Replacement of Fish Meal by Defatted Yellow Mealworm (Tenebrio molitor) Larvae in Diet Improves Growth Performance and Disease Resistance in Red Seabream (Pargus major)

 ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Feed Ingredients
2.2. Experimental Diets
2.3. Feeding Trials
2.3.1. Growth Test
- FL gain (mm) = FL at trial end − initial FL,
- BW gain (g) = BW at trial end − initial BW,
- FL gain rate (%) = FL gain/initial FL × 100,
- BW gain rate (%) = BW gain/initial BW × 100,
- SGR, % day−1 = [(ln final BW – ln initial BW)/number of feeding days] × 100,
- Feed intake per fish (g) = total feed intake per group (g)/number of fish,
- FCR = total feed intake per group (g)/BW gain per group.
2.3.2. Challenge Test with Edwardsiella tarda
2.4. Proximate Composition, Amino Acid, and Fatty Acid Analysis
2.5. Statistical Analysis
3. Results
3.1. Analysis of Amino Acids and Fatty Acids in Yellow Mealworm Larvae
3.2. Growth Performances of Red Seabream Fed Diets with Increasing Replacement of FM with DMW-1
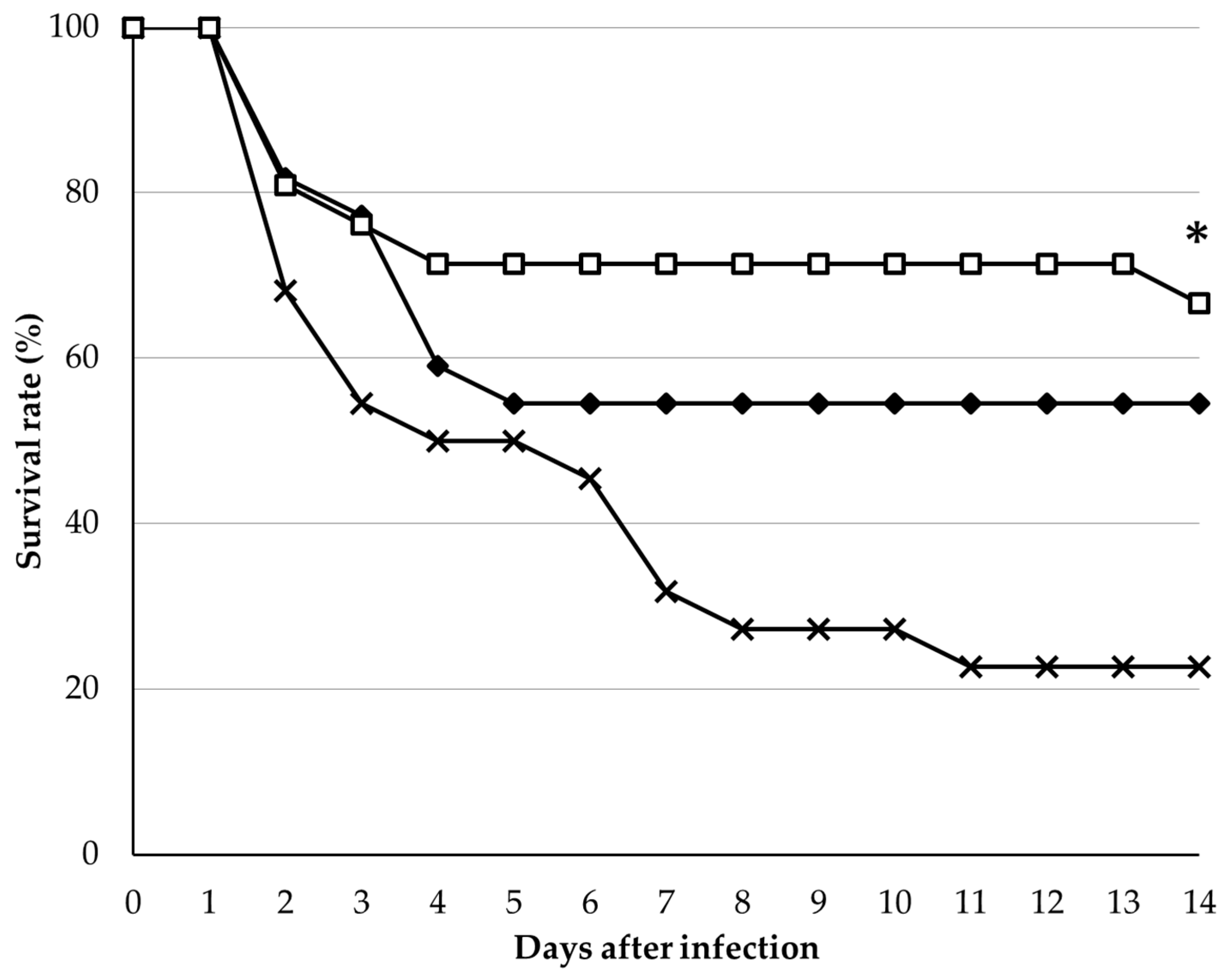
3.3. Challenge Test with Edwardsiella Tarda
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Huis, A.; Van Itterbeeck, J.; Harmke, K.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; ISBN 9789251075951. [Google Scholar]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Potential and challenges of insects as an innovative source for food and feed production. Innov. Food Sci. Emerg. Technol. 2013, 17, 1–11. [Google Scholar] [CrossRef]
- Sánchez-muros, M.; Barroso, F.G.; Manzano-agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the use of insects in the diet of farmed fish: Past and future. Anim. Feed Sci. Technol. 2015, 203, 1–22. [Google Scholar] [CrossRef]
- Piccolo, G.; Iaconisi, V.; Marono, S.; Gasco, L.; Loponte, R.; Nizza, S.; Bovera, F.; Parisi, G. Effect of Tenebrio molitor larvae meal on growth performance, in vivo nutrients digestibility, somatic and marketable indexes of gilthead sea bream (Sparus aurata). Anim. Feed Sci. Technol. 2017, 226, 12–20. [Google Scholar] [CrossRef]
- Gasco, L.; Henry, M.; Piccolo, G.; Marono, S.; Gai, F.; Renna, M.; Lussiana, C.; Antonopoulou, E.; Mola, P.; Chatzifotis, S. Tenebrio molitor meal in diets for European sea bass (Dicentrarchus labrax L.) juveniles: Growth performance, whole body composition and in vivo apparent digestibility. Anim. Feed Sci. Technol. 2016, 220, 34–45. [Google Scholar] [CrossRef]
- Iaconisi, V.; Marono, S.; Parisi, G.; Gasco, L.; Genovese, L.; Maricchiolo, G.; Bovera, F.; Piccolo, G. Dietary inclusion of Tenebrio molitor larvae meal: Effects on growth performance and final quality treats of blackspot sea bream (Pagellus bogaraveo). Aquaculture 2017, 476, 49–58. [Google Scholar] [CrossRef]
- Belforti, M.; Gai, F.; Lussiana, C.; Renna, M.; Malfatto, V.; Rotolo, L.; De Marco, M.; Dabbou, S.; Schiavone, A.; Zoccarato, I.; et al. Tenebrio Molitor Meal in Rainbow Trout (Oncorhynchus Mykiss) Diets: Effects on Animal Performance, Nutrient Digestibility and Chemical Composition of Fillets. Ital. J. Anim. Sci. 2015, 14, 4170. [Google Scholar] [CrossRef]
- Iaconisi, V.; Bonelli, A.; Pupino, R.; Gai, F.; Parisi, G. Mealworm as dietary protein source for rainbow trout: Body and fillet quality traits. Aquaculture 2018, 484, 197–204. [Google Scholar] [CrossRef]
- Song, S.G.; Chi, S.Y.; Tan, B.P.; Liang, G.L.; Lu, B.Q.; Dong, X.H.; Yang, Q.H.; Liu, H.Y.; Zhang, S. Effects of fishmeal replacement by Tenebrio molitor meal on growth performance, antioxidant enzyme activities and disease resistance of the juvenile pearl gentian grouper (Epinephelus lanceolatus ♂ × Epinephelus fuscoguttatus ♀). Aquac. Res. 2018, 49, 2210–2217. [Google Scholar] [CrossRef]
- Gasco, L.; Finke, M.; van Huis, A. Can diets containing insects promote animal health? J. Insects Food Feed 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Ido, A.; Iwai, T.; Ito, K.; Ohta, T.; Mizushige, T.; Kishida, T.; Miura, C.; Miura, T. Dietary effects of housefly (Musca domestica) (Diptera: Muscidae) pupae on the growth performance and the resistance against bacterial pathogen in red sea bream (Pagrus major) (Perciformes: Sparidae). Appl. Entomol. Zool. 2015, 50, 213–221. [Google Scholar] [CrossRef]
- Henry, M.A.; Gasco, L.; Chatzifotis, S.; Piccolo, G. Does dietary insect meal affect the fish immune system? The case of mealworm, Tenebrio molitor on European sea bass, Dicentrarchus labrax. Dev. Comp. Immunol. 2018, 81, 204–209. [Google Scholar] [PubMed]
- Henry, M.A.; Gai, F.; Enes, P.; Peréz-jiménez, A.; Gasco, L. Effect of partial dietary replacement of fishmeal by yellow mealworm (Tenebrio molitor) larvae meal on the innate immune response and intestinal antioxidant enzymes of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2018, 83, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Sankian, Z.; Khosravi, S.; Kim, Y.O.; Lee, S.M. Effects of dietary inclusion of yellow mealworm (Tenebrio molitor) meal on growth performance, feed utilization, body composition, plasma biochemical indices, selected immune parameters and antioxidant enzyme activities of mandarin fish (Siniperca scherzeri) juveniles. Aquaculture 2018, 496, 79–87. [Google Scholar]
- Koshio, S. Red sea bream, Pagrus major. In Nutrient Requirements and Feeding of Finfish for Aquaculture; Webster, C.D., Lim, C., Eds.; CABI Publishing: New York, NY, USA, 2002; pp. 51–62. ISBN 0851995195. [Google Scholar]
- Kim, E.J.; Utterback, P.L.; Parsons, C.M. Comparison of amino acid digestibility coefficients for corn, corn gluten meal, and corn distillers dried grains with solubles among 3 different bioassays. Poult. Sci. 2012, 91, 3141–3147. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis of AOAC International, 20th ed.; Latimer, W.G., Ed.; AOAC International: Gaithersburg, MD, USA, 2016; ISBN 0935584870. [Google Scholar]
- Forster, I.; Ogata, H.Y. Lysine requirement of juvenile Japanese flounder Paralichthys olivaceus and juvenile red sea bream Pagrus major. Aquaculture 1998, 161, 131–142. [Google Scholar] [CrossRef]
- Takagi, S.; Murata, H.; Goto, T.; Ichiki, T.; Endo, M.; Hatate, H.; Yoshida, T.; Sakai, T.; Yamashita, H.; Ukawa, M. Efficacy of taurine supplementation for preventing green liver syndrome and improving growth performance in yearling red sea bream Pagrus major fed low-fishmeal diet. Fish. Sci. 2006, 72, 1191–1199. [Google Scholar] [CrossRef]
- Takagi, S.; Murata, H.; Goto, T.; Hatate, H.; Endo, M.; Yamashita, H.; Miyatake, H.; Ukawa, M. Role of taurine deficiency in inducing green liver symptom and effect of dietary taurine supplementation in improving growth in juvenile red sea bream Pagrus major fed non-fishmeal diets based on soy protein concentrate. Fish. Sci. 2011, 77, 235–244. [Google Scholar] [CrossRef]
- El-Sayed, A.-F.M. Is dietary taurine supplementation beneficial for farmed fish and shrimp? a comprehensive review. Rev. Aquac. 2014, 6, 241–255. [Google Scholar] [CrossRef]
- National Agriculture and Food Research Organization. Standard Tables of Feed Composition in Japan, Japan Livestock Industry Association; National Agriculture and Food Research Organization: Tokyo, Japan, 2009; ISBN 978-4-901311-54-0. [Google Scholar]
- Kroeckel, S.; Harjes, A.G.E.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the Black Soldier Fly (Hermetia illucens) as fish meal substitute—Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364–365, 345–352. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef] [PubMed]
- Kono, M.; Matsui, T.; Shimizu, C. Effect of Chitin, Chitosan, and Cellulose as Diet Supplements on the Growth of Cultured Fish. Nippon Suisan Gakkaish 1987, 53, 125–129. [Google Scholar] [CrossRef]
- Ohta, T.; Ido, A.; Kusano, K.; Miura, C.; Miura, T. A Novel Polysaccharide in Insects Activates the Innate Immune System in Mouse. PLoS ONE 2014, 9, e114823. [Google Scholar] [CrossRef] [PubMed]
- Ohta, T.; Kusano, K.; Ido, A.; Miura, C.; Miura, T. Silkrose: A novel acidic polysaccharide from the silkmoth that can stimulate the innate immune response. Carbohydr. Polym. 2016, 136, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.F.Z.; Yasin, I.A.; Ohta, T.; Hashizume, A.; Ido, A.; Takahashi, T.; Miura, C.; Miura, T. The silkrose of Bombyx mori effectively prevents vibriosis in penaeid prawns via the activation of innate immunity. Sci. Rep. 2018, 8, 3–5. [Google Scholar] [CrossRef]
- Lee, J.; Kim, Y.-M.; Park, Y.-K.; Yang, Y.-C.; Jung, B.-G.; Lee, B.-J. Black soldier fly (Hermetia illucens) larvae enhances immune activities and increases survivability of broiler chicks against experimental infection of Salmonella Gallinarum. J. Vet. Med. Sci. 2018, 80, 736–740. [Google Scholar] [CrossRef]
- Thévenot, A.; Rivera, J.L.; Wilfart, A.; Maillard, F.; Hassouna, M.; Senga-Kiesse, T.; Le Féon, S.; Aubin, J. Mealworm meal for animal feed: Environmental assessment and sensitivity analysis to guide future prospects. J. Clean. Prod. 2018, 170, 1260–1267. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; van Broekhoven, S.; van Huis, A.; van Loon, J.J.A. Feed Conversion, Survival and Development, and Composition of Four Insect Species on Diets Composed of Food By-Products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ingredients (%) | 65% FM a | 25% MW b | 40% MW c | 65% MW d | 65% MW+ MO e |
| Fish meal | 65.00 | 40.00 | 25.00 | ||
| DMW-1 | 25.00 | 40.00 | 65.00 | 65.00 | |
| Fish oil | 4.00 | 5.00 | 6.00 | 7.00 | |
| Mealworm oil | 5.16 | ||||
| DHA | 1.84 | ||||
| Starch | 12.00 | 14.00 | 15.00 | 17.00 | 17.00 |
| Corn gluten meal | 8.00 | 5.00 | 3.00 | ||
| Taurine | 1.50 | 1.50 | 1.50 | 1.50 | 1.50 |
| Vitamin mix | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 |
| Mineral mix | 0.40 | 0.40 | 0.40 | 0.40 | 0.40 |
| Choline chloride | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| Vitamin C derivatives | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| NaH2PO4 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 |
| KH2PO4 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 |
| Calcium lactate | 1.50 | 1.50 | 1.50 | 1.50 | 1.50 |
| Carboxymethyl cellulose | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
| Proximate composition (% on a dry matter basis) f | |||||
| Crude Protein | 54.7 | 54.1 | 53.0 | 52.2 | 52.4 |
| Crude fat | 11.4 | 11.0 | 9.4 | 10.0 | 8.4 |
| Ash | 13.6 | 10.6 | 8.5 | 5.8 | 6.2 |
| Indispensable amino acids (% total amino acids) g | |||||
| Arg | 6.2 | 6.0 | 5.9 | 5.7 | 5.7 |
| His | 3.2 | 3.2 | 3.2 | 3.2 | 3.2 |
| Ile | 4.3 | 4.4 | 4.5 | 4.6 | 4.6 |
| Leu | 9.0 | 8.6 | 8.2 | 7.8 | 7.8 |
| Lys | 7.5 | 6.9 | 6.5 | 5.9 | 5.9 |
| Met + Cys | 4.6 | 3.7 | 3.2 | 2.3 | 2.3 |
| Phe | 4.5 | 4.3 | 4.1 | 3.9 | 3.9 |
| Thr | 4.4 | 4.3 | 4.3 | 4.2 | 4.2 |
| Trp + Tyr | 2.2 | 3.1 | 3.6 | 4.4 | 4.4 |
| Val | 5.1 | 5.7 | 6.0 | 6.6 | 6.6 |
| Ingredients (%) | Control a | 5% MW b | 10% MW c |
|---|---|---|---|
| Fish meal | 50.00 | 45.00 | 40.00 |
| DMW-2 | 0.00 | 5.00 | 10.00 |
| Fish oil | 6.00 | 6.25 | 6.50 |
| Starch | 6.00 | 6.00 | 6.00 |
| Wheat meal | 11.20 | 10.85 | 10.50 |
| Soybean meal | 16.00 | 16.00 | 16.00 |
| Corn gluten meal | 8.00 | 8.00 | 8.00 |
| Taurine | 0.00 | 0.10 | 0.20 |
| Vitamin mix | 0.40 | 0.40 | 0.40 |
| Mineral mix | 0.30 | 0.30 | 0.30 |
| Choline chloride | 0.05 | 0.05 | 0.05 |
| Vitamin C derivatives | 0.05 | 0.05 | 0.05 |
| KH2PO4 | 1.00 | 1.00 | 1.00 |
| Carboxymethyl cellulose | 1.00 | 1.00 | 1.00 |
| Total | 100.00 | 100.00 | 100.00 |
| Proximate Compositions and Amino Acid Compositions | DMW-1 | DMW-2 | FM |
|---|---|---|---|
| Proximate composition (% on a dry matter basis) a | |||
| Crude protein | 75.3 | 76.5 | 71.3 |
| Crude fat | 5.6 | 5.3 | 9.8 |
| Ash | 5.1 | 11.7 | 16.4 |
| Amino acids (% total amino acids) | |||
| Ala | 7.4 | 5.8 | 7.0 |
| Arg | 5.7 | 7.0 | 6.5 |
| Asp | 8.7 | 8.3 | 9.5 |
| Cys | 1.0 | 3.2 | 1.0 |
| Glu | 12.8 | 13.4 | 13.2 |
| Gly | 5.6 | 6.8 | 7.7 |
| His | 3.2 | 2.1 | 3.3 |
| Ile | 4.6 | 3.8 | 4.3 |
| Leu | 7.8 | 8.5 | 8.2 |
| Lys | 5.9 | 7.0 | 8.3 |
| Met | 1.3 | 1.6 | 3.1 |
| Phe | 3.9 | 4.4 | 4.3 |
| Pro | 7.4 | 6.2 | 5.0 |
| Ser | 4.9 | 7.0 | 4.3 |
| Thr | 4.2 | 4.5 | 4.6 |
| Trp | 1.4 | 0.8 | 1.3 |
| Tyr | 7.6 | 3.5 | 3.3 |
| Val | 6.6 | 6.1 | 5.2 |
| Taurine (mg/g dry matter) | 0.1 | 0.1 | 1.5 |
| Components (% Fatty Acids) | Dried Mealworm Larvae a | Fish Oil |
|---|---|---|
| Saturated fatty acid | ||
| 12:0 | 0.3 | 4.1 |
| 14:0 | 2.8 | - |
| 15:0 | 0.2 | 0.5 |
| 16:0 | 15.5 | 15.6 |
| 17:0 | 0.2 | 0.7 |
| 18:0 | 2.4 | 3.6 |
| 20:0 | - | 0.3 |
| 22:0 | - | - |
| Total | 21.4 | 24.8 |
| Monounsaturated fatty acid | ||
| 14:1 | - | - |
| 16:1 | 1.9 | 5.0 |
| 17:1 | 0.1 | 0.5 |
| 18:1 | 35.7 | 19.4 |
| 20:1 | - | 5.2 |
| 22:1 | - | 5.0 |
| 24:1 | - | 0.6 |
| Total | 37.7 | 35.7 |
| Polyunsaturated fatty acid | ||
| ω-3 fatty acid | ||
| 18:3n−3 | 1.7 | 1.0 |
| 20:3n−3 | - | 0.2 |
| 20:4n−3 | - | 0.7 |
| 20:5n−3 | - | 7.5 |
| 21:5n−3 | - | 0.3 |
| 22:5n−3 | - | 1.9 |
| 22:6n−3 | - | 15.7 |
| Total | 1.7 | 27.3 |
| ω-6 fatty acid | ||
| 18:2n−6 | 35.7 | 2.8 |
| 20:2n−6 | - | 0.3 |
| 20:3n−6 | - | 0.2 |
| 20:4n−6 | - | 1.1 |
| 22:5n−6 | - | 0.6 |
| Total | 35.7 | 5.0 |
| Others | ||
| 16:2 | - | 0.3 |
| 16:3 | - | 0.2 |
| 16:4 | - | 0.3 |
| Total | - | 0.8 |
| Not identified | 1.6 | 4.3 |
| Parameters | 65% FM | 25% MW | 40% MW | 65% MW | 65% MW+ MO |
|---|---|---|---|---|---|
| n | 29 | 31 | 32 | 32 | 30 |
| Initial FL (cm) | 10.3 (10.1, 10.5) a | 10.3 (10.1, 10.4) a | 10.4 (10.2, 10.6) a | 10.4 (10.2, 10.6) a | 10.2 (10.0, 10.4) a |
| FL gain (cm) | 1.1 (0.9, 1.2) a | 1.5 (1.4, 1.7) b | 1.5 (1.4, 1.6) b | 1.6 (1.5, 1.6) b | 1.6 (1.4, 1.7) b |
| Initial BW (g) | 24.3 (22.8, 25.8) a | 24.4 (23.2, 25.6) a | 25.9 (24.5, 27.3) a | 25.4 (24.0, 26.7) a | 24.5 (23.0, 26.0) a |
| BW gain (g) | 7.2 (5.6, 8.9) a | 10.4 (8.9, 11.9) b | 11.8 (10.7, 13.0) b, c | 12.8 (12.0, 13.5) c | 10.4 (9.5, 11.2) a, b |
| FL gain rate (%) | 10.7 (9.2, 12.2) a | 14.7 (13.2, 16.2) b | 14.8 (13.6, 15.9) b | 15.1 (14.3, 15.9) b | 15.2 (14.1, 16.4) b |
| BW gain rate (%) | 26.9 (20.9, 32.8) a | 37.9 (32.7, 43.2) b, c | 39.2 (35.8, 42.5) b, c | 42.3 (40.2, 44.3) c | 36.6 (32.7, 40.6) a, b |
| SGR (% day-1) | 0.89 (0.69, 1.10) a | 1.27 (1.08, 1.46) b, c | 1.34 (1.23, 1.46) b, c | 1.46 (1.39, 1.54) c | 1.27 (1.16, 1.36) a, b |
| Feed intake (g) | 8.5 | 11.2 | 13.8 | 14.6 | 12.2 |
| FCR | 1.17 | 1.08 | 1.17 | 1.14 | 1.18 |
| Survival rate (%) | 90.6 | 96.9 | 100.0 | 100.0 | 93.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ido, A.; Hashizume, A.; Ohta, T.; Takahashi, T.; Miura, C.; Miura, T. Replacement of Fish Meal by Defatted Yellow Mealworm (Tenebrio molitor) Larvae in Diet Improves Growth Performance and Disease Resistance in Red Seabream (Pargus major). Animals 2019, 9, 100. https://doi.org/10.3390/ani9030100
Ido A, Hashizume A, Ohta T, Takahashi T, Miura C, Miura T. Replacement of Fish Meal by Defatted Yellow Mealworm (Tenebrio molitor) Larvae in Diet Improves Growth Performance and Disease Resistance in Red Seabream (Pargus major). Animals. 2019; 9(3):100. https://doi.org/10.3390/ani9030100
Chicago/Turabian StyleIdo, Atsushi, Atsushi Hashizume, Takashi Ohta, Takayuki Takahashi, Chiemi Miura, and Takeshi Miura. 2019. "Replacement of Fish Meal by Defatted Yellow Mealworm (Tenebrio molitor) Larvae in Diet Improves Growth Performance and Disease Resistance in Red Seabream (Pargus major)" Animals 9, no. 3: 100. https://doi.org/10.3390/ani9030100
APA StyleIdo, A., Hashizume, A., Ohta, T., Takahashi, T., Miura, C., & Miura, T. (2019). Replacement of Fish Meal by Defatted Yellow Mealworm (Tenebrio molitor) Larvae in Diet Improves Growth Performance and Disease Resistance in Red Seabream (Pargus major). Animals, 9(3), 100. https://doi.org/10.3390/ani9030100