Habitat Selection of Wintering Birds in Farm Ponds in Taoyuan, Taiwan



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
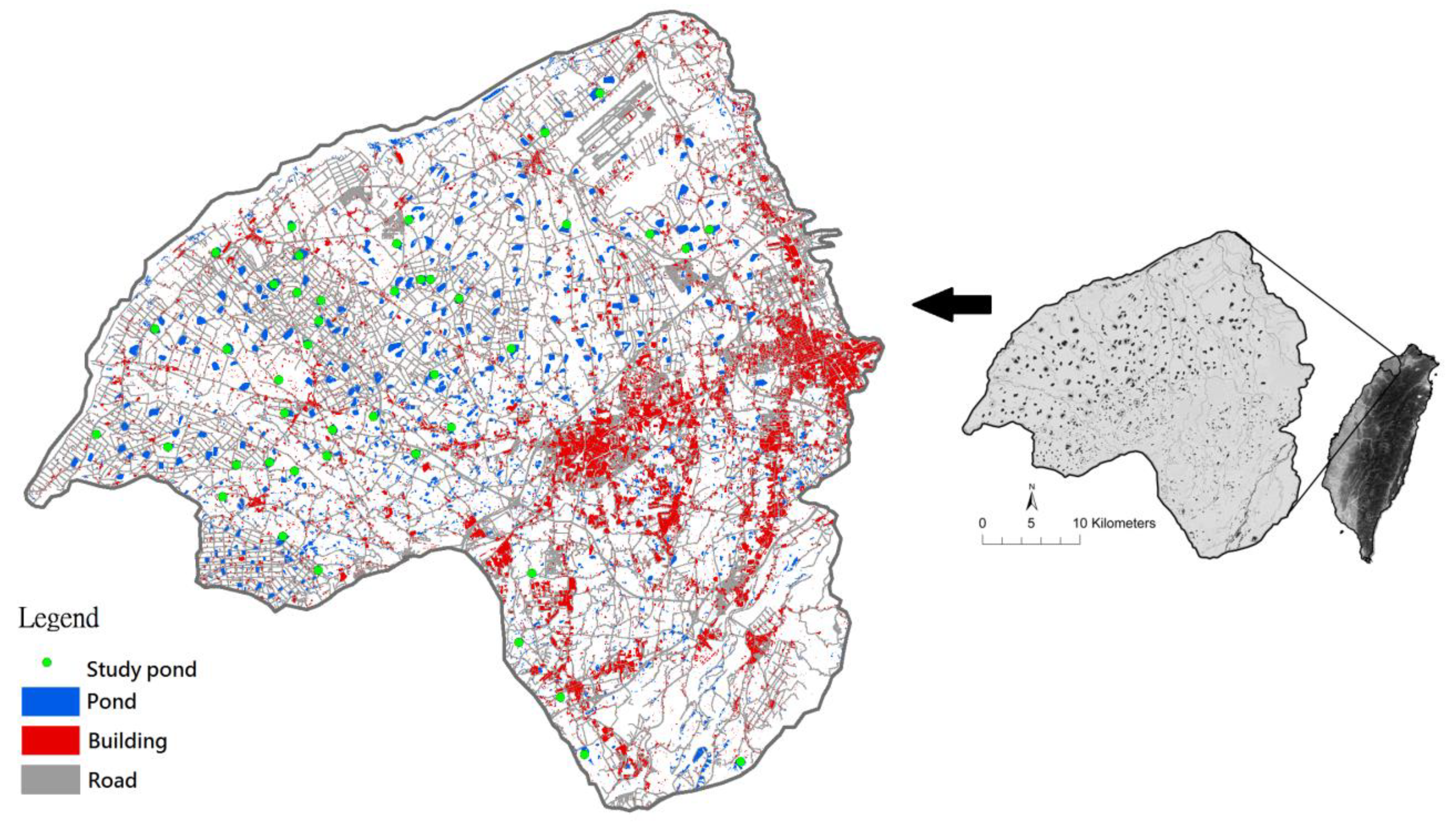
2.1. Study Site
2.2. Sampling
3. Results
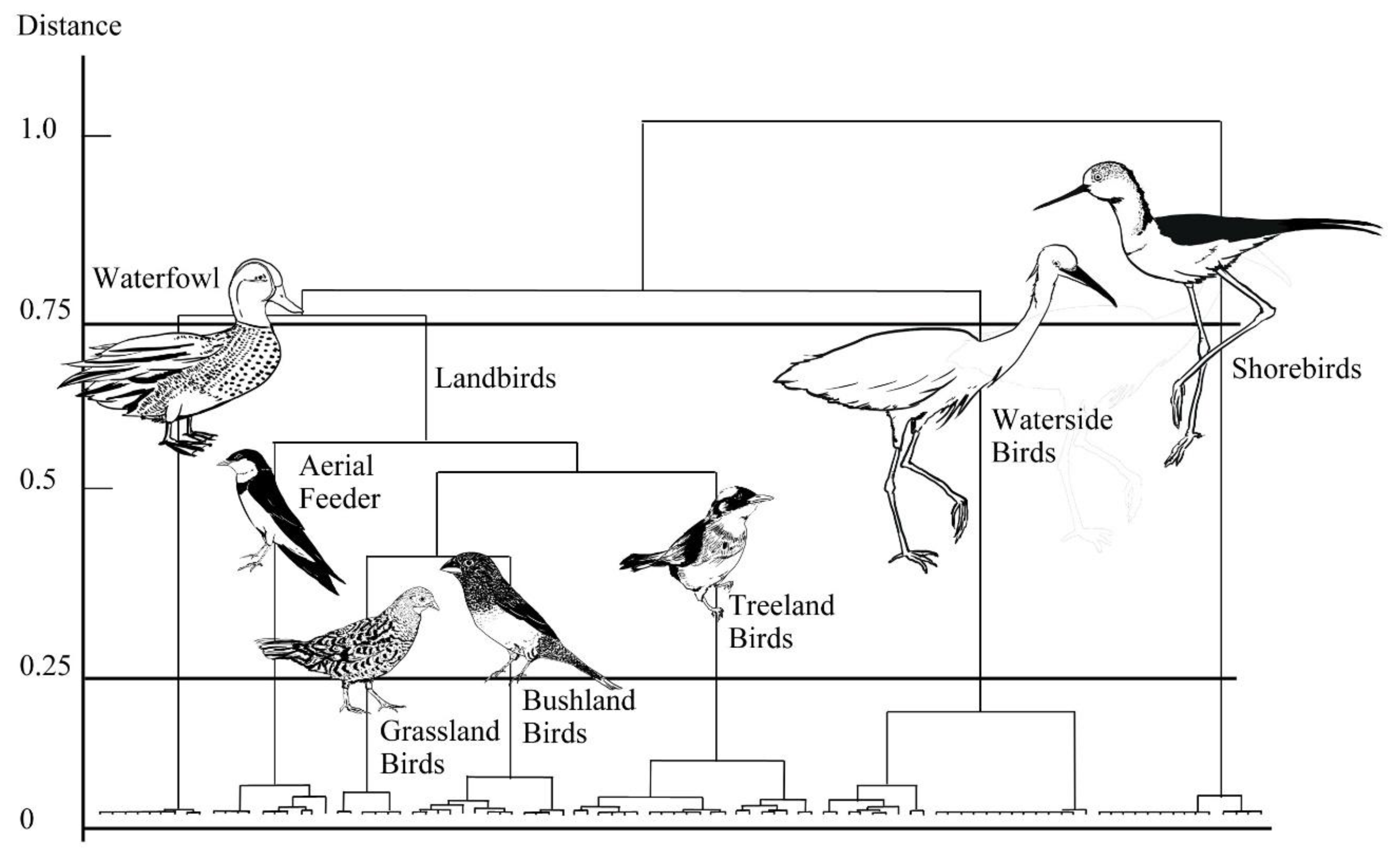
3.1. Classification of Guilds
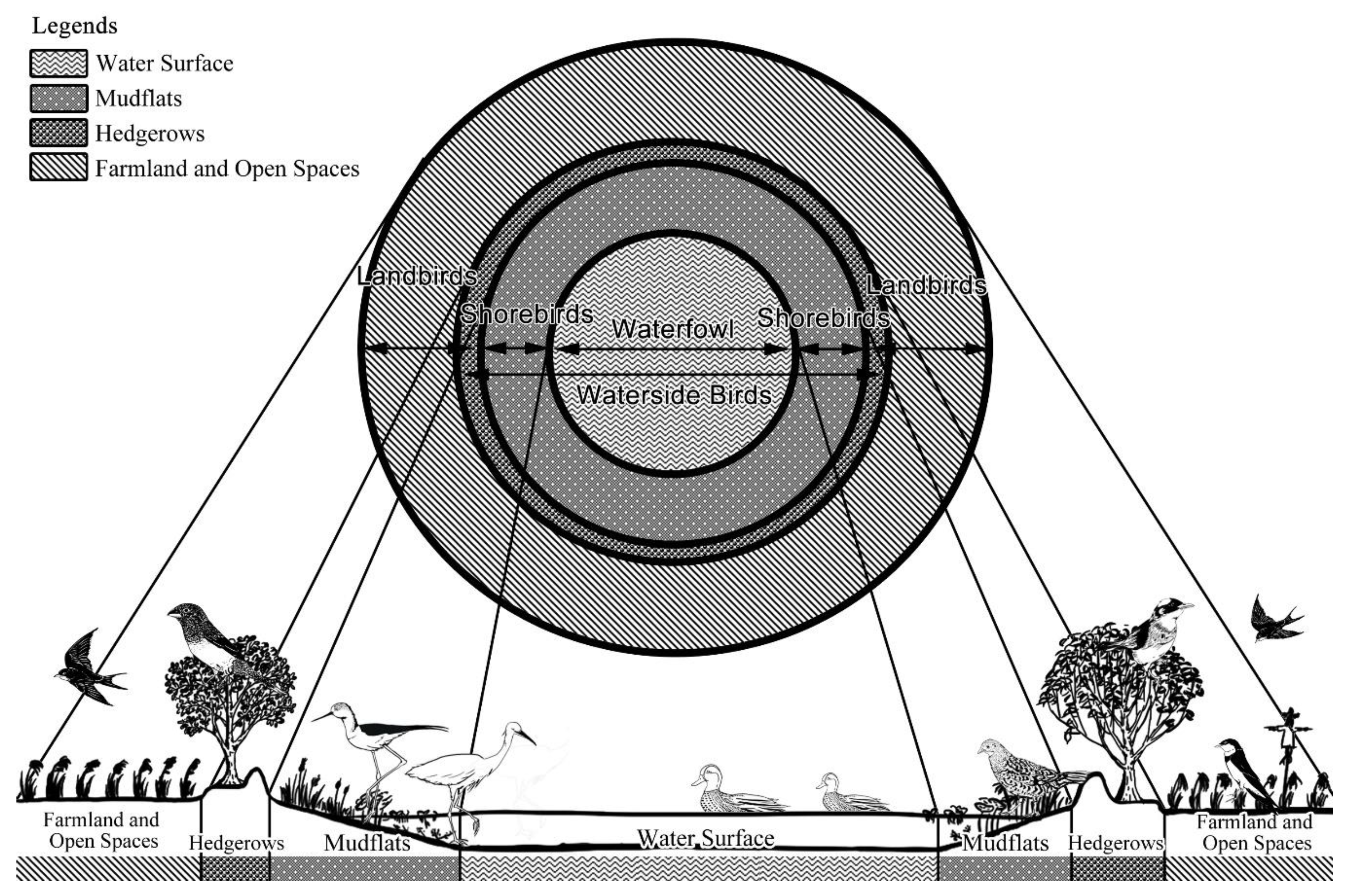
3.2. Habitat Differences with Classification of Guilds
3.3. Richness and Abundance of Guilds Associated with Environmental Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| ID | Code | Common Name | Scientific Name | Air | Waters | Mudflats | Trails | Grasslands | Bushlands | Woodlands | Avian Guilds |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1402 | Northern pintail | Anas acuta | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | Waterfowl ** |
| 2 | 1403 | Northern shoveller | Anas clypeata | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | Waterfowl ** |
| 3 | 1404 | Common teal | Anas crecca | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | Waterfowl ** |
| 4 | 1408 | Eurasian wigeon | Anas penelope | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | Waterfowl ** |
| 5 | 1410 | Spot-billed duck | Anas poecilorhyncha | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | Waterfowl ** |
| 6 | 1419 | Common pochard | Aythya ferina | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | Waterfowl ** |
| 7 | 1523 | Black-eared kite | Milvus migrans lineatus | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | Aerial Feeder ** |
| 8 | 2102 | Common coot | Fulica atra | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | Shorebird ** |
| 9 | 2106 | Ruddy-breasted crake | Porzana fusca | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | Shorebird ** |
| 10 | 2608 | Grey-headed lapwing | Vanellus cinereus | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | Shorebird ** |
| 11 | 2609 | Pacific golden plover | Pluvialis fulva | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | Shorebird ** |
| 12 | 2738 | Common redshank | Tringa totanus | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | Shorebird ** |
| 13 | 2801 | Black-winged stilt | Himantopus himantopus | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | Shorebird ** |
| 14 | 2802 | Pied avocet | Recurvirostra avosetta | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | Shorebird ** |
| 15 | 4002 | Fork-tailed swift | Apus pacificus | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | Aerial Feeder ** |
| 16 | 4903 | Striated swallow | Hirundo striolata | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | Aerial Feeder ** |
| 17 | 4905 | Plain sand martin | Riparia paludicola | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | Aerial Feeder ** |
| 18 | 6701 | Red-throated pipit | Anthus cervinus | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | Shorebird ** |
| 19 | 2735 | Common greenshank | Tringa nebularia | 0.00 | 0.00 | 0.97 | 0.03 | 0.00 | 0.00 | 0.00 | Shorebird |
| 20 | 1421 | Greater scaup | Aythya marila | 0.00 | 0.95 | 0.00 | 0.00 | 0.00 | 0.00 | 0.05 | Waterfowl |
| 21 | 205 | Little grebe | Tachybaptus ruficollis | 0.00 | 0.94 | 0.00 | 0.06 | 0.00 | 0.00 | 0.00 | Waterfowl |
| 22 | 2104 | Common moorhen | Gallinula chloropus | 0.00 | 0.00 | 0.89 | 0.08 | 0.03 | 0.00 | 0.01 | Shorebird |
| 23 | 1409 | Mallard | Anas platyrhynchos | 0.00 | 0.88 | 0.00 | 0.12 | 0.00 | 0.00 | 0.00 | Waterfowl |
| 24 | 4001 | House swift | Apus nipalensis | 0.81 | 0.00 | 0.00 | 0.19 | 0.00 | 0.00 | 0.00 | Aerial Feeder |
| 25 | 2731 | Wood sandpiper | Tringa glareola | 0.00 | 0.00 | 0.80 | 0.20 | 0.00 | 0.00 | 0.00 | Shorebird |
| 26 | 3207 | Common black-headed gull | Larus ridibundus | 0.75 | 0.00 | 0.00 | 0.25 | 0.00 | 0.00 | 0.00 | Aerial Feeder |
| 27 | 2601 | Kentish plover | Charadrius alexandrinus | 0.00 | 0.00 | 0.73 | 0.27 | 0.00 | 0.00 | 0.00 | Shorebird |
| 28 | 4904 | Pacific swallow | Hirundo tahitica | 0.73 | 0.00 | 0.00 | 0.19 | 0.01 | 0.00 | 0.07 | Aerial Feeder |
| 29 | 2611 | Northern lapwing | Vanellus vanellus | 0.00 | 0.00 | 0.72 | 0.28 | 0.00 | 0.00 | 0.00 | Shorebird |
| 30 | 2603 | Little ringed plover | Charadrius dubius | 0.00 | 0.00 | 0.77 | 0.17 | 0.06 | 0.00 | 0.00 | Shorebird * |
| 31 | 1108 | Great egret | Casmerodius alba | 0.00 | 0.59 | 0.00 | 0.34 | 0.00 | 0.00 | 0.07 | Waterside bird * |
| 32 | 4101 | Common kingfisher | Alcedo atthis | 0.55 | 0.00 | 0.00 | 0.25 | 0.02 | 0.11 | 0.08 | Aerial Feeder |
| 33 | 4902 | Barn swallow | Hirundo rustica | 0.53 | 0.00 | 0.00 | 0.47 | 0.00 | 0.00 | 0.00 | Aerial Feeder |
| 34 | 1706 | Common kestrel | Falco tinnunculus | 0.50 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.50 | Aerial Feeder |
| 35 | 1111 | Intermediate egret | Mesophoyx intermedia | 0.00 | 0.47 | 0.00 | 0.45 | 0.00 | 0.00 | 0.08 | Waterside bird |
| 36 | 1110 | Little egret | Egretta garzetta | 0.00 | 0.54 | 0.00 | 0.28 | 0.01 | 0.04 | 0.13 | Waterside bird * |
| 37 | 2733 | Common sandpiper | Tringa hypoleucos | 0.00 | 0.00 | 0.43 | 0.51 | 0.04 | 0.03 | 0.00 | Waterside bird |
| 38 | 1101 | Grey heron | Ardea cinerea | 0.00 | 0.34 | 0.00 | 0.46 | 0.02 | 0.01 | 0.17 | Waterside bird * |
| 39 | 901 | Common cormorant | Phalacrocorax carbo | 0.00 | 0.32 | 0.00 | 0.64 | 0.00 | 0.00 | 0.04 | Waterside bird |
| 40 | 2703 | Dunlin | Calidris alpina | 0.00 | 0.00 | 0.28 | 0.72 | 0.00 | 0.00 | 0.00 | Waterside bird |
| 41 | 1121 | Black-crowned night heron | Nycticorax nycticorax | 0.00 | 0.23 | 0.00 | 0.59 | 0.01 | 0.07 | 0.10 | Waterside bird * |
| 42 | 6707 | White wagtail | Motacilla alba | 0.00 | 0.22 | 0.00 | 0.27 | 0.51 | 0.00 | 0.00 | Grassland bird |
| 43 | 5410 | Black-billed magpie | Pica pica | 0.20 | 0.00 | 0.00 | 0.07 | 0.00 | 0.10 | 0.63 | Woodland bird |
| 44 | 1601 | Osprey | Pandion haliaetus | 0.17 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.83 | Woodland bird |
| 45 | 5914 | Rufous-capped babbler | Stachyris ruficeps | 0.00 | 0.14 | 0.00 | 0.14 | 0.00 | 0.00 | 0.71 | Woodland bird |
| 46 | 3509 | Red-collared dove | Streptopelia tranquebarica | 0.00 | 0.13 | 0.00 | 0.19 | 0.01 | 0.02 | 0.66 | Woodland bird * |
| 47 | 6710 | Yellow wagtail | Motacilla flava | 0.00 | 0.11 | 0.00 | 0.22 | 0.64 | 0.02 | 0.01 | Grassland bird |
| 48 | 5403 | Large-billed crow | Corvus macrorhynchos | 0.00 | 0.11 | 0.00 | 0.28 | 0.00 | 0.00 | 0.60 | Woodland bird |
| 49 | 6708 | Grey wagtail | Motacilla cinerea | 0.00 | 0.11 | 0.00 | 0.63 | 0.22 | 0.03 | 0.01 | Waterside bird |
| 50 | 5913 | Steak-breasted; Scimitar babbler | Pomatorhinus ruficollis | 0.00 | 0.05 | 0.00 | 0.16 | 0.00 | 0.68 | 0.11 | Bushland bird |
| 51 | 5103 | Black drongo | Dicrurus macrocercus | 0.00 | 0.05 | 0.00 | 0.29 | 0.19 | 0.14 | 0.34 | Woodland bird |
| 52 | 6313 | Daurian redstart | Phoenicurus auroreus | 0.00 | 0.04 | 0.00 | 0.21 | 0.38 | 0.13 | 0.25 | Woodland bird |
| 53 | 6422 | Plain prinia | Prinia inornata | 0.00 | 0.04 | 0.00 | 0.36 | 0.08 | 0.47 | 0.05 | Bushland bird |
| 54 | 7201 | Japanese white-eye | Zosterops japonica | 0.00 | 0.03 | 0.00 | 0.03 | 0.01 | 0.10 | 0.84 | Woodland bird * |
| 55 | 3507 | Spotted dove | Streptopelia chinensis | 0.00 | 0.02 | 0.00 | 0.21 | 0.08 | 0.00 | 0.68 | Woodland bird |
| 56 | 1105 | Cattle egret | Bubulcus ibis | 0.00 | 0.02 | 0.00 | 0.21 | 0.00 | 0.01 | 0.77 | Woodland bird |
| 57 | 7012 | White-vented myna | Acridotheres grandis | 0.00 | 0.02 | 0.00 | 0.53 | 0.00 | 0.00 | 0.45 | Woodland bird |
| 58 | 6003 | Light-vented bulbul | Pycnonotus sinensis | 0.00 | 0.01 | 0.00 | 0.14 | 0.02 | 0.14 | 0.69 | Woodland bird * |
| 59 | 7601 | Eurasian tree sparrow | Passer montanus | 0.00 | 0.01 | 0.00 | 0.23 | 0.04 | 0.21 | 0.51 | Woodland bird * |
| 60 | 1119 | Yellow bittern | Ixobrychus sinensis | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | Grassland bird |
| 61 | 1802 | Chinese bamboo partridge | Bambusicola thoracica | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | Grassland bird ** |
| 62 | 2101 | White-breasted waterhen | Amaurornis phoenicurus | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 63 | 2707 | Rufous-necked stint | Calidris ruficollis | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 64 | 2709 | Temminck’s stint | Calidris temminckii | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 65 | 2713 | Common snipe | Gallinago gallinago | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 66 | 2729 | Grey-tailed tattler | Tringa brevipes | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 67 | 3508 | Eastern turtle dove | Streptopelia orientalis | 0.00 | 0.00 | 0.00 | 0.05 | 0.00 | 0.03 | 0.91 | Woodland bird |
| 68 | 3512 | White-bellied green pigeon | Treron sieboldii | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | Woodland bird ** |
| 69 | 3601 | Lesser coucal | Centropus bengalensis | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 70 | 4501 | Black-browed barbet | Megalaima oorti | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | Woodland bird ** |
| 71 | 5407 | Grey treepie | Dendrocitta formosae | 0.00 | 0.00 | 0.00 | 0.00 | 0.43 | 0.43 | 0.14 | Bushland bird |
| 72 | 5502 | Vinous-throated parrotbill | Paradoxornis webbianus | 0.00 | 0.00 | 0.00 | 0.21 | 0.00 | 0.65 | 0.14 | Bushland bird |
| 73 | 6002 | Black bulbul | Hypsipetes leucocephalus | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 74 | 6307 | Siberian rubythroat | Luscinia calliope | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.67 | 0.00 | Bushland bird |
| 75 | 6317 | Orange-flanked bush-robin | Tarsiger cyanurus | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | Bushland bird ** |
| 76 | 6321 | Brown-headed thrush | Turdus chrysolaus | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 77 | 6325 | Dusky thrush | Turdus naumanni | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 78 | 6402 | Great reed warbler | Acrocephalus arundinaceus | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | Grassland bird ** |
| 79 | 6406 | Japanese bush warbler | Cettia diphone | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.67 | Woodland bird |
| 80 | 6407 | Brownish-flanked bush warbler | Cettia fortipes | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 81 | 6410 | Zitting cisticola | Cisticola juncidis | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | Bushland bird ** |
| 82 | 6421 | Yellow-bellied prinia | Prinia flaviventris | 0.00 | 0.00 | 0.00 | 0.22 | 0.12 | 0.56 | 0.10 | Bushland bird |
| 83 | 6703 | Olive-backed pipit | Anthus hodgsoni | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 84 | 6902 | Brown shrike | Lanius cristatus | 0.00 | 0.00 | 0.00 | 0.22 | 0.03 | 0.39 | 0.36 | Woodland bird |
| 85 | 6904 | Long-tailed shrike | Lanius schach | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.63 | 0.38 | Bushland bird |
| 86 | 7001 | Crested myna | Acridotheres cristatellus | 0.00 | 0.00 | 0.00 | 0.89 | 0.00 | 0.00 | 0.11 | Waterside bird |
| 87 | 7002 | Common myna | Acridotheres tristis | 0.00 | 0.00 | 0.00 | 0.31 | 0.00 | 0.23 | 0.46 | Woodland bird |
| 88 | 7005 | White-cheeked starling | Sturnus cineraceus | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | Grassland bird ** |
| 89 | 7007 | White-shouldered starling | Sturnus sinensis | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | Woodland bird ** |
| 90 | 7008 | Red-billed starling | Sturnus sericeus | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | Waterside bird ** |
| 91 | 7302 | Scaly-breasted munia | Lonchura punctulata | 0.00 | 0.00 | 0.00 | 0.15 | 0.26 | 0.48 | 0.11 | Bushland bird |
| 92 | 7303 | White-rumped munia | Lonchura striata | 0.00 | 0.00 | 0.00 | 0.00 | 0.14 | 0.86 | 0.00 | Bushland bird |
| 93 | 7511 | Black-faced bunting | Emberiza spodocephala | 0.00 | 0.00 | 0.00 | 0.20 | 0.20 | 0.54 | 0.07 | Bushland bird |
| 94 | 9902 | Rose-ringed parakeet | Psittacula krameri | 0.00 | 0.00 | 0.00 | 0.50 | 0.00 | 0.00 | 0.50 | Woodland bird |
References
- Gibbs, J.P. Wetland loss and biodiversity conservation. Conserv. Biol. 2000, 14, 314–317. [Google Scholar] [CrossRef]
- Gopal, B. Perspectives on wetland science, application and policy. Hydrobiologia 2003, 490, 1–10. [Google Scholar] [CrossRef]
- Takekawa, J.Y.; Lu, C.T.; Pratt, R.T. Avian communities in baylands and artificial salt evaporation ponds of the San Francisco Bay estuary. Hydrobiologia 2001, 466, 317–328. [Google Scholar] [CrossRef]
- French, D.; Picozzi, N. Functional groups’ of bird species, biodiversity and landscapes in Scotland. J. Biogeogr. 2002, 29, 231–259. [Google Scholar] [CrossRef]
- Froneman, A.; Mangnall, M.J.; Little, R.M.; Crowe, T.M. Waterbird assemblages and associated habitat characteristics of farm ponds in the Western Cape, South Africa. Biodivers. Conserv. 2001, 10, 251–270. [Google Scholar] [CrossRef]
- Eglington, S.M.; Noble, D.G.; Fuller, R.J. A meta-analysis of spatial relationships in species richness across taxa: Birds as indicators of wider biodiversity in temperate regions. J. Nat. Conserv. 2012, 20, 301–309. [Google Scholar] [CrossRef]
- Mistry, J.; Berardi, A.; Simpson, M. Birds as indicators of wetland status and change in the North Rupununi, Guyana. Biodivers. Conserv. 2008, 17, 2383–2409. [Google Scholar] [CrossRef]
- Duelli, P.; Obrist, M.K. Biodiversity indicators: The choice of values and measures. Agric. Ecosyst. Environ. 2003, 98, 87–98. [Google Scholar] [CrossRef]
- Fisher, I.J.; Pain, D.J.; Thomas, V.G. A review of lead poisoning from ammunition sources in terrestrial birds. Biol. Conserv. 2006, 131, 421–432. [Google Scholar] [CrossRef]
- Lambert, O.; Pouliquen, H.; Larhantec, M.; Thorin, C.; L’Hostis, M. Exposure of raptors and waterbirds to anticoagulant rodenticides (difenacoum, bromadiolone, coumatetralyl, coumafen, brodifacoum): Epidemiological survey in Loire Atlantique (France). Bull. Environ. Contam. Toxicol. 2007, 79, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Guitart, R.; Serratosa, J.; Thomas, V.G. Lead-poisoned wildfowl in Spain: A significant threat for human consumers. Int. J. Environ. Health Res. 2002, 12, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Holt, R.D. Population dynamics in two-patch environments: Some anomalous consequences of an optimal habitat distribution. Theor. Popul. Biol. 1985, 28, 181–208. [Google Scholar] [CrossRef]
- Tchouto, M.; De Wilde, J.; De Boer, W.; Van der Maesen, L.; Cleef, A. Bio-indicator species and Central African rain forest refuges in the Campo-Ma’an area, Cameroon. Syst. Biodivers. 2009, 7, 21–31. [Google Scholar] [CrossRef]
- Paracuellos, M. How can habitat selection affect the use of a wetland complex by waterbirds? Biodivers. Conserv. 2006, 15, 4569–4582. [Google Scholar] [CrossRef]
- Guadagnin, D.L.; Maltchik, L. Habitat and landscape factors associated with neotropical waterbird occurrence and richness in wetland fragments. In Vertebrate Conservation and Biodiversity; Springer: Berlin, Germany, 2006; pp. 405–418. [Google Scholar]
- Root, R.B. The niche exploitation pattern of the blue-gray gnatcatcher. Ecol. Monogr. 1967, 37, 317–350. [Google Scholar] [CrossRef]
- Blake, J.G. Trophic structure of bird communities in forest patches in east-central Illinois. Wilson Bull. 1983, 416–430. [Google Scholar]
- Bradley, R.A.; Bradley, D.W. Co-occurring groups of wintering birds in the lowlands of southern California. Auk 1983, 100, 491–493. [Google Scholar]
- Ma, Z.; Cai, Y.; Li, B.; Chen, J. Managing wetland habitats for waterbirds: An international perspective. Wetlands 2010, 30, 15–27. [Google Scholar] [CrossRef]
- Wong, M. Trophic organization of understory birds in a Malaysian dipterocarp forest. Auk 1986, 103, 100–116. [Google Scholar]
- Riffell, S.K.; Gutzwiller, K.J.; Anderson, S.H. Does repeated human intrusion cause cumulative declines in avian richness and abundance? Ecol. Appl. 1996, 6, 492–505. [Google Scholar] [CrossRef]
- Canterbury, G.E.; Martin, T.E.; Petit, D.R.; Petit, L.J.; Bradford, D.F. Bird communities and habitat as ecological indicators of forest condition in regional monitoring. Conserv. Biol. 2000, 14, 544–558. [Google Scholar] [CrossRef]
- Skowno, A.; Bond, W. Bird community composition in an actively managed savanna reserve, importance of vegetation structure and vegetation composition. Biodivers. Conserv. 2003, 12, 2279–2294. [Google Scholar] [CrossRef]
- Recher, H.; Gowing, G.; Kavanagh, R.; Shields, J.; Rohan-Jones, W. Birds, resources and time in a tablelands forest. Ecol. Soc. Aust. 1983, 12, 101–123. [Google Scholar]
- Andersen, A. Functional groups and patterns of organization in North American ant communities: A comparison with Australia. J. Biogeogr. 1997, 24, 433–460. [Google Scholar] [CrossRef]
- Watson, J.; Watson, A.; Paull, D.; Freudenberger, D. Woodland fragmentation is causing the decline of species and functional groups of birds in southeastern Australia. Pac. Conserv. Biol. 2002, 8, 261–270. [Google Scholar] [CrossRef]
- Keller, A.; Rödel, M.O.; Linsenmair, K.E.; Grafe, T.U. The importance of environmental heterogeneity for species diversity and assemblage structure in Bornean stream frogs. J. Anim. Ecol. 2009, 78, 305–314. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, T.J.; Jackson, L.E.; Brooks, R.P. Bird guilds as indicators of ecological condition in the central Appalachians. Ecol. Appl. 2000, 10, 1706–1721. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- Plotkin, J.B.; Chave, J.; Ashton, P.S. Cluster analysis of spatial patterns in Malaysian tree species. Am. Nat. 2002, 160, 629–644. [Google Scholar] [CrossRef] [PubMed]
- Dormann, C.F.; McPherson, J.M.; Araújo, M.B.; Bivand, R.; Bolliger, J.; Carl, G.; Davies, R.G.; Hirzel, A.; Jetz, W.; Kissling, W.D. Methods to account for spatial autocorrelation in the analysis of species distributional data: A review. Ecography 2007, 30, 609–628. [Google Scholar] [CrossRef]
- Buckley, G.; Forbes, J. Ecological evaluation using biological habitats an appraisal. Landsc. Plan. 1979, 5, 263–280. [Google Scholar] [CrossRef]
- Palmer, M.W.; White, P.S. Scale dependence and the species-area relationship. Am. Nat. 1994, 144, 717–740. [Google Scholar] [CrossRef]
- Gregory, R.D.; Van Strien, A.; Vorisek, P.; Gmelig Meyling, A.W.; Noble, D.G.; Foppen, R.P.; Gibbons, D.W. Developing indicators for European birds. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Beintema, A. Meadow birds as indicators. In Ecological Indicators for the Assessment of the Quality of Air, Water, Soil, and Ecosystems; Springer: Berlin, Germany, 1983; pp. 391–398. [Google Scholar]
- Weiher, E.; Clarke, G.P.; Keddy, P.A. Community assembly rules, morphological dispersion, and the coexistence of plant species. Oikos 1998, 81, 309–322. [Google Scholar] [CrossRef]
- Schulze, C.H.; Waltert, M.; Kessler, P.J.; Pitopang, R.; Veddeler, D.; Mühlenberg, M.; Gradstein, S.R.; Leuschner, C.; Steffan-Dewenter, I.; Tscharntke, T. Biodiversity indicator groups of tropical land-use systems: Comparing plants, birds, and insects. Ecol. Appl. 2004, 14, 1321–1333. [Google Scholar] [CrossRef]
- Emlen, J.T. Size and structure of a wintering avian community in southern Texas. Ecology 1972, 53, 317–329. [Google Scholar] [CrossRef]
- Karr, J.R. Structure of avian communities in selected Panama and Illinois habitats. Ecol. Monogr. 1971, 41, 207–233. [Google Scholar] [CrossRef]
- Koskiaho, J. Flow velocity retardation and sediment retention in two constructed wetland–ponds. Ecol. Eng. 2003, 19, 325–337. [Google Scholar] [CrossRef]
- Hansson, L.A.; Brönmark, C.; Anders Nilsson, P.; Åbjörnsson, K. Conflicting demands on wetland ecosystem services: Nutrient retention, biodiversity or both? Freshw. Biol. 2005, 50, 705–714. [Google Scholar] [CrossRef]
- Taoyuan City Government. Taoyuan Comprehensive Plan; Taoyuan City Government: Taoyuan, Taiwan, 1997.
- Fang, W.-T.; Chu, H.-J.; Cheng, B.-Y. Modeling waterbird diversity in irrigation ponds of Taoyuan, Taiwan using an artificial neural network approach. Paddy Water Environ. 2009, 7, 209–216. [Google Scholar] [CrossRef]
- Paracuellos, M.; Tellería, J.L. Factors affecting the distribution of a waterbird community: The role of habitat configuration and bird abundance. Waterbirds 2004, 27, 446–453. [Google Scholar] [CrossRef]
- Fang, T. The study of the spatial structure change of water land in Taoyuan terrace. Master’s Thesis, Department of Bioenvironmental Systems Engineering, National Taiwan University, Taipei, Taiwan, 2001. [Google Scholar]
- Bookhout, T.A. Research and Management Techniques for Wildlife and Habitats; Wildlife Society: Bethesda, MD, USA, 1996. [Google Scholar]
- Childs, C. Interpolating surfaces in ArcGIS spatial analyst. ArcUser 2004, 3235, 569. [Google Scholar]
- McGarigal, K.; Cushman, S.A.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps. Computer Software Program Produced by the Authors at the University of Massachusetts, Amherst. 2012. Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 21 November 2018).
- Cronk, B.C. How to Use SPSS®: A Step-by-Step Guide to Analysis and Interpretation; Routledge: Abingdon, UK, 2017. [Google Scholar]
- Tryon, R.C. Cluster Analysis: Correlation Profile and Orthometric (Factor) Analysis for the Isolation of Unities in Mind and Personality; Edwards Brother, Incorporated, Lithoprinters and Publishers: Ann Arbor, MI, USA, 1939. [Google Scholar]
- Oppel, S.; Schaefer, H.M.; Schmidt, V.; Schröder, B. Habitat selection by the pale-headed brush-finch (Atlapetes pallidiceps) in southern Ecuador: Implications for conservation. Biol. Conserv. 2004, 118, 33–40. [Google Scholar] [CrossRef]
- Wilks, D.S. Cluster analysis. In International Geophysics; Elsevier: Amsterdam, The Netherlands, 2011; Volume 100, pp. 603–616. [Google Scholar]
- SAS Institute. The SAS System for Windows, Release 8.0; SAS Institute: Cary, NC, USA, 1999.
- Kauppinen, J. Temporal variation in the wetland waterfowl communities of the Finnish lake district. Ornis Fennica 1995, 7, 145–153. [Google Scholar]
- Little, R.; Crowe, T. Conservation implications of deciduous fruit farming on birds in the Elgin district, Western Cape Province, South Africa. Trans. R. Soc. S. Afr. 1994, 49, 185–198. [Google Scholar] [CrossRef]
- Hockey, P. Atlas of the Birds of the Southwestern Cape; Cape Bird Club: Clareinch, South Africa, 1989. [Google Scholar]
- Wood, P.J.; Greenwood, M.T.; Agnew, M. Pond biodiversity and habitat loss in the UK. Area 2003, 35, 206–216. [Google Scholar] [CrossRef]
- Raebel, E.M.; Merckx, T.; Feber, R.E.; Riordan, P.; Thompson, D.J.; Macdonald, D.W. Multi-scale effects of farmland management on dragonfly and damselfly assemblages of farmland ponds. Agric. Ecosyst. Environ. 2012, 161, 80–87. [Google Scholar] [CrossRef]
- Boothby, J. Pond conservation: Towards a delineation of pondscape. Aquat. Conserv. Mar. Freshw. Ecosyst. 1997, 7, 127–132. [Google Scholar] [CrossRef]
- Durrett, R.; Levin, S. Spatial models for species-area curves. J. Theor. Biol. 1996, 179, 119. [Google Scholar] [CrossRef]
- Gaston, K.J.; Blackburn, T.M.; Lawton, J.H. Interspecific abundance-range size relationships: An appraisal of mechanisms. J. Anim. Ecol. 1997, 579–601. [Google Scholar] [CrossRef]
- McGill, B.J. A test of the unified neutral theory of biodiversity. Nature 2003, 422, 881. [Google Scholar] [CrossRef]
- Ney-Nifle, M.; Mangel, M. Species–area curves based on geographic range and occupancy. J. Theor. Biol. 1999, 196, 327–342. [Google Scholar] [CrossRef]
- Kingsland, S.E. Creating a science of nature reserve design: Perspectives from history. Environ. Model. Assess. 2002, 7, 61–69. [Google Scholar] [CrossRef]
- Harte, J.; Kinzig, A.P. On the implications of species-area relationships for endemism, spatial turnover, and food web patterns. Oikos 1997, 80, 417–427. [Google Scholar] [CrossRef]
- Traut, A.H.; Hostetler, M.E. Urban lakes and waterbirds: Effects of shoreline development on avian distribution. Landsc. Urban Plan. 2004, 69, 69–85. [Google Scholar] [CrossRef]
- Fox, A.; Kahlert, J.; Ettrup, H. Diet and habitat use of moulting Greylag Geese Anser anser on the Danish island of Saltholm. Ibis 1998, 140, 676–683. [Google Scholar] [CrossRef]
- Mathiasson, S. A moulting population of non-breeding Mute Swans with special reference to flight-feather moult, feeding ecology and habitat selection. Wildfowl 1973, 24, 43–53. [Google Scholar]
- Li, H.; Reynolds, J.F. A simulation experiment to quantify spatial heterogeneity in categorical maps. Ecology 1994, 75, 2446–2455. [Google Scholar] [CrossRef]
- Leitao, A.B.; Ahern, J. Applying landscape ecological concepts and metrics in sustainable landscape planning. Landsc. Urban Plan. 2002, 59, 65–93. [Google Scholar] [CrossRef]
- Fang, W.-T.; Cheng, B.-Y.; Shih, S.-S.; Chou, J.-Y.; Otte, M.L. Modelling driving forces of avian diversity in a spatial configuration surrounded by farm ponds. Paddy Water Environ. 2016, 14, 185–191. [Google Scholar] [CrossRef]
- Fang, W.-T.; Chou, J.-Y.; Lu, S.-Y. Simple patchy-based simulators used to explore pondscape systematic dynamics. PLoS ONE 2014, 9, e86888. [Google Scholar] [CrossRef] [PubMed]



| Item Acronym | Pondscape Variables (Metrics/Units) | Description |
|---|---|---|
| PS b | Pond size 1 | Pond size (m2) |
| FCA b | Foliage canopy area next to waterfront edge of a pond (m2) 1 | Boundary delineation of disturbance |
| MA a,b | Mudflat area in a pond (m2) 1 | Boundary delineation of disturbance |
| WASA a,b | Water surface area in a pond (m2) 1 | Boundary delineation of disturbance |
| %FCA b | FCA ÷ PS 1 | |
| %MA b | MA ÷ PS 1 | |
| %FARM b | The ratio of farmland areas within a radius of 100 ha from the pond’s geometric center (m2)/ha 1 | Pondscape isolation or connectivity from the center of each selected pond |
| %BUILD b | The ratio of permanent building areas within a radius of 100 ha from the pond’s geometric center (m2)/ha 1 | Pondscape isolation from center of each selected pond |
| %PONDS b | The ratio of multiple pond areas within a radius of 100 ha from the pond’s geometric center (m2)/ha 1 | Pondscape connectivity from the center of each selected pond |
| %RIVER b | The ratio of all watercourse areas covered by rivers, creeks, channels, and ditches within a radius of 100 ha from the pond’s geometric center (m2)/ha 1 | Pondscape connectivity from the center of each selected pond |
| %ROAD b | The ratio of all road and trail areas within a radius of 100 ha from the pond’s geometric center (m2)/ha 1 | Pondscape isolation from the center of each selected pond |
| Species | Occurrence Frequency | Occurrence Rate | Type of the Birds |
|---|---|---|---|
| Black-crowned night heron (Nycticorax nycticorax) | 2363 | 15.7% | Resident |
| Little egret (Egretta garzetta) | 1883 | 12.5% | Resident |
| Grey heron (Ardea cinerea) | 1829 | 12.2% | Wintering visitor |
| Light-vented bulbul (Pycnonotus sinensis) | 1575 | 10.5% | Resident |
| Eurasian tree sparrow (Passer montanus) | 1125 | 7.7% | Resident |
| Great egret (Casmerodius alba) | 726 | 4.8% | Wintering visitor |
| Red-collared dove (Streptopelia tranquebarica) | 509 | 3.4% | Resident |
| Japanese white-eye (Zosterops japonica) | 504 | 3.3% | Resident |
| Little ringed plover (Charadrius dubius) | 316 | 2.1% | Wintering visitor |
| Little grebe (Tachybaptus ruficollis) | 304 | 2% | Resident |
| Total | 11,134 | 74.2% |
| Classification | November 2003 | December 2003 | January 2004 | February 2004 |
|---|---|---|---|---|
| Aerial feeder | 96 (5) | 248 (7) | 90 (6) | 79 (4) |
| Waterfowl | 85 (6) | 209 (6) | 157 (7) | 132 (5) |
| Shorebirds | 240 (6) | 261 (10) | 212 (10) | 94 (6) |
| Waterside birds | 2192 (10) | 1776 (14) | 1775 (11) | 1465 (15) |
| Grassland birds | 31 (4) | 127 (3) | 9 (2) | 12 (4) |
| Bushland birds | 233 (11) | 213 (9) | 354 (9) | 296 (8) |
| Woodland birds | 844 (18) | 1438 (18) | 1303 (17) | 1082 (17) |
| Individual no. (species richness) | 3721 (60) | 4272 (67) | 3900 (62) | 3160 (59) |
| Classification | Aerial Feeders | Waterfowl | Shorebirds | Waterside Birds | Landbirds 1 |
|---|---|---|---|---|---|
| PS | −0.073 | 0.259 ** | 0.102 | 0.224 ** | −0.100 |
| FCA | −0.006 | 0.305 ** | 0.140 | 0.226 ** | 0.078 |
| MA | −0.076 | 0.406 ** | 0.364 ** | 0.021 | −0.108 |
| WASA | −0.048 | 0.112 | −0.043 | 0.236 ** | −0.063 |
| %FCA | 0.022 | −0.026 | 0.043 | −0.185 ** | −0.057 |
| %MA | −0.033 | 0.387 ** | 0.408 ** | 0.063 | −0.094 |
| %FARM | −0.039 | −0.104 | 0.006 | 0.208 ** | 0.044 |
| %BUILD | 0.109 | 0.016 | −0.059 | −0.292 ** | −0.028 |
| %PONDS | −0.197 ** | 0.162 * | 0.133 | 0.199 ** | −0.145 |
| %RIVER | 0.172 * | 0.163 * | 0.039 | −0.111 | 0.102 |
| %ROAD | 0.020 | −0.057 | −0.122 | −0.052 | −0.135 |
| Classification | Aerial Feeders | Waterfowl | Shorebirds | Waterside Birds | Landbirds 1 |
|---|---|---|---|---|---|
| PS | −0.059 | 0.193 ** | 0.091 | 0.272 ** | −0.059 |
| FCA | −0.002 | 0.503 ** | 0.007 | 0.205 ** | 0.140 |
| MA | −0.054 | 0.347 | 0.291 ** | 0.114 | −0.084 |
| WASA | −0.041 | 0.064 | −0.024 | 0.249 ** | −0.029 |
| %FCA | −0.034 | −0.046 | 0.005 | −0.120 | −0.018 |
| %MA | −0.021 | 0.398 ** | 0.367 ** | 0.184 * | −0.058 |
| %FARM | −0.091 | −0.005 | 0.011 | 0.169 * | 0.076 |
| %BUILD | 0.099 | −0.069 | −0.036 | −0.237 ** | −0.010 |
| %PONDS | −0.093 | 0.171 * | 0.075 | 0.189 * | −0.075 |
| %RIVER | 0.256 ** | 0.095 | −0.018 | −0.117 | −0.034 |
| %ROAD | 0.035 | −0.151 * | −0.064 | −0.110 | −0.199 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, C.-H.; Chou, J.-Y.; Fang, W.-T. Habitat Selection of Wintering Birds in Farm Ponds in Taoyuan, Taiwan. Animals 2019, 9, 113. https://doi.org/10.3390/ani9030113
Hsu C-H, Chou J-Y, Fang W-T. Habitat Selection of Wintering Birds in Farm Ponds in Taoyuan, Taiwan. Animals. 2019; 9(3):113. https://doi.org/10.3390/ani9030113
Chicago/Turabian StyleHsu, Chia-Hsuan, Jui-Yu Chou, and Wei-Ta Fang. 2019. "Habitat Selection of Wintering Birds in Farm Ponds in Taoyuan, Taiwan" Animals 9, no. 3: 113. https://doi.org/10.3390/ani9030113
APA StyleHsu, C.-H., Chou, J.-Y., & Fang, W.-T. (2019). Habitat Selection of Wintering Birds in Farm Ponds in Taoyuan, Taiwan. Animals, 9(3), 113. https://doi.org/10.3390/ani9030113