1. Introduction
Biodynamic farming was one of the earliest movements in organic agriculture [
1]. Biodynamic agriculture developed its own additional worldwide standards in addition to the regulations for organic agriculture. Biodynamics maintains the ecological principle of the farm as an organism. This limits the farmer’s ability to intensify the yield per hectare, as well as per animal, and the cycling of fodder and manure should be fulfilled. With regard to animal production, animals are kept according to their specific integrity. This means that ruminating animals are largely fed with roughage, not concentrates, whereas hay should always be part of the diet [
2,
3]. In comparison to conventional farming systems, the milk production per animal and per hectare is lower [
4,
5]. Biodynamic dairy production can be found in different areas of Europe; however, it is unknown whether the region of production impacts the farmer’s choices in a similar way to conventional farming, and how this affects the milk’s fatty acid (FA) composition. Farmers adapt their systems, their grazing policy, the length of the grazing period, and their choices for additional fodder according to the local climate conditions.
The level of n3 FA was found to differ between organic milk and conventional milk. The separation was more pronounced when the region of production was taken into account and stable carbon isotopes were integrated [
6]. In the comparison of the FA composition of conventional and biodynamic milk in southern Germany, the differentiation between the farm origin (conventional versus biodynamic) depended on the level of intensification (low-input versus high-input). The authors showed that there was a complete separation between low-input biodynamic and high-input conventional milk FA composition. However, the milk FA patterns from high-input biodynamic production and low-input conventional production were overlapping [
5]. The different intakes of fresh green fodder between conventional and organic cows played an important role, affecting the improved FA profile in organic milk fat (more: n3 FA, conjugated linolenic acids, vaccenic acid; less: palmitic acid) [
7]. In grass-based milk production in New Zealand, differences in the milk’s FA composition depended on the intake of grass rather than on the label of production [
8]. The importance of fresh grass intake was confirmed in recent studies in the US. Grass-based farming systems had more favorable FA profiles in terms of the conjugated linoleic acid (CLA) and n6/n3 polyunsaturated FA (PUFA) ratio compared to organic systems, due to the principle of roughage-based farming without additional concentrates [
9]. If farmers produced milk from fresh grass and grass-based winter forage, a mean yearly n6/n3 PUFA ratio of 0.99 was reached. From a health perspective, westernized diets are overloaded with n6 FA and contain suboptimal levels of n3 PUFA. The conventional milk produced in feedlots can reach an n6/n3 FA ratio of 5.8, according to Reference [
9].
Farming systems are very different across Europe. Soil type, slope, and the local climate impact the structure of the farm in terms of crops, grazing policy or length of the grazing season [
10]. An increased distance to the Atlantic Ocean results in greater temperature extremes (day/night, summer/winter) and a shorter growing season [
11]. Typical grassland areas in Europe are found in the coastal areas, because of the mild climate and higher precipitation, as well as in the mountainous and alpine regions [
12]. The impact of the abiotic circumstances, climate conditions, and crop growth is also reflected in the natural vegetation zones across Europe [
13,
14], and it is assumed that both the natural vegetation and the farming system are affected by this. The aim of this study was to analyze the differences in the milk FA composition of biodynamic farms in Central and Northern Europe and to correlate the FA patterns with the geological and ecological background data of the farm location. A second aim was to compare biodynamic shop milk with conventional shop milk. The n6/n3 PUFA ratio of milk was taken as the main indicator as described in Reference [
9] to estimate the impact of the label of production (biodynamic versus conventional).
2. Materials and Methods
Bulk tank milk samples: From different research projects conducted from 2008 to 2015, cow’s bulk tanker milk (N = 163) was sampled in biodynamic farms (N = 41). Samples were immediately cooled at 4 °C, transported, and frozen within 48 hours. All samples were analyzed at the Institute of Nutritional Sciences of the Friedrich Schiller University in Jena, following the same protocol over a period of 5 years. Bulk tank milk samples from published data [
5,
15,
16] and unpublished data [
17] were used for this purpose.
Shop milk samples: In September 2010, shop milk from both biodynamic (N = 10) and conventional farms (N = 10) was sampled in Sweden, Denmark, Germany, The Netherlands, and Switzerland. Samples were cooled at 4 °C and were frozen within 48 hours. All samples were analyzed at the Institute of Nutritional Sciences of the Friedrich Schiller University in Jena. Based on the creamery that delivered the shop milk, the location of processing in terms of elevation, latitude, and altitude was connected with the FA outcome.
Lipid extraction and preparation of fatty acid methyl esters (FAME) and analysis by gas chromatography (GC): Frozen milk samples were thawed at room temperature and freeze-dried (fd60-1; Pharma & Food, Dresden, Germany). Milk powder was used for Soxhlet extraction using a Soxtherm2000 S306 A (Sartorius, Göttingen, Germany). FAME was prepared using NaOCH
3. GC procedures were used to resolve all FA as described in Reference [
18]. Briefly, for the separation of C4–C22, a fused-silica capillary column of medium polarity was used (GC-17 V3, Shimadzu, Kyoto, Japan; DB-225 MS: 60 m × 0.25 mm i.d., with 0.25 μm film thickness, Agilent Technologies, Palo Alto, CA, USA). Oven temperature was initially maintained for 2 min at 70 °C, then it was increased by 10 °C min
−1 to 180 °C, and then further increased by 2 °C min
−1 to 220 °C, and held for 5 min. In the final step, the temperature was increased by 2 °C min
−1 to 230 °C and held for 27 min. The cis and trans isomers of C18:1 were separated using a fused-silica capillary column of high polarity (GC-2010 plus, Shimadzu; CP-select for FAME; 200 m × 0.25 mm i.d., with 0.25 μm film thickness, Varian, Houten, Netherlands). These isomers were separated under isothermal conditions at 176 °C. For the GC analysis, 1 μL of 2% FAME in n-hexane was injected with a split ratio of 1:100. For both procedures, the injector and detector temperatures were maintained at 260 and 270 °C, respectively. The carrier gas was hydrogen.
Farm ecology and geography: All the farms under study were located in Europe. Abiotic farm environmental and climate data were derived from the ecological vegetation zones in Europe described in References [
13,
19]. The farm location was pinpointed in Google maps, and longitude, altitude, and elevation above sea level were notified. The map of the environmental stratification of Europe (EnS), which included 84 environmental strata [
12], was connected with the farm location. For each of the 84 strata, monthly values were present for five climate variables over the period from 1900 to 2000 [
12,
13]. For the purpose of this study, mean values of the following three variables over the period from 1960 to 1990 were used:
- -
Monthly precipitation and year sum (mm);
- -
Monthly average temperature and year mean (°C);
- -
The monthly number of sun hours and year sum (N).
In addition, agronomic indicators were present for:
- -
The length of the growing season (N number of days warmer than 5 °C), and
- -
The annual temperature (°C) sums expressed as growing degree days with a 0 °C base (GDD
0), as described in Reference [
12].
Statistics: Single farm milk samples (N = 163) were obtained from 41 different farms. The number of samples per farm ranged between 1 and 27. Based on the date of sampling, milk was divided into summer (April to October) or winter (November to March) samples. This separation was further adjusted based on additional information of the cow’s rations, if present. All statistical calculation was done using SPSS version 20 (IBM SPSS Statistics for Mac, IBM Corporation, Armonk, NY, USA). The main data exploration was based on a principal component analysis following a stepwise linear regression. Based on the Scree plot structure, and after eliminating single FA with an expression lower than 0.3, four components were derived after Varimax rotation. The impact of the region of production was shown in a bi-plot. The values of the three main regions of production were rescaled based on the explained variance within each component. The linear data from the environmental background were correlated with the expressed factor scores of the four components based on a Pearson correlation and two-tailed significance. There were no fodder data present for each single milk sample. Based on the existing information between the correlation of fresh grass intake and the n6/n3 PUFA ratio in milk as in Reference [
5], the amount of fresh grass intake was estimated for each of the milk samples.
Shop milk samples from biodynamic and conventional origins were compared based on a paired T-test. Results were considered significant if p ≤ 0.05.
4. Discussion
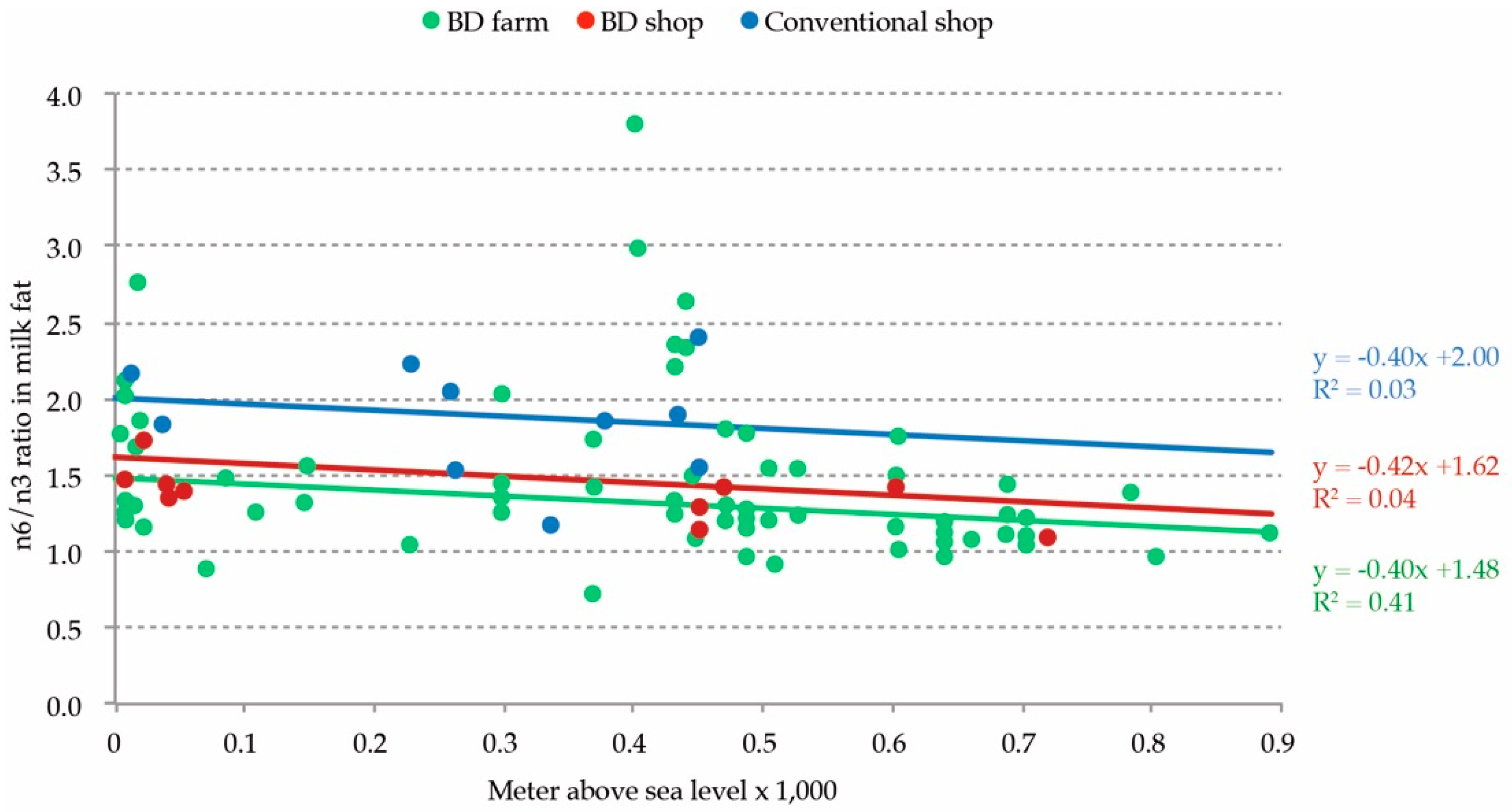
In this study, the FA composition of 163 biodynamic milk samples taken across Europe was analyzed. The patterns in the FA composition were correlated with the ecological and climate data of the farm environment. Based on a principal component analysis, 63% of the variance was explained by four components. The different FA patterns could be correlated with the season of production, precipitation level, the farm elevation, and the length of the growing season. In a small set of paired shop milk samples, a more favorable FA profile in relation to health was found in the biodynamic milk compared with the conventional milk. The ratio n6/n3 PUFA in both the biodynamic shop milk and biodynamic farm milk had a similar correlation with the elevation level of production, whilst the conventional shop milk was different. Based on the n6/n3 ratio, biodynamic milk was based on a >82% roughage and grass intake by the cows.
For the grass growth across Europe, productivity was closely related to the annual precipitation and to a lesser extent to the mean temperature and length of the growing season [
20]. Climate, soil type, slope, and elevation affected the farmer’s choice of the type of pasture, regular grassland renewal, and crop rotation. The growing pattern of the grassland and the start of the growing season depended on the climatic conditions. Early spring growth or even winter growth is found in the southern regions in Europe, in lower elevated areas, and in areas affected by the Gulf Stream (mild, rainy). To keep cows grazing during the summer, continuous growth of grass is necessary and is connected with a high amount of precipitation. Typical European grassland areas are found on the British continent, the coastal areas in Denmark, Northern-Germany, and The Netherlands, Normandy (France), Galicia (Spain), and the Alpine regions [
20]. The continental climate is the typical area for mixed farming, where besides grass-legumes mixtures, cows can be fed with annual fodder crops like maize silage or whole plant silage. A high daily summer temperature forces farmers to reduce the cow’s grazing time. The growing conditions for fresh grass are poor due to the low precipitation and high evaporation and temperatures. Moreover, dairy cows reach the upper-temperature limit of their comfort zone, and therefore, are kept indoors [
21]. When kept indoors, cows will often be fed with conserved forages and concentrates rather than fresh grass. In combination with a short grazing season, especially in more Nordic European regions, cows have a long winter period inside and rely on conserved forages and concentrates.
The highest precipitation and most sun hours were in the Pre-Alpine area in combination with the lowest average daily temperature. The climate of the Central region, together with the Pre-Alpine climate, led to the shortest growing season. In the Central region, the grass growth can be limited in the middle of the season, especially when the summer precipitation is low and temperatures are high. In the Pre-Alpine region in spring, there is a high speed of grass growth, after a relatively late start of the growing season. The time of this burst of grass growth takes place in relation to the elevation of the land. Typical also for the Pre-Alpine region is the high precipitation during the growing season. Comparable to the Atlantic area with its high precipitation, the choice of annual fodder plants is undesirable in the Pre-Alpine regions, also due to the slope of the fields. Particularly, in the Pre-Alpine area, farmers depend on permanent pastures, often rich in plant species [
22].
Several authors [
5,
8,
9] discussed the question of whether an organic label of production had more impact on the FA composition and specific FA markers (n3, n6/n3, CLA, and its precursors) than the overall grazing and feeding policy at the farm. A complete separation of farming systems was only possible if conventional farms had a high-input strategy and biodynamic farms a low-input strategy [
5]. However, at grass-based farms in New Zealand, conventional farms produced a more favorable FA composition in terms of CLAc9t11 and its precursor C18:1t11 than organic farms [
8], even though organic cows had more access to grass than conventional cows. In fresh grass, there is a linear relationship between the ALA concentration and the protein concentration [
23]. Therefore, the explanation for the unexpected difference in New Zealand milk CLA levels might be due to the higher speed of grass growth after the grass received nitrogen fertilizer at the conventional farms [
8]. Not only the access to fresh grass but also the constant creation of new plant cells affect the final level of CLAc9t11 and C18:1t11 in milk fat. In Reference [
16], a high content of phytanic acid in the milk fat was a better prediction of the fresh grass intake. The concentration of phytanic acid was under the direct influence of the chlorophyll content of the cow’s feed, and a further differentiation could be made between fresh grass and grass silage on the one hand and dried hay on the other hand. A strong reciprocal correlation was found between the sum amount of concentrates plus maize silage in the cow’s diet and the phytanic acid level [
16].
In this study, after the principal component analysis we have found four components to differentiate the 163 biodynamic milk samples. Other authors have expressed the total explained variance across only the first two, main components [
5,
8,
24,
25]. In Reference [
25], the milk from French farms in a mountainous area in summer and winter was investigated, whereas, in Reference [
8], the impact of the season and label (organic versus conventional) on the milk FA composition at New Zealand farms was compared. In Reference [
25], the main separation was based on the seasonal impact, which was similar in our study. Grass feeding (summer) versus feeding of conserved forages (winter) increased the levels of C18:1c9 and C18:1t11, in general, all trans-isomers of C18:1, CLAc9t11, and C18:0. In our four-dimensional solution, the effect of a higher fresh grass intake was reflected in the outcomes of the components one and four covering the differentiation of summer and winter milk, respectively, and milk from the Pre-Alpine and Central regions. The amount of precipitation was an important abiotic factor differentiating these results, and especially in component four (Pre-Alpine versus Central), where the correlation was high. This implies that when water becomes restrictive for grass growth, like in the Central region, biodynamic farmers start feeding other forages and concentrates for maintaining their milk yield. In Reference [
25], a correlation between a higher altitude in the mountains and the presence of permanent pastures was found. For our milk samples, the elevation was positively correlated with component one and negatively with components two and four, and the typical ‘Pre-Alpine effect’ was reflected in components two and four. If cows were only consuming semi-permanent forages and low amounts of concentrate [
25], they would produce milk with the highest concentration of CLAc9t11 and ALA. This was reflected in component four.
According to Reference [
25], the access to permanent pastures was correlated with the group of iso- and ante-iso FA from C15:0 and C17:0, as well as C18:3c9c12c15 (ALA) and C20:5n-3. This differentiation was in concordance with component three, differentiating the milk from the Atlantic and Central region. According to Reference [
25], the branched-chain FA was not affected by the season, which was confirmed by our data. Based on these outcomes, permanent pastures, as well as access to grassland and grazing, is limited in Northern and Continental Europe. In Reference [
26], a correlation between the increased amount of C14iso and the decrease of concentrates in the diet could be calculated. C14iso is part of the third component separating milk from the Atlantic and Central regions, suggesting higher concentrate levels at the Central farms to compensate for a lower forage intake.
In Reference [
25], the intake of hay was solely correlated with C15:0. C15:0 is part of the third component, suggesting a higher intake of hay at the Atlantic farms compared to the Central farms. They showed that feeding of grass silage and concentrates was correlated with C12:0 and C14:0, C14:1c9 and C16:0, respectively. These results were found in component one, where they built the negative direction of the outcome, in accordance with the winter fodder situation. C16:0 was found to be a strong separator between the summer-grazing and winter-feeding period, which was reflected in the strong negative correlation of C16:0 in component one (−0.921). In Reference [
25], maize silage was solely correlated with C18:1c12. In our analysis, C18:1c12 was found in relation to components one, three, and four. In components 3 three and four, C18:1c12 was found in connection with milk from the Central area. Reduced precipitation is one of the motivations for farmers to grow fodder maize. Farms relied more on a crop rotation of fodder plants, like whole plant silage, maize silage, and grass-legume silage as winter fodder, but also as supplementation for conserved fodders during summer. Biodynamic high input farms in the German Allgäu region had a reduced grass intake during summer in contrast to the biodynamic low-input farms [
5]. Cows on biodynamic high-input farms produced more milk based on the supplemental feed from concentrates, grass silage, and maize silage compared to the biodynamic low-input farms. In addition, low-input farms were more dependent on roughage [
15], and only used a limited amount of concentrates, as well as some hay [
5].
Our results showed that biodynamic farm milk had a mean ratio n6/n3 PUFA in summer of 1.45 and 1.64 in the winter. When farmers have low access to grass and grazing, and when they depend on high yielding cows, they adapt the cow’s diet and apply concentrates, maize, and other conserved fodder rather than grass only. This choice impacts the n6/n3 ratio, which increased rapidly [
16,
23]. In a smaller sample set of milk samples taken in the German Allgäu region, all the conventional farms— low-input farms and high-input farms—fed higher amounts of concentrates per cow, and biodynamic farms were more dependent on roughage [
5]. The roughage-based character of biodynamic milk production in other areas of Europe was confirmed in the present study. Based on the average n6/n3 ratio, the FA quality of biodynamic summer milk (1.45) was quite near to the FA quality of the US systems based on pure grazing (0.99), although there was quite some difference between the lowest and the highest values (
Table 3).

 ,
, 




{kind=link}
{kind=link}