Corticosterone Injection Impairs Follicular Development, Ovulation and Steroidogenesis Capacity in Mice Ovary

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Animals
2.3. Calculation of Ovulation Number
2.4. Follicle Qualitative Standards
2.5. Radioimmunoassay
2.6. Real-Time Quantitative Polymerase Chain Reaction (qRT-PCR)
2.7. Statistical Analysis
3. Results
3.1. Effect of Corticosterone (CORT) Injection on Body Weight and Ovarian Development in Mice
3.2. Injection of CORT Inhibits Ovulation in Mice
3.3. Effects of CORT Injection on Mouse Follicles
3.4. Effect of CORT Injection on Steroidogenesis in Mice
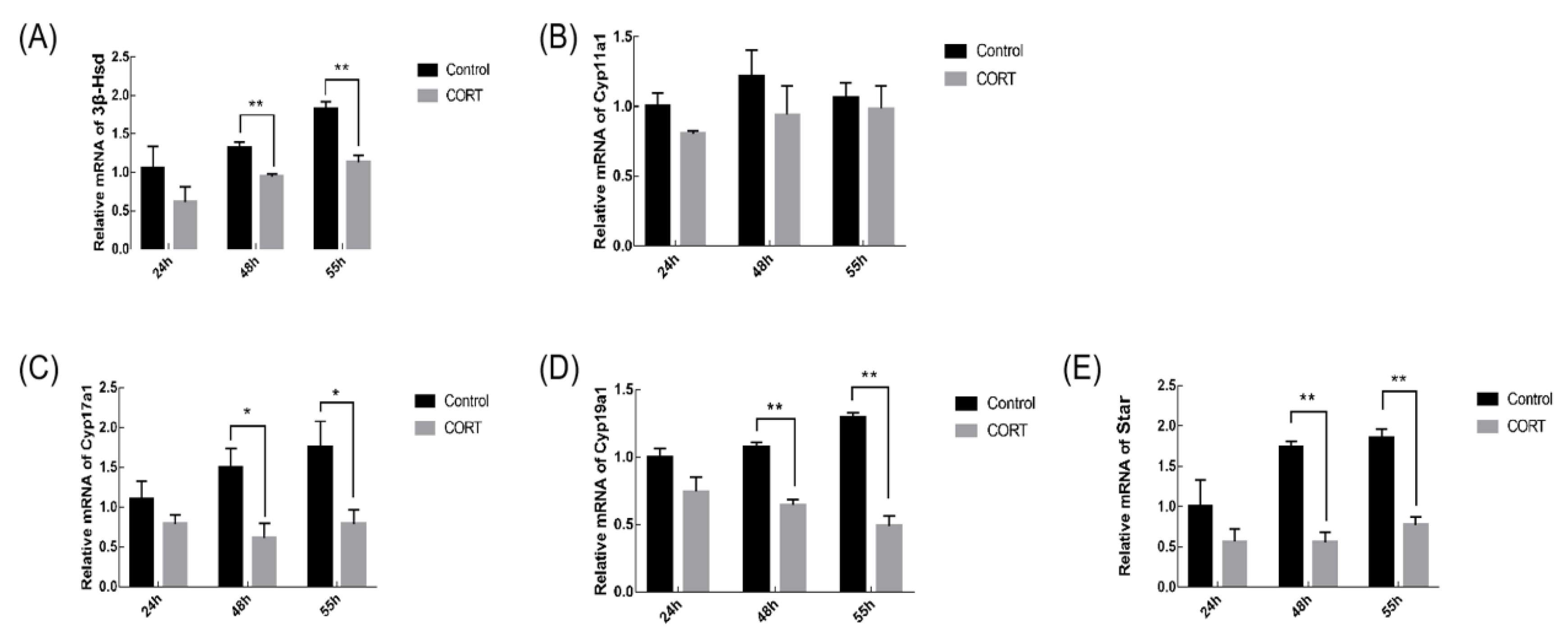
3.5. Effects of CORT Injection on the Transcription Level of Steroid Synthase Gene in Mouse Granulosa Cells (GCs)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eiland, L.; McEwen, B.S. Early life stress followed by subsequent adult chronic stress potentiates anxiety and blunts hippocampal structural remodeling. Hippocampus 2012, 22, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Valsamakis, G.; Chrousos, G.; Mastorakos, G. Stress, female reproduction and pregnancy. Psychoneuroendocrinology 2019, 100, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [PubMed]
- Carroll, J.A.; Burdick, N.C.; Chase, C.C., Jr.; Coleman, S.W.; Spiers, D.E. Influence of environmental temperature on the physiological, endocrine, and immune responses in livestock exposed to a provocative immune challenge. Domest. Anim. Endocrinol. 2012, 43, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Breen, K.M.; Karsch, F.J. New insights regarding glucocorticoids, stress and gonadotropin suppression. Front. Neuroendocrinol. 2006, 27, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Mormede, P.; Andanson, S.; Auperin, B.; Beerda, B.; Guemene, D.; Malmkvist, J.; Manteca, X.; Manteuffel, G.; Prunet, P.; Van Reenen, C.G.; et al. Exploration of the hypothalamic-pituitary-adrenal function as a tool to evaluate animal welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef]
- Dickmeis, T.; Weger, B.D.; Weger, M. The circadian clock and glucocorticoids—Interactions across many time scales. Mol. Cell. Endocrinol. 2013, 380, 2–15. [Google Scholar] [CrossRef]
- McEwen, B.S. Physiology and neurobiology of stress and adaptation: Central role of the brain. Endocrinol. Rev. 2007, 87, 873–904. [Google Scholar] [CrossRef]
- Oster, H.; Challet, E.; Ott, V.; Arvat, E.; De Kloet, E.R.; Dijk, D.J.; Lightman, S.; Vgontzas, A.; Van Cauter, E. The Functional and Clinical Significance of the 24-Hour Rhythm of Circulating Glucocorticoids. Endocr. Rev. 2017, 38, 3–45. [Google Scholar]
- Tilbrook, A.J.; Turner, A.I.; Clarke, I.J. Stress and reproduction: Central mechanisms and sex differences in non-rodent species. Stress 2002, 5, 83–100. [Google Scholar] [CrossRef]
- Turner, A.I.; Tilbrook, A.J. Stress, cortisol and reproduction in female pigs. Soc. Reprod. Fertil. Suppl. 2006, 62, 191–203. [Google Scholar] [PubMed]
- Joels, M. Corticosteroids and the brain. J. Endocrinol. 2018, 238, R121–R130. [Google Scholar] [CrossRef] [PubMed]
- Moloney, R.D.; Desbonnet, L.; Clarke, G.; Dinan, T.G.; Cryan, J.F. The microbiome: Stress, health and disease. Mamm. Genome. 2014, 25, 49–74. [Google Scholar] [CrossRef] [PubMed]
- Daley, C.A.; Sakurai, H.; Adams, B.M.; Adams, T.E. Effect of stress-like concentrations of cortisol on the feedback potency of oestradiol in orchidectomized sheep. Anim. Reprod. Sci. 2000, 59, 167–178. [Google Scholar] [CrossRef]
- Edwards, K.L.; Edes, A.N.; Brown, J.L. Stress, Well-Being and Reproductive Success. Adv. Exp. Med. Biol. 2019, 1200, 91–162. [Google Scholar]
- Kalantaridou, S.N.; Zoumakis, E.; Makrigiannakis, A.; Lavasidis, L.G.; Vrekoussis, T.; Chrousos, G.P. Corticotropin-releasing hormone, stress and human reproduction: An update. J. Reprod. Immunol. 2010, 85, 33–39. [Google Scholar] [CrossRef]
- Dobson, H.; Ghuman, S.; Prabhakar, S.; Smith, R. A conceptual model of the influence of stress on female reproduction. Reproduction 2003, 125, 151–163. [Google Scholar] [CrossRef]
- Ciechanowska, M.; Lapot, M.; Antkowiak, B.; Mateusiak, K.; Paruszewska, E.; Malewski, T.; Paluch, M.; Przekop, F. Effect of short-term and prolonged stress on the biosynthesis of gonadotropin-releasing hormone (GnRH) and GnRH receptor (GnRHR) in the hypothalamus and GnRHR in the pituitary of ewes during various physiological states. Anim. Reprod. Sci. 2016, 174, 65–72. [Google Scholar] [CrossRef]
- Lee, H.Y.; Acosta, T.J.; Tanikawa, M.; Sakumoto, R.; Komiyama, J.; Tasaki, Y.; Piskula, M.; Skarzynski, D.J.; Tetsuka, M.; Okuda, K. The role of glucocorticoid in the regulation of prostaglandin biosynthesis in non-pregnant bovine endometrium. J. Endocrinol. 2007, 193, 127–135. [Google Scholar] [CrossRef]
- Vitellius, G.; Trabado, S.; Bouligand, J.; Delemer, B.; Lombes, M. Pathophysiology of Glucocorticoid Signaling. Ann. Endocrinol. (Paris) 2018, 79, 98–106. [Google Scholar] [CrossRef]
- Chen, Y.; Arsenault, R.; Napper, S.; Griebel, P. Models and Methods to Investigate Acute Stress Responses in Cattle. Animals 2015, 5, 1268–1295. [Google Scholar] [CrossRef] [PubMed]
- Dobson, H.; Smith, R.F. What is stress, and how does it affect reproduction? Anim. Reprod. Sci. 2000, 60–61, 743–752. [Google Scholar] [CrossRef]
- Turner, A.I.; Hemsworth, P.H.; Canny, B.J.; Tilbrook, A.J. Sustained but not repeated acute elevation of cortisol impaired the luteinizing hormone surge, estrus, and ovulation in gilts. Biol. Reprod. 1999, 61, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.M.; Liu, Y.S.; Tong, X.H.; Shen, N.; Jin, R.T.; Han, H.; Hu, M.H.; Wang, W.; Zhou, G.X. Inhibition of follicular development induced by chronic unpredictable stress is associated with growth and differentiation factor 9 and gonadotropin in mice. Biol. Reprod. 2012, 86, 121. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Lin, F.; Zhang, J.; Tang, Y.; Chen, W.K.; Liu, H. Involvement of the up-regulated FoxO1 expression in follicular granulosa cell apoptosis induced by oxidative stress. J. Biol. Chem. 2012, 287, 25727–25740. [Google Scholar] [CrossRef]
- Shen, M.; Liu, Z.; Li, B.; Teng, Y.; Zhang, J.; Tang, Y.; Sun, S.C.; Liu, H. Involvement of FoxO1 in the effects of follicle-stimulating hormone on inhibition of apoptosis in mouse granulosa cells. Cell. Death Dis. 2014, 5, E1475. [Google Scholar] [CrossRef]
- Shen, M.; Jiang, Y.; Guan, Z.; Cao, Y.; Sun, S.C.; Liu, H. FSH protects mouse granulosa cells from oxidative damage by repressing mitophagy. Sci. Rep. 2016, 6, 38090. [Google Scholar] [CrossRef]
- Shen, M.; Jiang, Y.; Guan, Z.; Cao, Y.; Li, L.; Liu, H.; Sun, S.C. Protective mechanism of FSH against oxidative damage in mouse ovarian granulosa cells by repressing autophagy. Autophagy 2017, 13, 1364–1385. [Google Scholar] [CrossRef]
- Cooke, R.F.; Schubach, K.M.; Marques, R.S.; Peres, R.F.; Silva, L.G.; Carvalho, R.S.; Cipriano, R.S.; Bohnert, D.W.; Pires, A.V.; Vasconcelos, J.L. Effects of temperament on physiological, productive, and reproductive responses in beef cows. J. Anim. Sci. 2017, 95, 1–8. [Google Scholar] [CrossRef]
- Ennab, W.; Mustafa, S.; Wei, Q.; Lv, Z.; Kavita, N.M.X.; Ullah, S.; Shi, F. Resveratrol Protects against Restraint Stress Effects on Stomach and Spleen in Adult Male Mice. Animals 2019, 9, 736. [Google Scholar] [CrossRef]
- Papp, M.; Gruca, P.; Lason-Tyburkiewicz, M.; Adham, N.; Kiss, B.; Gyertyan, I. Attenuation of anhedonia by cariprazine in the chronic mild stress model of depression. Behav. Pharmacol. 2014, 25, 567–574. [Google Scholar] [PubMed]
- Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and usage. Neurobiol. Stress 2017, 6, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Antoniuk, S.; Bijata, M.; Ponimaskin, E.; Wlodarczyk, J. Chronic unpredictable mild stress for modeling depression in rodents: Meta-analysis of model reliability. Neurosci. Biobehav. Rev. 2019, 99, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. Validity, reliability and utility of the chronic mild stress model of depression: A 10-year review and evaluation. Psychopharmacology (Berl.) 1997, 134, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.F.; Sun, X.F.; Han, Z.L.; Li, L.; Ge, W.; Zhao, Y.; De Felici, M.; Shen, W.; Cheng, S.F. Protective effects of melatonin against nicotine-induced disorder of mouse early folliculogenesis. Aging (Albany NY) 2018, 10, 463–480. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.X.; Zhang, Y.; Li, Y.Y.; Liu, X.M.; Wang, X.X.; Zhang, C.L.; Hao, C.F.; Deng, S.L. Regulation of follicular development and differentiation by intra-ovarian factors and endocrine hormones. Front. Biosci. (Landmark Ed.) 2019, 24, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, A.; Kafi, M.; Zamiri, M.J. Factors affecting the size of ovulatory follicles and conception rate in high-yielding dairy cows. Theriogenology 2016, 85, 747–753. [Google Scholar] [CrossRef]
- McGee, E.A.; Hsueh, A.J. Initial and cyclic recruitment of ovarian follicles. Endocr. Rev. 2000, 21, 200–214. [Google Scholar] [CrossRef]
- Carreau, S.; Bourguiba, S.; Lambard, S.; Galeraud-Denis, I.; Genissel, C.; Levallet, J. Reproductive system: Aromatase and estrogens. Mol. Cell Endocrinol. 2002, 193, 137–143. [Google Scholar] [CrossRef]
- Miller, W.L. Role of mitochondria in steroidogenesis. Endocr. Dev. 2011, 20, 1–19. [Google Scholar]
- Rasmussen, M.K.; Ekstrand, B.; Zamaratskaia, G. Regulation of 3beta-hydroxysteroid dehydrogenase/Delta(5)-Delta(4) isomerase: A review. Int. J. Mol. Sci. 2013, 14, 17926–17942. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.S.; Qian, Z.; Liu, H.L.; Bao, E.D. ACTH-induced stress in weaned sows impairs LH receptor expression and steroidogenesis capacity in the ovary. Reprod. Biol. Endocrinol. 2016, 14, 80. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence | Reverse Sequence |
|---|---|---|
| 3β-Hsd | 5′-GCTGCACAGCCCTCCTAAG-3′ | 5′-TGATCCTCTGGCCCACAAAC-3′ |
| Cyp11a1 | 5′-CCCGGAGAGCTTGTGCAAAT-3′ | 5′-CCCATGCTGAGCCAGATGTC-3′ |
| Cyp17a1 | 5′-TGGAGGCCACTATCCGAGAA-3′ | 5′-GAAGCGCTCAGGCATAAACC-3′ |
| Cyp19a1 | 5′-ATCCGGTTTTTAAACGGCTGC-3′ | 5′-TCTTGCGCTATTTGGCCTGG-3′ |
| Star | 5′-AACGGGGACGAAGTGCTAAG-3′ | 5′-CCTCTGCAGGACCTTGATCTC-3′ |
| Gapdh | 5′-AAGGTGGTGAAGCAGGCAT-3′ | 5′-GGTCCAGGGTTTCTTACTCCT -3′ |
| Time (h) | Groups | Estradiol (pg/mL) | Progesterone (ng/mL) |
|---|---|---|---|
| 24 | Control group | 51.30 ± 9.62 | 44.97 ± 3.86 |
| CORT treatment group | 33.53 ± 16.46 | 26.50 ± 3.49 * | |
| 48 | Control group | 80.91 ± 7.80 | 31.23 ± 3.08 |
| CORT treatment group | 57.10 ± 14.57 | 21.79 ± 1.96 * | |
| 55 | Control group | 81.70 ± 18.47 | 37.63 ± 1.60 |
| CORT treatment group | 31.42 ± 5.63 ** | 15.33 ± 2.30 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Y.; Li, W.; Meng, X.; Zhang, L.; Shen, M.; Liu, H. Corticosterone Injection Impairs Follicular Development, Ovulation and Steroidogenesis Capacity in Mice Ovary. Animals 2019, 9, 1047. https://doi.org/10.3390/ani9121047
Wei Y, Li W, Meng X, Zhang L, Shen M, Liu H. Corticosterone Injection Impairs Follicular Development, Ovulation and Steroidogenesis Capacity in Mice Ovary. Animals. 2019; 9(12):1047. https://doi.org/10.3390/ani9121047
Chicago/Turabian StyleWei, Yinghui, Weijian Li, Xueqing Meng, Liangliang Zhang, Ming Shen, and Honglin Liu. 2019. "Corticosterone Injection Impairs Follicular Development, Ovulation and Steroidogenesis Capacity in Mice Ovary" Animals 9, no. 12: 1047. https://doi.org/10.3390/ani9121047
APA StyleWei, Y., Li, W., Meng, X., Zhang, L., Shen, M., & Liu, H. (2019). Corticosterone Injection Impairs Follicular Development, Ovulation and Steroidogenesis Capacity in Mice Ovary. Animals, 9(12), 1047. https://doi.org/10.3390/ani9121047