens—Methodology and reliability. Br. Poult. Sci. 2015, 56, 157–163. [Google Scholar] [CrossRef]
- Zhang, L.; Li, P.; Liu, R.R.; Zheng, M.Q.; Sun, Y.; Wu, D.; Zhao, G.P.; Wen, J. The identification of loci for immune traits in chickens using a genome-wide association study. PLoS ONE 2015, 10, e0117269. [Google Scholar] [CrossRef]
- Bayona, J.A.M.; Karuppannan, A.K.; Barreda, D.R. Contribution of leukocytes to the induction and resolution of the acute inflammatory response in chickens. Dev. Comp. Immunol. 2017, 74, 167–177. [Google Scholar] [CrossRef]
- Pieper, J.; Locke, M.; Ruzaike, G.; Voigt, S.; Methner, U.; Berndt, A. In vitro and in vivo generation of heterophil extracellular traps after Salmonella exposure. Vet. Immunol. Immunopathol. 2017, 188, 1–11. [Google Scholar] [CrossRef]
- Al-Murrani, W.K.; Kassab, A.; Al-Sam, H.Z.; Al-Athari, A.M.K. Heterophil/lymphocyte ratio as a selection criterion for heat resistance in domestic fowls. Br. Poult. Sci. 1997, 38, 159–163. [Google Scholar] [CrossRef]
- Mutibvu, T.; Chimonyo, M.; Halimani, T. Tonic immobility, heterophil to lymphocyte ratio, and organ weights in slow-growing chickens. J. Appl. Poult. Res. 2017, 26, 226–235. [Google Scholar] [CrossRef]
- Scanes, C.G. Biology of stress in poultry with emphasis on glucocorticoids and the heterophil to lymphocyte ratio. Poult. Sci. 2016, 95, 2208–2215. [Google Scholar] [CrossRef]
- Clark, P. Observed variation in the heterophil to lymphocyte ratio values of birds undergoing investigation of health status. Comp. Clin. Pathol. 2015, 24, 1151–1157. [Google Scholar] [CrossRef]
- Fidan, E.D.; Nazligül, A.; Türkyilmaz, M.K.; Karaarslan, S.; Kaya, M. Effects of photoperiod length and light intensity on performance, carcass characteristics and heterophil to lymphocyte ratio in broilers. Kafkas Univ. Vet. Fak. Derg. 2017, 23, 39–45. [Google Scholar]
- Wang, W.; Zhang, T.; Wang, J.; Zhang, G.; Wang, Y.; Zhang, Y.; Zhang, J.H.; Li, G.H.; Xue, Q.; Han, K.P.; et al. Genome-wide association study of 8 carcass traits in Jinghai Yellow chickens using specific-locus amplified fragment sequencing technology. Poult. Sci. 2016, 95, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation plink: rising to the challenge of larger and richer datasets. Giga Sci. 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Toddbrown, K.; Thomas, L.; Ferreira, M.; Bender, D. Plink: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Stephens, M. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar]
- McLaren, W.; Pritchard, B.; Rios, D.; Chen, Y.; Flicek, P.; Cunningham, F. Deriving the consequences of genomic variants with the ensembl API and SNP effect predictor. Bioinformatics 2010, 26, 2069–2070. [Google Scholar] [CrossRef]
- Kasprzyk, A. Biomart: Driving a paradigm change in biological data management. Database 2011, 1, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, R.J.; Kähäri, A.; Haider, S.; Zamora, J.; Proctor, G.; Spudich, G.; Almeida-King, J.; Staines, D.; Derwent, P.; Kersey, A.K.P.; et al. Ensembl BioMarts: A hub for data retrieval across taxonomic space. Database 2011, 2011, bar030. [Google Scholar] [CrossRef] [PubMed]
- Mcfarlane, J.M.; Curtis, S.E. Multiple concurrent stressors in chicks. 3. Effects on plasma corticosterone and the heterophil:lymphocyte ratio. Poult. Sci. 1989, 68, 522–527. [Google Scholar] [CrossRef]
- Gross, W.B.; Curtis, S.E. Factors affecting chicken thrombocyte morphology and the relationship with heterophil: Lymphocyte ratios. Br. Poult. Sci. 1989, 30, 919–925. [Google Scholar] [CrossRef]
- Campo, J.L.; Davila, S.G. Estimation of heritability for heterophil: Lymphocyte ratio in chickens by restricted maximum likelihood. Effects of age, sex, and crossing. Poult. Sci. 2002, 81, 1448–1453. [Google Scholar] [CrossRef]
- Gross, O.; Grupp, C.; Steinberg, C.; Zimmermann, S.; Strasser, D.; Hannesschläger, N.; Reindl, W.; Jonsson, H.; Huo, H.; Littman, D.R.; et al. Multiple ITAM-coupled NK-cell receptors engage the Bcl10/Malt1 complex via Carma1 for NF-κB and MAPK activation to selectively control cytokine production. Blood 2008, 112, 2421–2428. [Google Scholar] [CrossRef]
- Georg, L. Oncogenic CARD11 mutations in human diffuse large B cell lymphoma. Science 2008, 319, 1676–1679. [Google Scholar]
- Bertin, J.; Wang, L.; Guo, Y.; Jacobson, M.D.; Poyet, J.L.; Srinivasula, S.M. CARD11 and CARD14 are novel caspase recruitment domain (CARD)/membrane-associated guanylate kinase (MAGUK) family members that interact with BCL10 and activate NF-kappa B. J. Biol. Chem. 2001, 276, 11877–11882. [Google Scholar] [CrossRef]
- Pomerantz, J.L.; Denny, E.M.; David, B. CARD11 mediates factor-specific activation of NF-kappaB by the T cell receptor complex. EMBO J. 2014, 21, 5184–5194. [Google Scholar] [CrossRef]
- Gaide, O.; Martinon, F.; Micheau, O.; Bonnet, D.; Thome, M.; Tschopp, J. Carma1, a CARD-containing binding partner of Bcl10, induces Bcl10 phosphorylation and NF-κB activation 1. FEBS Lett. 2001, 496, 121–127. [Google Scholar] [CrossRef]
- Siwek, M.; Slawinska, A.; Rydzanicz, M.; Wesoly, J.; Fraszczak, M.; Suchocki, T. Identification of candidate genes and mutations in qtl regions for immune responses in chicken. Anim. Genet. 2015, 46, 247–254. [Google Scholar] [CrossRef]
- Slawinska, A.; Witkowski, A.; Bednarczyk, M.; Siwek, M. In silico analysis of candidate genes associated with humoral innate immune response in chicken. BMC Proc. 2011, 5, S36. [Google Scholar] [CrossRef]
- Feng, L.; Yang, X.; Asweto, C.O.; Wu, J.; Zhang, Y.; Hu, H. Genome-wide transcriptional analysis of cardiovascular-related genes and pathways induced by PM2.5 in human myocardial cells. Environ. Sci. Pollut. Res. 2017, 24, 11683–11693. [Google Scholar] [CrossRef]
- Zhu, M.; Xu, Y.; Chen, Y.; Yan, F. Circular BANP, an upregulated circular RNA that modulates cell proliferation in colorectal cancer. Biomed. Pharmacother. 2017, 88, 138–144. [Google Scholar] [CrossRef]
- Li, S.S.; Qu, Z.; Haas, M.; Ngo, L.; Heo, Y.J.; Kang, H.J.; Britto, M.J.; Cullen, H.D.; Vanyai, H.K.; Tan, S.S.; et al. The HSA21 gene EURL/C21ORF91 controls neurogenesis within the cerebral cortex and is implicated in the pathogenesis of Down Syndrome. Sci. Rep. 2016, 6, 29514. [Google Scholar] [CrossRef]
- Fu, W.Y.; Chen, Y.; Sahin, M.; Zhao, X.S.; Shi, L.; Bikoff, J.B. Cdk5 regulates EphA4-mediated dendritic spine retraction through an ephexin1-dependent mechanism. Nat. Neurosci. 2007, 10, 67–76. [Google Scholar] [CrossRef]
- Town, L.; Mcglinn, E.; Davidson, T.L.; Browne, C.M.; Chawengsaksophak, K.; Koopman, P. Tmem26 is dynamically expressed during palate and limb development but is not required for embryonic survival. PLoS ONE 2011, 6, e25228. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Shi, Q.; Wei, J.; Zhou, W.; Xiao, K.; Wang, J. The associations of two SNPs in miRNA-146a and one SNP in ZBTB38-RASA2 with the disease susceptibility and the clinical features of the Chinese patients of sCJD and FFI. Prion 2018, 12, 34–41. [Google Scholar] [CrossRef]
- Nass, N.; Dittmer, A.; Hellwig, V.; Lange, T.; Beyer, J.M.; Leyh, B.; Ignatov, A.; Weiβenborn, C.; Kirkegaard, T.; Lykkesfeldt, A.E.; et al. Expression of transmembrane protein 26 (TMEM26) in breast cancer and its association with drug response. Oncotarget 2016, 7, 38408–38426. [Google Scholar] [CrossRef]
- Barr, A.J.; Ugochukwu, E.; Lee, W.H.; King, O.N.F.; Filippakopoulos, P.; Alfano, I. Large-scale structural analysis of the classical human protein tyrosine phosphatome. Cell 2009, 136, 352–363. [Google Scholar] [CrossRef]
- Shah, T.M.; Patel, N.V.; Patel, A.B.; Upadhyay, M.R.; Joshi, C.G. A genome-wide approach to screen for genetic variants in broilers (gallus gallus) with divergent feed conversion ratio. Mol. Genet. Genom. 2016, 291, 1715–1725. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, M.; Ni, Y.H.; Liu, F.; Fan, H.Q.; Fei, L.; Pan, X.Q.; Guo, M.; Chen, R.H.; Guo, X.R. Identification and characterization of NYGGF4, a novel gene containing a phosphotyrosine-binding (PTB) domain that stimulates 3T3-L1 preadipocytes proliferation. Gene 2006, 379, 132–140. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Mean | SD | Deviation | Min | Max | CV1 (%) |
|---|---|---|---|---|---|---|
| Monocytes (M, n) | 3 | 3 | 7 | 0 | 17 | 87 |
| Heterophils (H, n) | 27 | 9 | 88 | 2 | 64 | 34 |
| Lymphocytes (L, n) | 73 | 18 | 333 | 21 | 88 | 25 |
| H /L (H/L, %) | 40 | 20 | 0 | 0 | 1.7 | 50 |
| Traits | Chromosome | SNP ID | Position 1 (BP) | p_Wald | Nearest Gene | Distance 2 |
|---|---|---|---|---|---|---|
| H/L | 14 | rs15005639 | 3332117 | 1.55 × 10−9 | CARD11 | D103.6 |
| H/L | Z | rs314642216 | 10570600 | 1.59 × 10−9 | BRIX1 | D3.4 |
| H/L | 11 | rs14028611 | 18280341 | 2.27 × 10−5 | BANP | within |
| H/L | 12 | New | 12911436 | 4.16 × 10−5 | PTPRG | within |
| H/L | 7 | rs312628231 | 15894092 | 4.32 × 10−5 | NFE2L2 | D10.6 |
| Traits | Chromosome | SNP ID | Position 1 (BP) | p_Wald | Nearest Gene | Distance 2 |
|---|---|---|---|---|---|---|
| Heterophils | 14 | rs15005639 | 3332117 | 2.54 × 10−9 | CARD11 | D103.6 |
| Heterophils | Z | rs314642216 | 10570600 | 8.06 × 10−9 | BRIX1 | D3.4 |
| Heterophils | 11 | rs14028611 | 18280341 | 8.81 × 10−6 | BANP | within |
| Heterophils | 3 | rs316444238 | 3269267 | 3.13 × 10−5 | PIK3CD | U192.3 |
| Heterophils | 2 | rs312628231 | 15894092 | 3.52 × 10−5 | NFE2L2 | D10.6 |
| Lymphocytes | Z | rs314642216 | 10570600 | 5.88 × 10−6 | BRIX1 | U3.4 |
| Lymphocytes | 1 | New | 1.02E+08 | 9.87 × 10−6 | C1H21ORF91 | U2524.9 |
| Lymphocytes | 14 | rs15005639 | 3332117 | 2.14 × 10−5 | CARD11 | D103.6 |
| Lymphocytes | 21 | New | 12911436 | 4.30 × 10−5 | PTPRG | within |
| Traits | Chromosome | SNP ID | Position 1 (BP) | p_wald | Nearest Gene | Distance 2 |
|---|---|---|---|---|---|---|
| Monocytes | 9 | New | 7719173 | 8.62 × 10−7 | EPHA4 | within |
| Monocytes | 6 | rs313943680 | 8556872 | 7.59 × 10−6 | TMEM26 RHOBTB1 | within U87.7 |
| Monocytes | 9 | New | 10113376 | 2.08 × 10−5 | RASA2 | U100.5 |
| PID1 | U189.3 | |||||
| Monocytes | 9 | New | 8301605 | 3.42 × 10−5 | WDFY1 | within |
| Monocytes | 5 | rs313016555 | 44638604 | 4.01 × 10−5 | CPSF2 | within |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, B.; Li, Q.; Liu, R.; Zheng, M.; Wen, J.; Zhao, G. Genome-Wide Association Study of H/L Traits in Chicken. Animals 2019, 9, 260. https://doi.org/10.3390/ani9050260
Zhu B, Li Q, Liu R, Zheng M, Wen J, Zhao G. Genome-Wide Association Study of H/L Traits in Chicken. Animals. 2019; 9(5):260. https://doi.org/10.3390/ani9050260
Chicago/Turabian StyleZhu, Bo, Qinghe Li, Ranran Liu, Maiqing Zheng, Jie Wen, and Guiping Zhao. 2019. "Genome-Wide Association Study of H/L Traits in Chicken" Animals 9, no. 5: 260. https://doi.org/10.3390/ani9050260
APA StyleZhu, B., Li, Q., Liu, R., Zheng, M., Wen, J., & Zhao, G. (2019). Genome-Wide Association Study of H/L Traits in Chicken. Animals, 9(5), 260. https://doi.org/10.3390/ani9050260