Waste Disposal Sites as All-You-Can Eat Buffets for Carrion Crows (Corvus corone)


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
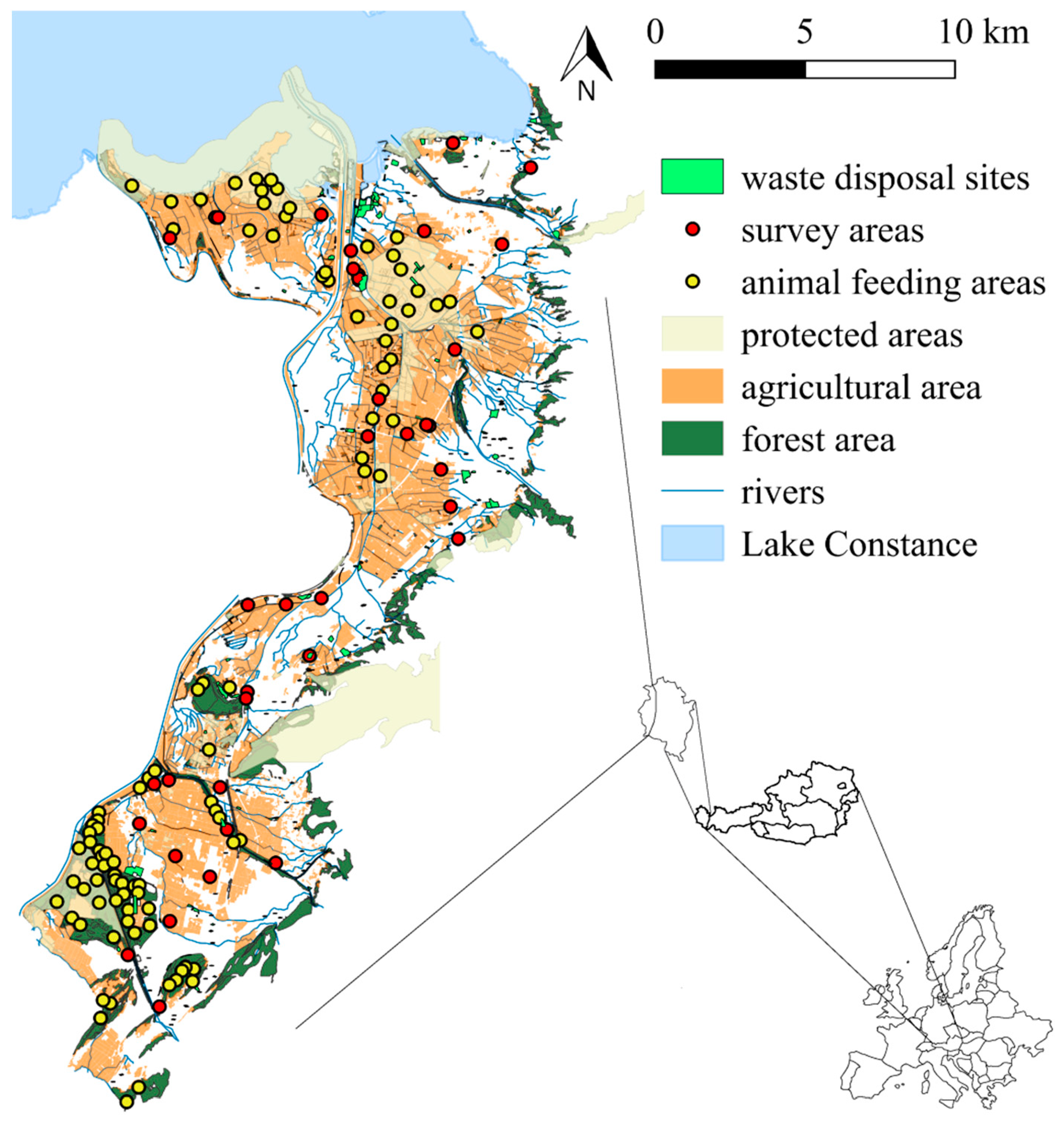
2.2. Study Species and Site
2.3. Data Collection and Analysis
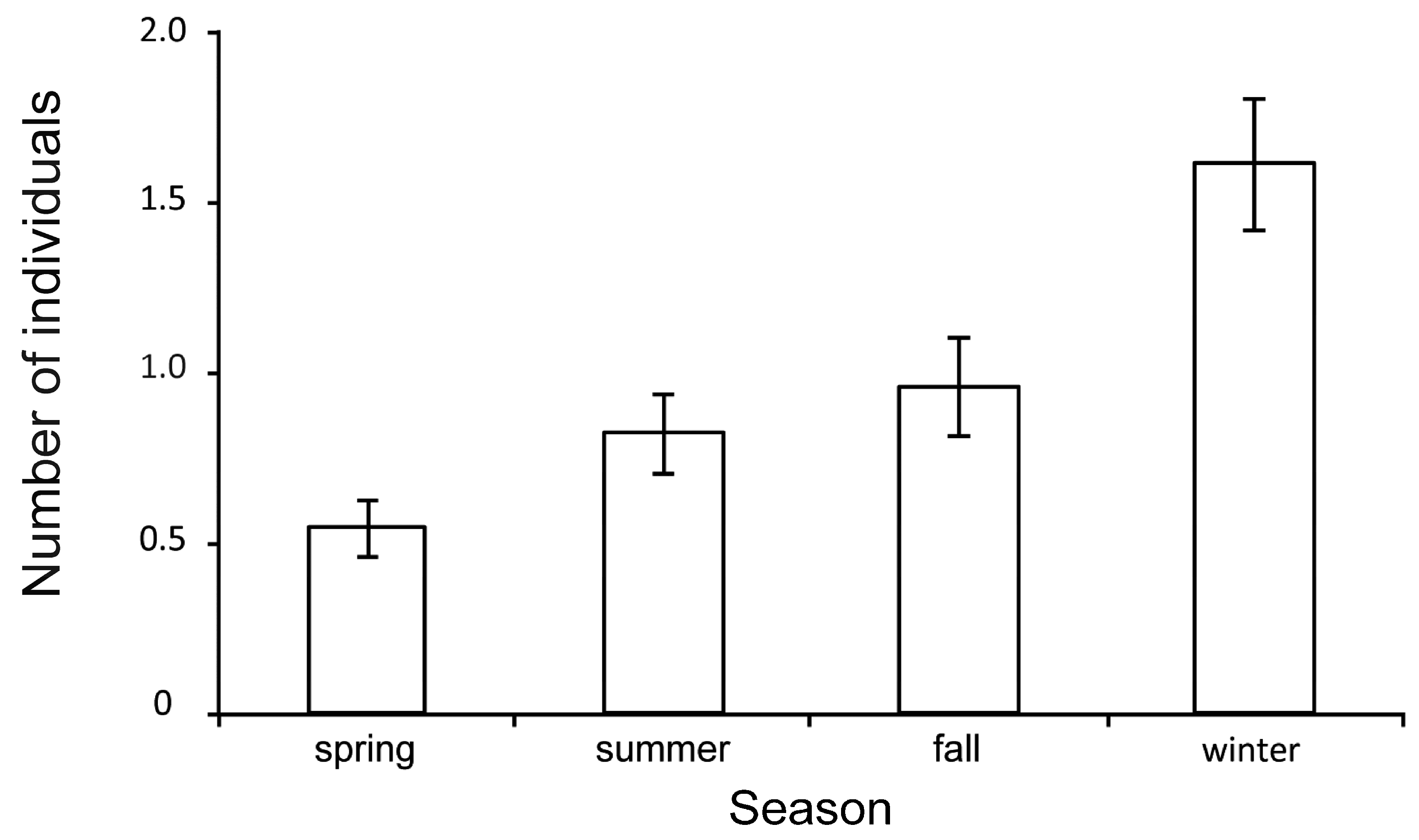
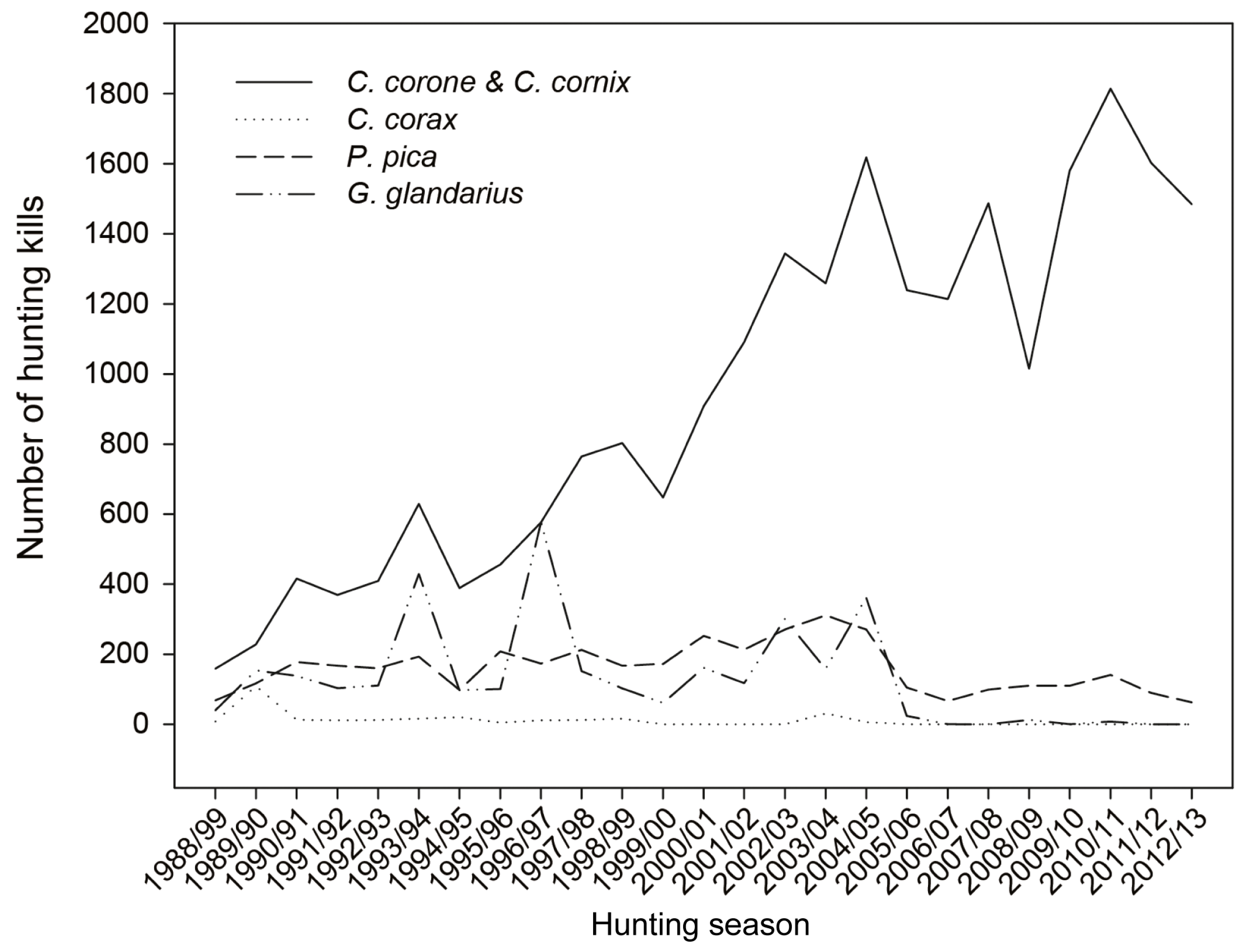
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Kövér, L.; Gyüre, P.; Balogh, P.; Huettmann, F.; Lengyel, S.; Juhász, L. Recent colonization and nest site selection of the Hooded Crow (Corvus corone cornix L.) in an urban environment. Landsc. Urban Plan. 2015, 133, 78–86. [Google Scholar] [CrossRef]
- Marzluff, J.M.; Neatherlin, E. Corvid response to human settlements and campgrounds: Causes, consequences, and challenges for conservation. Biol. Conserv. 2006, 130, 301–314. [Google Scholar] [CrossRef]
- Marzluff, J.M.; McGowan, K.J.; Donnelly, R.; Knight, R.L. Causes and consequences of expanding American Crow populations. In Avian Ecology and Conservation in an Urbanizing World; Springer: Berlin, Germany, 2001; pp. 331–363. [Google Scholar]
- Kristan, W.B., III; Boarman, W.I.; Crayon, J.J. Diet composition of common ravens across the urban-wildland interface of the West Mojave Desert. Wildl. Soc. Bull. 2004, 32, 244–253. [Google Scholar] [CrossRef]
- Tratalos, J.; Fuller, R.A.; Evans, K.L.; Davies, R.G.; Newson, S.E.; Greenwood, J.J.; Gaston, K.J. Bird densities are associated with household densities. Glob. Chang. Biol. 2007, 13, 1685–1695. [Google Scholar] [CrossRef]
- Withey, J.C.; Marzluff, J.M. Multi-scale use of lands providing anthropogenic resources by American Crows in an urbanizing landscape. Landsc. Ecol. 2009, 24, 281. [Google Scholar] [CrossRef]
- Withey, J.C.; Marzluff, J.M.; Brittingham, M. Dispersal by juvenile American crows (Corvus brachyrhynchos) influences population dynamics across a gradient of urbanization. Auk 2005, 122, 205–221. [Google Scholar] [CrossRef]
- Marzluff, J.M.; Withey, J.C.; Whittaker, K.A.; David Oleyar, M.; Unfried, T.M.; Rullman, S.; DeLap, J. Consequences of habitat utilization by nest predators and breeding songbirds across multiple scales in an urbanizing landscape. Condor 2007, 109, 516–534. [Google Scholar] [CrossRef]
- Helb, H. Wissenschaftliche Begleituntersuchung an Elster (Pica pica) und Rabenkrähe (Corvus c. corone) in Rheinland-Pfalz. Pollichia 1999, 15, 6–10. [Google Scholar]
- Ramseier, H.; Vonlanthen-Rentsch, I. Krähenschäden im Kanton Bern. 2006. Available online: https://www.vol.be.ch/vol/de/index/landwirtschaft/landwirtschaft/publikationen.assetref/dam/documents/VOL/LANAT/de/Landwirtschaft/PUB_LANAT_LW_Kraehenschaeden_de.pdf (accessed on 30 April 2019).
- Bugnyar, T.; Kotrschal, K. Movement coordination and signalling in ravens (Corvus corax): An experimental field study. Acta Ethol. 2001, 3, 101–109. [Google Scholar] [CrossRef]
- Wright, J.; Stone, R.E.; Brown, N. Communal roosts as structured information centres in the raven, Corvus corax. J. Anim. Ecol. 2003, 72, 1003–1014. [Google Scholar] [CrossRef]
- Sonerud, G.; Hansen, H.; Smedshaug, C. Individual roosting strategies in a flock-living bird: Movement and social cohesion of hooded crows (Corvus corone cornix) from pre-roost gatherings to roost sites. Behav. Ecol. Sociobiol. 2002, 51, 309–318. [Google Scholar] [CrossRef]
- Boeckle, M.; Szipl, G.; Bugnyar, T. Who wants food? Individual characteristics in raven yells. Anim. Behav. 2012, 84, 1123–1130. [Google Scholar] [CrossRef]
- Sonerud, G.A.; Smedshaug, C.A.; Bråthen, Ø. Ignorant hooded crows follow knowledgeable roost-mates to food: Support for the information centre hypothesis. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2001, 268, 827–831. [Google Scholar] [CrossRef]
- JNCC. Directive 2009/147/EC on the Conservation of Wild Birds (Codified Version). Available online: http://jncc.defra.gov.uk/page-1373 (accessed on 28 February 2019).
- Zduniak, P.; Kuczyński, L. Breeding biology of the hooded crow Corvus corone cornix in Warta River Valley (W Poland). Acta Ornithol. 2003, 38, 143–150. [Google Scholar] [CrossRef][Green Version]
- Bivand, R.; Rundel, C. Rgeos: Interface to Geometry Engine-Open Source (GEOS). R Package ver. 0.3–3. 2013. Available online: https://rdrr.io/cran/rgeos/ (accessed on 1 March 2019).
- Vuorisalo, T.; Andersson, H.; Hugg, T.; Lahtinen, R.; Laaksonen, H.; Lehikoinen, E. Urban development from an avian perspective: Causes of hooded crow (Corvus corone cornix) urbanisation in two Finnish cities. Landsc. Urban Plan. 2003, 62, 69–87. [Google Scholar] [CrossRef]
- Charles, J.K. Territorial Behavior and the Limitation of Population Size in Crows Corvus Corone and Corvus Cornix. Ph.D. Thesis, University of Aberdeen, Aberdeen, Scotland, 1972. [Google Scholar]
- Bui, T.V.D.; Marzluff, J.M.; Bedrosian, B. Common raven activity in relation to land use in western wyoming: Implications for greater sage-grouse reproductive success. Condor 2010, 112, 65–78. [Google Scholar] [CrossRef]
- Service, U.S. Human Influences on Predators of Nesting Birds on the North Slope of Alaska; Fish and Wildlife Service: Fairbanks, AK, USA, 2003. [Google Scholar]
- Boarman, W.I. Managing a subsidized predator population: Reducing common raven predation on desert tortoises. Environ. Manag. 2003, 32, 205–217. [Google Scholar] [CrossRef]
- Boarman, W.I.; Patten, M.A.; Camp, R.J.; Collis, S.J. Ecology of a population of subsidized predators: Common ravens in the central Mojave Desert, California. J. Arid Environ. 2006, 67, 248–261. [Google Scholar] [CrossRef]
- Jagdverordnung. In LGBl. Nr. 24/1995 10. Stück. Available online: http://www.ris.bka.gv.at/ (accessed on 1 March 2019).
- Crook, J.H.; Ward, P. The Quelea Problem in Africa. In The Problems of Birds as Pests (Proceedings of a Symposium Held at the Royal Geographical Society London); Murton, R.K., Wright, E.N., Eds.; Academic Press: London, UK; New York, NY, USA, 1968. [Google Scholar]
- Feare, C.J. Ecological studies of the rook (Corvus frugilegus L.) in north-east scotland. Damage and its control. J. Appl. Ecol. 1974, 11, 897–914. [Google Scholar] [CrossRef]
- Bomford, M. Review of Research on Control of Bird Pests in Australia; University of Nebraska-Lincoln: Lincoln, NE, USA, 1992; pp. 93–96. [Google Scholar]
- Reichholf, J.H. Rabenschwarze Intelligenz: Was wir von Krähen Lernen können; Piper Verlag GmbH: München, Germany, 2011. [Google Scholar]
- Baglione, V.; Marcos, J.M.; Canestrari, D. Cooperatively breeding groups of carrion crow (Corvus corone corone) in northern Spain. Auk 2002, 119, 790–799. [Google Scholar]
- Bedrosian, B. Nesting and post-fledging ecology of the common raven in Grand Teton National Park, Wyoming. Master’s Thesis, Arkansas State University, Arkansas, AR, USA, 2005. [Google Scholar]
- Braun, A.; Walsdorff, T.; Fraser, O.N.; Bugnyar, T. Socialized sub-groups in a temporary stable Raven flock? J. Ornithol. 2012, 153, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, B.; Kaye, D.; Knight, T.; Schaumburg, K. Dispersal and association among common ravens. Condor 1994, 96, 545–551. [Google Scholar] [CrossRef]
- Heinrich, B.; Marzluff, J.; Marzluff, C. Common ravens are attracted by appeasement calls of food discoverers when attacked. Auk 1993, 110, 247–254. [Google Scholar]
- Boeckle, M.; Szipl, G.; Bugnyar, T. Raven food calls indicate sender’s age and sex. Front. Zool. 2018, 15, 5. [Google Scholar] [CrossRef] [PubMed]
- Szipl, G.; Boeckle, M.; Wascher, C.A.F.; Spreafico, M.; Bugnyar, T. With whom to dine? Ravens’ responses to food-associated calls depend on individual characteristics of the caller. Anim. Behav. 2015, 99, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, B.; Marzluff, J. Do common ravens yell because they want to attract others? Behav. Ecol. Sociobiol. 1991, 28, 13–21. [Google Scholar] [CrossRef]
- Bugnyar, T.; Kijne, M.; Kotrschal, K. Food calling in ravens: Are yells referential signals? Anim. Behav. 2001, 61, 949–958. [Google Scholar] [CrossRef]
- Loretto, M.C.; Reimann, S.; Schuster, R.; Graulich, D.M.; Bugnyar, T. Shared space, individually used: Spatial behaviour of non-breeding ravens (Corvus corax) close to a permanent anthropogenic food resource. J. Ornithol. 2016, 157, 339–450. [Google Scholar] [CrossRef]
- Epple, W. Zum Schutz der Rabenvögel; Niedersächsisches Landesamt für Ökologie, Abt. Naturschutz: Hannover, Germany, 1997. [Google Scholar]
- Glutz von Blotzheim, U.; Bauer, K.M. Handbuch der Vögel Mitteleuropas: Sittidae—Laniidae; Aula-Verlag: Wiesbaden, Germany, 1993. [Google Scholar]
- Parkin, D.T.; Collinson, M.; Helbig, A.J.; Knox, A.G.; Sangster, G. The taxonomic status of Carrion and Hooded Crows. Br. Birds 2003, 96, 274–290. [Google Scholar]
- Saino, N. Selection of foraging habitat and flocking by crow Corvus corone phenotypes in a hybrid zone. Ornis Scand. 1992, 23, 111–120. [Google Scholar] [CrossRef]
- Landesamt für Vermessung und Geoinformation Feldkirch. Geographical Data from the State of Vorarlberg. Available online: http://vogis.cnv.at (accessed on 18 April 2016).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Pebesma, E.J.; Bivand, R.S. Classes and methods for spatial data in R. R News 2005, 5, 9–13. [Google Scholar]
- Bivand, R.S.; Pebesma, E.J.; Gómez-Rubio, V.; Pebesma, E.J. Applied Spatial Data Analysis with R; Springer: Berlin, Germany, 2013; Volume 10. [Google Scholar]
- Keitt, T.H.; Bivand, R.; Pebesma, E.; Rowlingson, B. Rgdal: Bindings for the Geospatial Data Abstraction Library. Available online: https://r-forge.r-project.org/projects/rgdal/ (accessed on 30 April 2019).
- Hijmans, R.J. Introduction to the ”Geosphere” Package, Version 1.3-11. 2014. Available online: https://mran.microsoft.com/snapshot/2014-10-25/web/packages/geosphere/vignettes/geosphere.pdf (accessed on 30 April 2019).
- Bivand, R.; Wong, D.W.S. Comparing implementations of global and local indicators of spatial association. Test 2018, 27, 716–748. [Google Scholar] [CrossRef]
- Fortin, M.; Dale, M. Spatial autocorrelation. In The SAGE Handbook of Spatial Analysis; Fotheringham, A.S., Rogerson, P.A., Eds.; SAGE Publications: London, UK, 2008. [Google Scholar]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in {R}. Bioinformatics 2018, 35, 526–528. [Google Scholar] [CrossRef]
- Bartoń, K. MuMIn: Multi-Model Inference; R Package Version 1.43.6. Available online: https://cran.r-project.org/web/packages/MuMIn/MuMIn.pdf (accessed on 30 April 2019).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 2nd ed.; Sage: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multi-Model Inference: A Practical Information-Theoretic Approach; Springer: London, UK, 2010. [Google Scholar]
- Bauer, H.G.; Heine, G.; Bodensee-Brutvogelatlas 2000; Herausgegeben von der Ornithologischen Arbeitsgemeinschaft Bodensee (OAB). Stand Oktober 2005. Available online: https://www.bodensee-ornis.de/avifaunistik/brutvogelkartierung/ (accessed on 1 March 2019).
- Teufelbauer, N.; Seaman, B.S.; Dvorak, M. Bestandsentwicklungen Häufiger Österreichischer Brutvögel im Zeitraum 1998–2016—Ergebnisse des Brutvogel-Monitoring. Egretta 2017, 55, 43–76. [Google Scholar]
- Mayer, G. Die Kolkraben im Windischgarstner Becken. Jahresbuch Mus. 1986, 131, 157–171. [Google Scholar]
- Gereben, B.; Wolf, B.; Krenn, H. Der Bestand der Saatkrähen an den großen Winterschlafplätzen in Wien 1994/95. Vogelkundl. Nachr. Ostösterreich 1995, 6, 43–46. [Google Scholar]
- Wolf, B.; Krenn, H.W.; Gereben-Krenn, B.A. Der Bestand der Saatkrähen an den großen Winterschlafplätzen in Wien im Winter 1996/97. Vogelkdl. Beob. Ostösterreich 1997, 8, 71–74. [Google Scholar]
- Loretto, M.C.; Schuster, R.; Bugnyar, T. GPS tracking of non-breeding ravens reveals the importance of anthropogenic food sources during their dispersal in the Eastern Alps. Cur. Zool. 2016, 62, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Ekanayake, K.B.; Sutherland, D.R.; Dann, P.; Weston, M.A. Out of sight but not out of mind: Corvids prey extensively on eggs of burrow-nesting penguins. Wildl. Res. 2015, 42, 509–517. [Google Scholar] [CrossRef]
- Stork, H.J. Zur Historie der Überwinterung russischer Krähen in Berlin. Vogelwarte 2011, 49, 238. [Google Scholar]
- Bezzel, E. Corvus corone L. 1758-Aaskrähe. In Kompendium der Vögel Mitteleuropas; AULA-Verlag GmbH: Wiesbaden, Germany, 1993. [Google Scholar]
- Bossema, I.; Benus, R.F. Territorial defence and intra-pair cooperation in the carrion crow (Corvus corone). Behav. Ecol. Sociobiol. 1985, 16, 99–104. [Google Scholar] [CrossRef]
- Tompa, S. A preliminary investigation of the carrion crow Corvus corone problem in Switzerland. Ornithol. Beob. 1975, 72, 181–198. [Google Scholar]
- Heinrich, B. Mind of the Raven: Investigations and Adventures with Wolf-Birds; Cliff Street Books: New York, NY, USA, 1999. [Google Scholar]
- Richner, H. Habitat-specific growth and fitness in carrion crows (Corvus corone corone). J. Anim. Ecol. 1989, 58, 427–440. [Google Scholar] [CrossRef]
- Group, E.E.I. Composite European Commission Report on the Derogations in 2008 According to Article 9 of Directive 79/409/EEC on the Conservation of Wild Birds. Available online: http://ec.europa.eu/environment/nature/knowledge/rep_birds/docs/derogation_report_2008.pdf (accessed on 30 April 2019).
- Heinrich, B. Why do ravens fear their food? Condor 1988, 90, 950–952. [Google Scholar] [CrossRef]
- Patterson, I. Territorial behaviour and the limitation of population density. Ardea 1980, 68, 53–62. [Google Scholar]
- Efford, M.; Warburton, B.; Spencer, N. Home-range changes by brushtail possums in response to control. Wildl. Res. 2000, 27, 117–127. [Google Scholar] [CrossRef]
- Yom-Tov, Y. The effect of food and predation on breeding density and success, clutch size and laying date of the crow (Corvus corone L.). J. Anim. Ecol. 1974, 43, 479–498. [Google Scholar] [CrossRef]
- Whisson, D.A.; Weston, M.A.; Shannon, K. Home range, habitat use and movements by the little raven (Corvus mellori) in a coastal peri-urban landscape. Wildl. Res. 2015, 42, 500–508. [Google Scholar] [CrossRef]
- Matsubara, H. Comparative study of territoriality and habitat use in syntopic Jungle Crow (Corvus macrorhynchos) and Carrion Crow (C. corone). Ornithol. Sci. 2003, 2, 103–111. [Google Scholar] [CrossRef]
- Baltensperger, A.P.; Mullet, T.C.; Schmid, M.S.; Humphries, G.R.W.; Kover, L.; Huettmann, F. Seasonal observations and machine-learning-based spatial model predictions for the common raven (Corvus corax) in the urban, sub-arctic environment of Fairbanks, Alaska. Polar Biol. 2013, 36, 1587–1599. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Intercept | Predictors | df | logLik | AICc | delta | weight | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 220 | 1944 | 1/ | 4/ | 6/ | 7/ | 8/ | 9 | −9875.68 | 19,769.60 | 0.00 | 0.43 | |||
| 224 | 1878 | 1/ | 3/ | 4/ | 6/ | 7/ | 8/ | 10 | −9875.17 | 19,770.60 | 1.03 | 0.26 | ||
| 252 | 1949 | 1/ | 4/ | 5/ | 6/ | 7/ | 8/ | 10 | −9875.42 | 19,771.10 | 1.54 | 0.20 | ||
| 256 | 1886 | 1/ | 2/ | 3/ | 4/ | 5/ | 6/ | 7/ | 8/ | 11 | −9874.97 | 19,772.20 | 2.68 | 0.11 |
| Parameter | Estimate | Unconditional SE | CI | Relative Importance |
|---|---|---|---|---|
| (Intercept) | 1.921 | 0.070 | (1.784, 2.05) | |
| Capacity value | 0.001 | 0.000 | (0.00081, 0.00089) | 1.00 |
| Season 2 * | −0.279 | 0.031 | (−0.340, −0.218) | 1.00 |
| Season 3 | 0.229 | 0.027 | (0.175, 0.283) | 1.00 |
| Season 4 | −0.680 | 0.035 | (−0.750, −0.611) | 1.00 |
| Distance to feeding area | 0.00004 | 0.000 | (0.000024, 0.000059) | 1.0 |
| Auto-covariation | 0.006 | 0.002 | (0.0020, 0.0094) | 1.0 |
| Number or waste disposal sites | −0.011 | 0.001 | (−0.0125, −0.0095) | 1.0 |
| Number of animal feeding sites | 0.021 | 0.002 | (0.018, 0.025) | 1.0 |
| Daytime | 0.00003 | 0.000 | (−0.000088, 0.000268) | 0.37 |
| Distance to lake of Constance | −0.0000003 | 0.000 | (−0.0000034, 0.0000017) | 0.31 |
| Survey Point Number/Description | Number of Crows/Site | |||
|---|---|---|---|---|
| Mean | SE | Minimum | Maximum | |
| ID4/Biogas | 19.0 | 6.7 | 9.5 | 37.9 |
| ID9/Biogas | 88.2 | 27.6 | 47.7 | 163.2 |
| ID11/Biogas | 14.1 | 5.4 | 6.6 | 29.9 |
| ID26/Biogas | 56.3 | 15.0 | 33.3 | 95.1 |
| ID27/Transfer station | 46.6 | 16.0 | 23.7 | 91.5 |
| ID29/Green waste | 16.8 | 5.4 | 9.0 | 31.5 |
| ID32/Biogas | 15.4 | 5.3 | 7.8 | 30.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Preininger, D.; Schoas, B.; Kramer, D.; Boeckle, M. Waste Disposal Sites as All-You-Can Eat Buffets for Carrion Crows (Corvus corone). Animals 2019, 9, 215. https://doi.org/10.3390/ani9050215
Preininger D, Schoas B, Kramer D, Boeckle M. Waste Disposal Sites as All-You-Can Eat Buffets for Carrion Crows (Corvus corone). Animals. 2019; 9(5):215. https://doi.org/10.3390/ani9050215
Chicago/Turabian StylePreininger, Doris, Bjoern Schoas, Diether Kramer, and Markus Boeckle. 2019. "Waste Disposal Sites as All-You-Can Eat Buffets for Carrion Crows (Corvus corone)" Animals 9, no. 5: 215. https://doi.org/10.3390/ani9050215
APA StylePreininger, D., Schoas, B., Kramer, D., & Boeckle, M. (2019). Waste Disposal Sites as All-You-Can Eat Buffets for Carrion Crows (Corvus corone). Animals, 9(5), 215. https://doi.org/10.3390/ani9050215