Pigeon and Poultry Breeders, Friends or Enemies of the Northern Goshawk Accipiter gentilis? A Long-Term Study of a Population in Central Poland


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
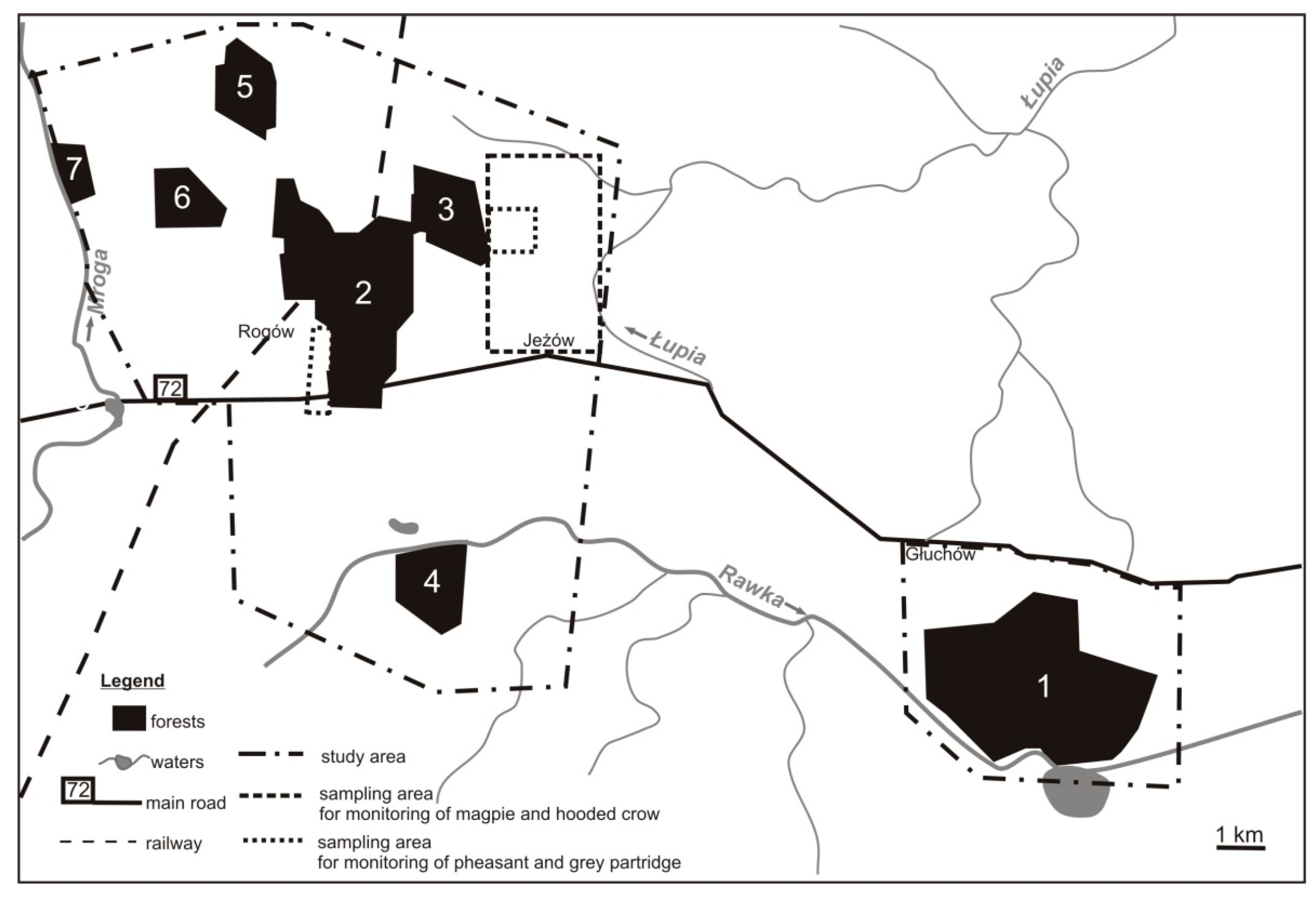
2.1. Study Area
2.2. Inventory of Goshawk Population and Measurements of Nesting Trees
2.3. Diet Composition
2.4. Food Base Assessment
2.5. Statistical Analysis
3. Results
3.1. Changes in the Population Density
3.2. Breeding Parameters
3.3. Characteristics of Nesting Trees
3.4. Food Composition
3.5. Prey Availability
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Prey | N | Body Mass | Total Biomass | Biomass | Prey Items |
|---|---|---|---|---|---|
| g | % | ||||
| Columba livia f. domestica | 243 | 400 | 97200 | 45.9 | 22.8 |
| Gallus domesticus | 13 | 500 | 6500 | 3.1 | 1.2 |
| Numidia meleagris | 1 | 500 | 500 | 0.2 | 0.1 |
| ∑ Anthropogenic birds | 257 | 104200 | 49.2 | 24.1 | |
| Columba palumbus | 97 | 475 | 46075 | 21.8 | 9.1 |
| Streptopelia decaocto | 10 | 225 | 2250 | 1.1 | 0.9 |
| Streptopelia turtur | 1 | 150 | 150 | 0.1 | 0.1 |
| Columba oenas | 1 | 250 | 250 | 0.1 | 0.1 |
| Phasianus colchicus | 2 | 500 | 1000 | 0.5 | 0.2 |
| Coturnix coturnix | 1 | 100 | 100 | 0.0 | 0.1 |
| Anas platyrhynchos | 1 | 500 | 500 | 0.2 | 0.1 |
| Accipiter nissus | 1 | 200 | 200 | 0.1 | 0.1 |
| Vanellus vanellus | 1 | 280 | 280 | 0.1 | 0.1 |
| Scolopax rusticola | 1 | 300 | 300 | 0.1 | 0.1 |
| Chroicocephalus ridibundus | 1 | 300 | 300 | 0.1 | 0.1 |
| Cuculus canorus | 1 | 120 | 120 | 0.1 | 0.1 |
| Asio otus | 1 | 250 | 250 | 0.1 | 0.1 |
| Strix aluco | 1 | 500 | 500 | 0.2 | 0.1 |
| Dendropcopos major | 29 | 70 | 2030 | 1.0 | 2.7 |
| Dendrocopos spp. | 9 | 70 | 630 | 0.3 | 0.8 |
| Driocopus martius | 2 | 300 | 600 | 0.3 | 0.2 |
| Motacilla alba | 2 | 24 | 48 | 0.0 | 0.2 |
| Turdus philomelos | 58 | 50 | 2900 | 1.4 | 5.4 |
| Turdus merula | 34 | 70 | 2380 | 1.1 | 3.2 |
| Turdus pilastris | 15 | 70 | 1050 | 0.5 | 1.4 |
| Turdus viscivorus | 7 | 80 | 560 | 0.3 | 0.7 |
| Turdus spp. | 40 | 65 | 2600 | 1.2 | 3.8 |
| Phylloscopus spp. | 2 | 8 | 16 | 0.0 | 0.2 |
| Ficedula spp. | 4 | 15 | 60 | 0.0 | 0.4 |
| Parus major | 5 | 18 | 90 | 0.0 | 0.5 |
| Paridae spp. | 4 | 20 | 80 | 0.0 | 0.4 |
| Sitta europea | 18 | 22 | 396 | 0.2 | 1.7 |
| Lanius sp. | 4 | 50 | 200 | 0.1 | 0.4 |
| Garrulus glandarius | 140 | 175 | 24500 | 11.6 | 13.1 |
| Pica pica | 3 | 200 | 600 | 0.3 | 0.3 |
| Oriolus oriolus | 1 | 73 | 73 | 0.0 | 0.1 |
| Sturnus vulgaris | 30 | 80 | 2400 | 1.1 | 2.8 |
| Passer spp. | 4 | 25 | 100 | 0.0 | 0.4 |
| Fringilla coelebs | 12 | 20 | 240 | 0.1 | 1.1 |
| Chloris chloris | 3 | 26 | 78 | 0.0 | 0.3 |
| Coccothraustes coccothraustes | 29 | 55 | 1595 | 0.8 | 2.7 |
| Loxia sp. | 4 | 35 | 140 | 0.1 | 0.4 |
| Emberiza citrinella | 4 | 30 | 120 | 0.1 | 0.4 |
| Small bird unident. | 27 | 25 | 675 | 0.3 | 2.5 |
| Medium bird unident. | 40 | 75 | 3000 | 1.4 | 3.8 |
| ∑ Aves | 907 | 203636 | 96.2 | 85.2 | |
| Talpa europea | 2 | 80 | 160 | 0.1 | 0.2 |
| Apodemus agrarius | 4 | 24 | 96 | 0.0 | 0.4 |
| Apodemus spp. | 9 | 27 | 243 | 0.1 | 0.8 |
| Rattus norvegicus | 3 | 200 | 600 | 0.3 | 0.3 |
| Myodes glareolus | 28 | 18 | 504 | 0.2 | 2.6 |
| Microtus spp. | 6 | 22 | 132 | 0.1 | 0.6 |
| Rodentia unident. | 45 | 20 | 900 | 0.4 | 4.2 |
| Sciurus vulgaris | 3 | 300 | 900 | 0.4 | 0.3 |
| Oryctolagus cuniculus f. dom. | 3 | 200 | 600 | 0.3 | 0.3 |
| Lepus europeus juv. | 1 | 100 | 100 | 0.0 | 0.1 |
| Felis catus | 9 | 200 | 1800 | 0.9 | 0.8 |
| Mustela sp. | 1 | 45 | 45 | 0.0 | 0.1 |
| Vulpes vulpes | 1 | 200 | 200 | 0.1 | 0.1 |
| Canis familiaris | 3 | 200 | 600 | 0.3 | 0.3 |
| Sus domestica | 3 | 200 | 600 | 0.3 | 0.3 |
| Capreolus capreolus | 2 | 200 | 400 | 0.2 | 0.2 |
| ∑ Mammalia | 123 | 7880 | 3.7 | 11.5 | |
| Rana sp. | 1 | 15 | 15 | 0.0 | 0.1 |
| Natrix natrix | 1 | 100 | 100 | 0.0 | 0.1 |
| Insecta unident. | 33 | 1 | 33 | 0.0 | 3.1 |
| Total | 1065 | 211,664 | 100.0 | 100.0 | |
| Species | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | Mean |
|---|---|---|---|---|---|---|---|---|
| brown hare (ind./km2) | 3 | 4 | 1 | 2 | 2 | 1 | 2 | 2.1 |
| European rabbit | no colonies recorded | |||||||
| grey partridge (n males/km2) | 0 | 0 | 0 | 0.5 | 0 | 0.5 | 0 | 0.1 |
| pheasant (n males/km2) | 2 | 1 | 3 | 2 | 3 | 3 | 2 | 2.3 |
References
- Cramp, S.; Simmons, K.E.L. (Eds.) Handbook of the Birds of Europe, the Middle East and North Africa: The Birds of the Western Palearctic; Hawks to Bustards; Oxford University Press: Oxford, UK, 1980; Volume 2. [Google Scholar]
- Palmer, R.S. Handbook of North American Birds; Yale University Press: New Haven, CT, USA, 1988; Volume 4. [Google Scholar]
- Rutz, C.; Bijlsma, R.G.; Marquiss, M.; Kenward, R.E. Population limitation in the Northern Goshawk in Europe: A review with case studies. Stud. Avian Biol. 2006, 31, 158–197. [Google Scholar]
- Pielowski, Z. Die Greifvögel; Verlag J. Neumann-Neudamm GmbH and Co.: Morshen/Heina, Germany, 1993. [Google Scholar]
- Petty, S. History of northern goshawk Accipiter gentilis in Britain. In The Introduction and Naturalization of Birds; Holmes, J.S., Simons, J.R., Eds.; Joint Nature Conservation Committee: London, UK, 1996; pp. 95–102. [Google Scholar]
- Widén, P. How, and why, is the Goshawk (Accipiter gentilis) affected by modern forest management in Fennoscandia. J. Raptor Res. 1997, 31, 107–113. [Google Scholar]
- Newton, I. Population Limitation in Birds; Academic Press: London, UK, 1998. [Google Scholar]
- Kovács, A.; Burfield, I. Diurnal forest raptors in Europe: Population estimates, trends, threats and conservation. In Ecology and Conservation of European Forest-Dwelling Raptors; Zuberogoitia, I., Martínez, J.E., Eds.; Departamento de Agricultura de la Diputación Foral de Bizkaia: Bilbao, Spain, 2012; pp. 28–39. [Google Scholar]
- Petty, S.J. Understanding the impact of large-scale reforestation on raptors in the uplands of Britain: An overview of some recent research and implications for forest management. In Ecology and Conservation of European Forest-Dwelling Raptors; Zuberogoitia, I., Martínez, J.E., Eds.; Departamento de Agricultura de la Diputación Foral de Bizkaia: Bilbao, Spain, 2012; pp. 304–313. [Google Scholar]
- Kjellén, N.; Roos, G. Population trends in Swedish raptors demonstrated by migration counts at Falsterbo, Sweden 1942–97. Bird Study 2000, 47, 95–211. [Google Scholar]
- Saurola, P.; Björklund, H. Forest raptors: Conservation, ecology, behavior and management implications. Losers and winners among Finnish forest dwelling birds of prey. In Ecology and Conservation of European Forest-Dwelling Raptors; Zuberogoitia, I., Martínez, J.E., Eds.; Departamento de Agricultura de la Diputación Foral de Bizkaia: Bilbao, Spain, 2012; pp. 56–69. [Google Scholar]
- Chylarecki, P.; Chodkiewicz, T.; Neubauer, G.; Sikora, A.; Meissner, W.; Woźniak, B.; Wylegała, P.; Ławicki, Ł.; Marchowski, D.; Betleja, J.; Bzoma, Sz.; Cenian, Z.; et al. Trendy Liczebności ptaków w Polsce; GIOŚ: Warsaw, Poland, 2018. (In Polish)
- Kennedy, P.L. The northern goshawk (Accipiter gentilis atricapillus): Is there evidence of a population decline? J. Rapt. Res. 1997, 31, 95–106. [Google Scholar]
- Boyce, D.A.; Reynolds, R.T.; Graham, R.T. Goshawk status and management: What do we know, what have we done, where are we going? Stud. Avian. Biol. 2006, 31, 312–325. [Google Scholar]
- Saga, Ø.; Selås, V. Nest reuse by Goshawks after timber harvesting: Importance of distance to logging remaining mature forest area and tree species composition. For. Ecol. Manag. 2012, 270, 66–70. [Google Scholar] [CrossRef]
- Rodriguez, S.A.; Kennedy, P.L.; Parker, T.H. Timber harvest and tree size near nest explains variation in nest site occupancy but not productivity in northern goshawks (Accipiter gentilis). For. Ecol. Manag. 2016, 374, 220–229. [Google Scholar] [CrossRef]
- Adamski, A.; Olech, B.; Lontkowski, J. Goshawk Accipiter gentilis. In The Atlas of Breeding Birds in Poland 1985–2004; Sikora, A., Rodhe, Z., Gromadzki, M., Neubauer, G., Chylarecki, P., Eds.; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2007; pp. 145–147. [Google Scholar]
- Król, W. Breeding density of diurnal raptors in the neighbourhood of Susz (Iława Lakeland, Poland) in the years 1977–79. Acta Ornithol. 1985, 21, 95–114. [Google Scholar]
- Goszczyński, J. Density and productivity of common buzzard Buteo buteo and Goshawk Accipiter gentilis populations in Rogów, central Poland. Acta Ornithol. 1997, 32, 149–155. [Google Scholar]
- Kwieciński, Z.; Mizera, T. Numbers and breeding effects of bird of prey Falconiformes in the Kotlina Śremska region in 2001–2002. Not. Orn. 2006, 47, 230–240. [Google Scholar]
- Rebollo, S.; Martínez-Hesterkamp, S.; García-Salgado, G.; Pérez-Comacho, L.; Fernández-Pereira, J.M.; Jenness, J. Spatial relationships and mechanisms of coexistence between dominant and subdominant top predators. J. Avian Biol. 2017, 48, 1226–1237. [Google Scholar] [CrossRef]
- Goszczyński, J.; Piłatowski, T. Diet of common buzzards (Buteo buteo) and Goshawks (Accipiter gentilis) in the nesting period. Ekol. Pol. 1986, 34, 655–667. [Google Scholar]
- Goszczyński, J. The food habits of Buzzards and Goshawks during the nesting period. In Proceedings of the XXth Congress of the International Union of Game Biologists, Gödöllö, Hungary, 21–26 August 1991; pp. 387–390. [Google Scholar]
- Gryz, J.; Krauze, D.; Goszczyński, J. Abundance of bird of prey Falconiformes and raven Corvus corax in the vicinity of Rogów, central Poland. Not. Orn. 2006, 47, 43–57. [Google Scholar]
- Gryz, J.; Krauze-Gryz, D. Density dynamics, diet composition, and productivity of sparrowhawk population in central Poland. For. Res. Papers 2018, 79, 245–251. [Google Scholar]
- Kenward, R.E.; Marcstrȯm, V.; Karlbom, M. Demographic estimates from radio-tagging: Models of age-specific survival and breeding in the goshawk. J. Anim. Ecol. 1999, 68, 1020–1033. [Google Scholar] [CrossRef]
- Ivanovsky, V.V. Current status and breeding ecology of the goshawk Accipiter gentilis in northern Belarus. In Holarctic Birds of Prey. Proceedings of the International Conference on Holarctic Birds of Prey and Owls, 1995 at Badajoz, Extremadura, Spain; Chancellor, R.D., Meyburg, B.U., Ferrero, J.J., Eds.; ADENEX: Mérida, Spain; WWGBP: Berlin, Germany, 1998; pp. 111–115. [Google Scholar]
- Erdman, T.C.; Brinker, D.F.; Jacobs, J.P.; Wilde, J.; Meyer, T.O. Productivity, population trends, and status of Northern Goshawk, Accipiter gentilis atricapillus, in north-eastern Wisconsin. Can. Field Nat. 1998, 112, 17–27. [Google Scholar]
- Goszczyński, J. The breeding performance of the Common Buzzard Buteo buteo and Goshawk Accipiter gentilis in Central Poland. Acta Ornithol. 2001, 36, 105–110. [Google Scholar] [CrossRef]
- Juszko, S. Influence of Predation on Mortality of Brown Hare in Central Poland. Ph.D. Thesis, Warsaw University of Life Sciences, Warsaw, Poland, 2005. [Google Scholar]
- Krauze, D.; Gryz, J.; Goszczyński, J. Food composition of the goshawk Accipiter gentilis L. 1758 during the nesting season in the Rogów Forest (central Poland). Folia For. Pol. Ser A-For. 2005, 47, 45–53. [Google Scholar]
- Gryz, J.; Krauze-Gryz, D. The influence of raptors (Falconiformes) and raven (Corvus corax) on populations of game animals. Ann. WULS-SGGW. For. Wood Tech. 2014, 86, 114–124. [Google Scholar]
- Goszczyński, J.; Gryz, J.; Krauze, D. Fluctuations of a Common Buzzard Buteo buteo population in Central Poland. Acta Ornithol. 2005, 40, 75–78. [Google Scholar] [CrossRef]
- Wasilewski, G. (The Rogów Forest District). Personal communication, 2018.
- Pucek, Z. (Ed.) Keys to Vertebrates of Poland-Mammals; Polish Scientific Publishers: Warsaw, Poland, 1981. [Google Scholar]
- Moreno, E. Clave osteologica para la identificatión de los Passeriformes Ibéricos. 1. Aegithalidae, Remizidae, Fringilliade, Alaudidae. Ardeola 1985, 32, 295–377. [Google Scholar]
- Moreno, E. Clave osteologica para la identificatión de los Passeriformes Ibéricos. 2. Hirundinidae, Prunellidae, Sittidae, Certhiidae, Troglodytidae, Cinclidae, Laniidae, Oriolidae, Corvidae, Sturnidae, Motacillidae. Ardeola 1986, 33, 69–129. [Google Scholar]
- März, R. Gewöll–und Rupfungskunde; Akademie-Verlang: Berlin, Germany, 1987. [Google Scholar]
- Ujhelyi, P. Identification of the Fringillidae of Europe on the basis of craniometric characteristics. Aquila 1992, 99, 99–110. [Google Scholar]
- Teerink, B.J. Hair of West-European Mammals; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Szczepski, B.; Kozłowski, P. Pomocnicze Tabele Ornitologiczne; PWN: Warsaw, Poland, 1953. [Google Scholar]
- Jędrzejewska, B.; Jędrzejewski, W. Predation in Vertebrate Communities, The Białowieża Primeval Forest as a Case Study; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Aulak, W.; Rowiński, P. Tabele Biologiczne Kręgowców; Wydawnictwo SGGW: Warsaw, Poland, 2010. [Google Scholar]
- Gryz, J.; Chojnacka-Ożga, L.; Krauze-Gryz, D. A tawny owl (Strix aluco) population remains stable despite changing environmental conditions—A case study from central Poland. Pol. J. Ecol. 2019, in press. [Google Scholar]
- Penteriani, V.; Faivre, B. Effects of harvesting timber stands on goshawk nesting in two European areas. Biol. Conserv. 2001, 101, 211–216. [Google Scholar] [CrossRef]
- Mañosa, S. Goshawk diet in a Mediterranean area of northern Spain. J. Rapt. Res. 1994, 28, 84–92. [Google Scholar]
- Lindén, H.; Wikman, M. Goshawk predation on tetraonids: Availability of prey and diet of the predator. J. Anim. Ecol. 1983, 52, 953–968. [Google Scholar] [CrossRef]
- Tornberg, R. Prey selection of the Goshawk Accipiter gentilis during the breeding season: The role of prey profitability and vulnerability. Orn. Fenn. 1997, 74, 15–28. [Google Scholar]
- Tomešek, M.; Čermák, P. Distribution and biology of goshawk (Accipiter gentilis) in the Chřiby Upland, the Czech Republic. Acta Univ. Agric. Silvic Mendel Brun 2009, 57, 1–11. [Google Scholar] [CrossRef]
- Tomešek, M.; Kratěna, L.; Čermák, P.; Tomešek, K. Utilisation of an video camera in study of the goshawk (Accipiter gentilis) diet. Acta Univ. Agric Silvic Mendel Brun 2011, 59, 227–234. [Google Scholar] [CrossRef]
- Nielsen, J.T.; Drachmann, J. Prey selection of goshawks Accipiter gentilis during the breeding season in Vendsyssel, Denmark. Dansk Orn. Foren. Tidsskr. 1999, 93, 85–90. [Google Scholar]
- Dudziński, W. Grey partridge (Perdix perdix)—Predator relationship in cropland and forest habitat of central Poland. Gibier Faune Sauvage 1992, 9, 455–466. [Google Scholar]
- Wasilewski, M. Population dynamics of pheasants near Rogów, central Poland. Ekol. Pol. 1986, 34, 669–680. [Google Scholar]
- Jacewicz, J. Population of Magpie (Pica pica) in North-East Part of 59 SGGW-AR Hunting District. Master’s Thesis, Warsaw University of Life Sciences, Warsaw, Poland, 1984. [Google Scholar]
- Gryz, J. Ecology of Northern Goshawk Accipiter Gentilis Population in the Experimental Forest Station in Rogów. Master’s Thesis, Warsaw University of Life Sciences, Warsaw, Poland, 2003. [Google Scholar]
- Gryz, J.; Krauze, D.; Juszko, S. Long-Term Decrease of Brown Hare Population in Central Poland; Book of Abstracts; International Union of Game Biologists XXVIII Congress: Uppsala, Sweden, 2007; p. 209. [Google Scholar]
- Gryz, J.; Krauze-Gryz, D.; Lesiński, G. Mammals in the vicinity of Rogów (central Poland). Fragm. Faun. 2011, 54, 183–197. [Google Scholar] [CrossRef]
- Olech, B. Diet of the goshawk Accipiter gentilis in Kampinoski National Park (central Poland) in 1982–1993. Acta Ornithol. 1997, 32, 191–200. [Google Scholar]
- Zawadzka, D.; Zawadzki, J. The goshawk Accipiter gentilis in Wigry National Park (NE Poland)—Numbers, breeding results, diet composition and prey selection. Acta Ornithol. 1998, 33, 181–190. [Google Scholar]
- Rebollo, S.; García-Salgado, G.; Pérez-Camacho, L.; Martínez-Hesterkamp, S.; Navarro, A.; Fernández-Pereira, J.M. Prey preferences and recent changes in diet of breeding population of the Northern Goshawk Accipiter gentilis in Southwestern Europe. Bird Study 2017, 64, 464–475. [Google Scholar] [CrossRef]
- Penteriani, V.; Rutz, Ch.; Kenvard, R. Hunting behaviour and breeding performance of northern goshawks Accipiter gentilis, in relations to resources availability, sex, age and morphology. Naturwissenschaften 2013, 100, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Tornberg, R.; Rytkönen, S.; Välimäki, P.; Valkama, J.; Helle, P. Northern goshawk (Accipiter gentilis) may improve black grouse breeding success. J. Ornithol. 2015, 157, 363–370. [Google Scholar] [CrossRef]
- Olech, B. Protection of birds of prey in the Kampinos National Park—Present state and indications. Ochr. Przyr. 1991, 49, 65–92. [Google Scholar]
- Goszczyński, J.; Misiorowska, M.; Juszko, S. Changes in the density and spatial distribution of red fox dens and cub numbers in central Poland following rabies vaccination. Acta Theriol. 2008, 53, 121–127. [Google Scholar] [CrossRef]
- Gryz, J.; Krauze-Gryz, D. Dynamics of red fox population in Białowieża Primeval Forest in the years 1981–2016. Sylwan 2017, 161, 328–333. [Google Scholar]
- Panek, M.; Kamieniarz, R.; Bresiński, W. The effect of experimental removal of red foxes Vulpes vulpes on spring density of brown hares Lepus europaeus in western Poland. Acta Theriol. 2006, 51, 187–193. [Google Scholar] [CrossRef]
- Selås, V. Does food competition from red fox (Vulpes vulpes) influence the breeding density of goshawk (Accipiter gentilis)? Evidence from a natural experiment. J. Zool. 1998, 246, 325–553. [Google Scholar] [CrossRef]
- Gryz, J.; Krauze-Gryz, D. Rare species of birds nesting in the area of Rogów Forest District in the years 1949–2015. For. Res. Papers 2016, 77, 134–140. [Google Scholar] [CrossRef]
- Greenwald, D.N.; Crocker-Bedford, D.C.; Broberg, L.; Suckling, K.F.; Tibbitts, T. Review of Northern Goshawk Habitat Selection in the Home Range and Implications for Forest Management in the Western United States. Wildl. Soc. Bull. 2005, 33, 120–129. [Google Scholar] [CrossRef]
- Reich, R.M.; Joy, S.M.; Reynolds, R.T. Predicting the location of northern goshawk nests: Modelling the spatial dependency between nest location and forest structure. Ecol. Model. 2004, 176, 109–133. [Google Scholar] [CrossRef]
- García-Salgado, G.; Rebollo, S.; Pérez-Camacho, L.; Martinez-Hesterkamp, S.; de la Montaña, E.; Domingo-Muñoz, R.; Madrigal-González, J.; Fernández-Pereira, J.M. Breeding habitat preferences and reproductive success of Northern Goshawk (Accipiter gentilis) in exotic Eucaliptus plantation in southwestern Europe. For. Ecol. Manag. 2018, 409, 817–825. [Google Scholar] [CrossRef]
- Natsukawa, H.; Ichinose, T.; Higuchi, H. Factors affecting breeding–site selection of Northern Goshawks at two spatial scale in urbanized area. J. Rapt. Res. 2017, 51, 417–427. [Google Scholar] [CrossRef]
- Zielony, R. (Ed.) Natural Conditions of Experimental Forest of Agricultural University of Warsaw in Rogów; Wydawnictwo SGGW: Warszawa, Poland, 1993. [Google Scholar]
- Pugacewicz, E. Breeding Birds of the Białoweża Forest; Północnopodlaskie Towarzystwo Ochrony Ptaków: Białystok, Poland, 1996. [Google Scholar]


| Stand Characteristics | Year of Assessment | ||
|---|---|---|---|
| 1989 | 1999 | 2009 | |
| % of Area | |||
| stands 60–100 years old | 36.4 | 34.2 | 42.1 |
| stands over 100 years old | 1.3 | 6.0 | 7.9 |
| Scots pine Pinus sylvestris | 76.9 | 69.9 | 49.9 |
| larch Larix spp. | 4.1 | 6.1 | 6.7 |
| oaks Quercus spp. | 10.7 | 13.8 | 19.4 |
| common beech Fagus sylvatica | 1.0 | 3.0 | 9.7 |
| Forest Complex | Study Periods | ||
|---|---|---|---|
| 1982–1992 | 2001–2003 | 2011–2018 (SD) | |
| 1 | 6.7 | 2.9 | 3.6 (0.7) |
| 2 | 3.8 | 2.7 | 1.6 (0.9) |
| 3 | 1.8 | 2.0 | 0.4 (0.5) |
| 4 | 1.1 | 1.0 | 0.9 (0.3) |
| 5 | 1.5 | 2.0 | 1.0 (0.0) |
| 6 | 1.1 | 0 | 0 (0.0) |
| 7 | 0.8 | 0 | 0.5 (0.5) |
| whole area | 17.1 | 11.0 | 8.0 |
| Breeding Parameter | 1982–1992 | 2001–2002 (SD) | 2011–2018 (SD) |
|---|---|---|---|
| number of fledglings per pair | 2.1 | 1.8 (1.19) | 1.6 (1.07) |
| number of fledglings per successful pair | 2.8 | 2.4 (0.53) | 2.1 (0.69) |
| breeding success (%) | 75 | 75 | 76 (7.2) |
| Nesting Trees/Nests Characteristics | Study Period | ||
|---|---|---|---|
| 1982–1992 | 2001–2003 | 2011–2018 | |
| Tree species (%) | |||
| Pinus sylvestris | 84.7 | 60.0 | 45.9 |
| Larix decidua | 5.5 | 13.0 | 32.4 |
| Abies alba | 0.0 | 13.0 | 2.7 |
| Picea abies | 1.2 | 7.0 | 2.7 |
| Quercus spp. | 3.1 | 0.0 | 0.0 |
| Alnus glutinosa | 2.4 | 7.0 | 16.2 |
| Betula verrucosa | 3.1 | 0.0 | 0.0 |
| Tree measurements | |||
| diameter (cm) | 42 | 44 | 48 |
| age (years) | 75 | 77 | 84 |
| Nest placement | |||
| height (m) | 18 | 20 | 21 |
| number of nesting trees | 163 | 15 | 37 |
| Prey Category | 1982–1990 (%) | 2001–2002 (%) | 2011–2018 (%) | |||
|---|---|---|---|---|---|---|
| Prey Items | Biomass | Prey Items | Biomass | Prey Items | Biomass | |
| Domestic pigeon | 38.0 | nd | 32.3 | 57.4 | 22.8 | 45.9 |
| Poultry | 2.9 | 10.1 | 2.2 | 5 | 1.3 | 3.3 |
| Wood pigeon | 2.9 | nd | 6.8 | 14.3 | 9.0 | 21.8 |
| European jay | 2.9 | 3.0 | 8.1 | 6.3 | 13.1 | 11.6 |
| Small game | 2.7 | 4.0 | 2.6 | 3.4 | 0.3 | 0.3 |
| total N prey items/biomass (g) | 1 513 | nd | 310 | 69 629 | 1 065 | 211 664 |
| Prey | 1982–1992 | 2001–2003 | 2011–2018 |
|---|---|---|---|
| brown hare | 30 ind./km2 [56] | 8–13 ind./km2 [56] | 2.1 ind./km2 |
| European rabbit | at least five big colonies [57] | last rabbits in 2007 [57] | no colonies recorded |
| grey partridge | 21–33 ind./km2 (autumn) [52] | nd | 0.1 males/km2 (spring) |
| pheasant | 5.7–11.0 ind./km2 [53] | nd | 2.3 males/km2 |
| magpie | 1.6 pairs/km2 [54] | nd | 1.6 pairs/km2 |
| rook | nd | nd | no nests recorded |
| hooded crow | nd | nd | 0.15 pairs/km2 |
| jackdaw | nd | nd | 0.25 pairs/km2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gryz, J.; Krauze-Gryz, D. Pigeon and Poultry Breeders, Friends or Enemies of the Northern Goshawk Accipiter gentilis? A Long-Term Study of a Population in Central Poland. Animals 2019, 9, 141. https://doi.org/10.3390/ani9040141
Gryz J, Krauze-Gryz D. Pigeon and Poultry Breeders, Friends or Enemies of the Northern Goshawk Accipiter gentilis? A Long-Term Study of a Population in Central Poland. Animals. 2019; 9(4):141. https://doi.org/10.3390/ani9040141
Chicago/Turabian StyleGryz, Jakub, and Dagny Krauze-Gryz. 2019. "Pigeon and Poultry Breeders, Friends or Enemies of the Northern Goshawk Accipiter gentilis? A Long-Term Study of a Population in Central Poland" Animals 9, no. 4: 141. https://doi.org/10.3390/ani9040141
APA StyleGryz, J., & Krauze-Gryz, D. (2019). Pigeon and Poultry Breeders, Friends or Enemies of the Northern Goshawk Accipiter gentilis? A Long-Term Study of a Population in Central Poland. Animals, 9(4), 141. https://doi.org/10.3390/ani9040141