Variants in BMP7 and BMP15 3’-UTRs Associated with Reproductive Traits in a Large White Pig Population


Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Genomic DNA Extraction
2.3. Primer Design
2.4. PCR Amplification and Sequencing
2.5. Sequence Analysis
2.6. Genotyping
2.7. Association Analysis
3. Results
3.1. Isolation and Characterization of the 3’-UTR of the Porcine BMP7 Gene
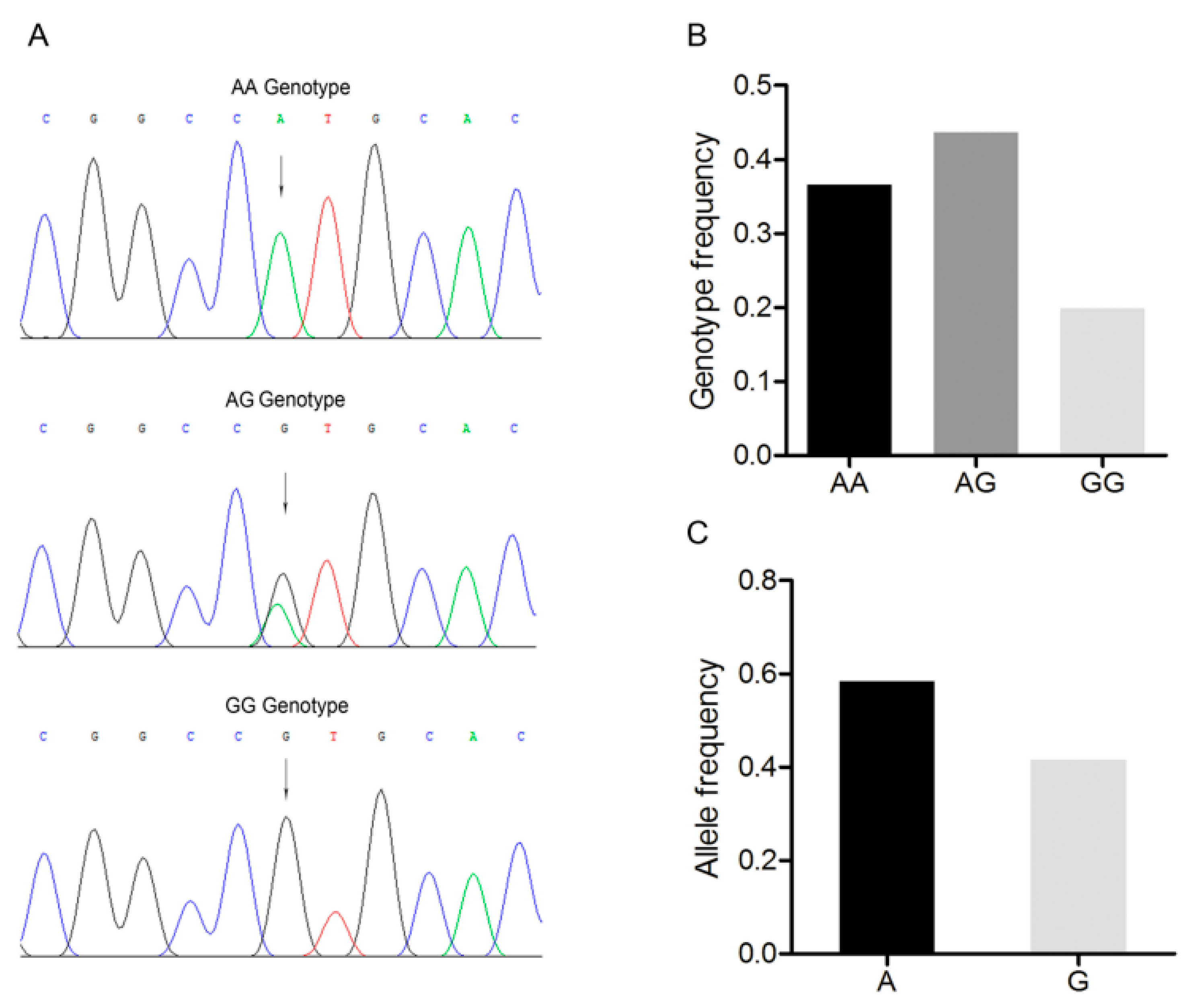
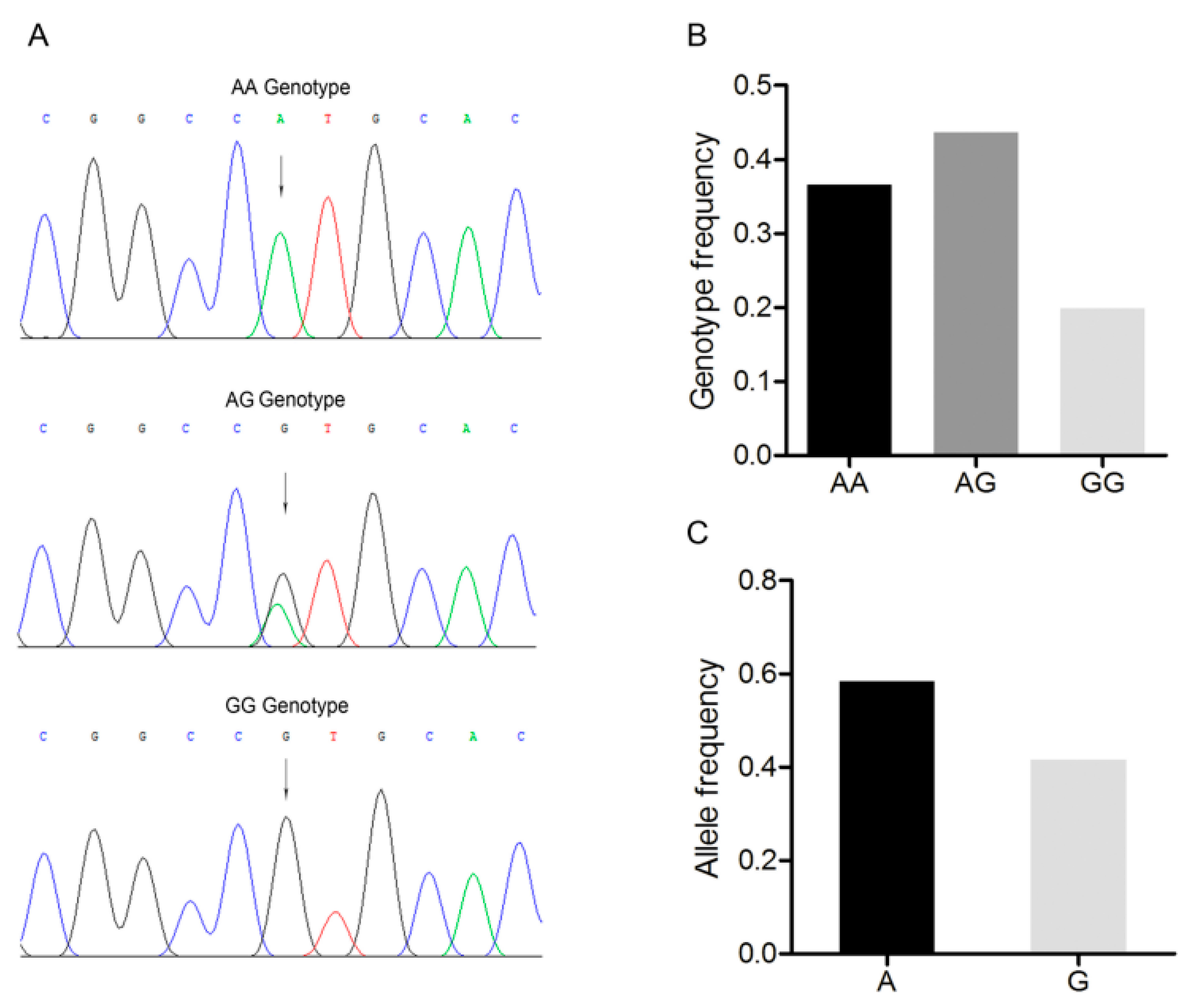
3.2. Polymorphism of the 3’-UTR of the Porcine BMP7 Gene
3.3. Association Analysis between the BMP7 c.1569A>G Polymorphism and Reproductive Traits
3.4. Isolation and Characterization of the 3’-UTR of the Porcine BMP15 Gene
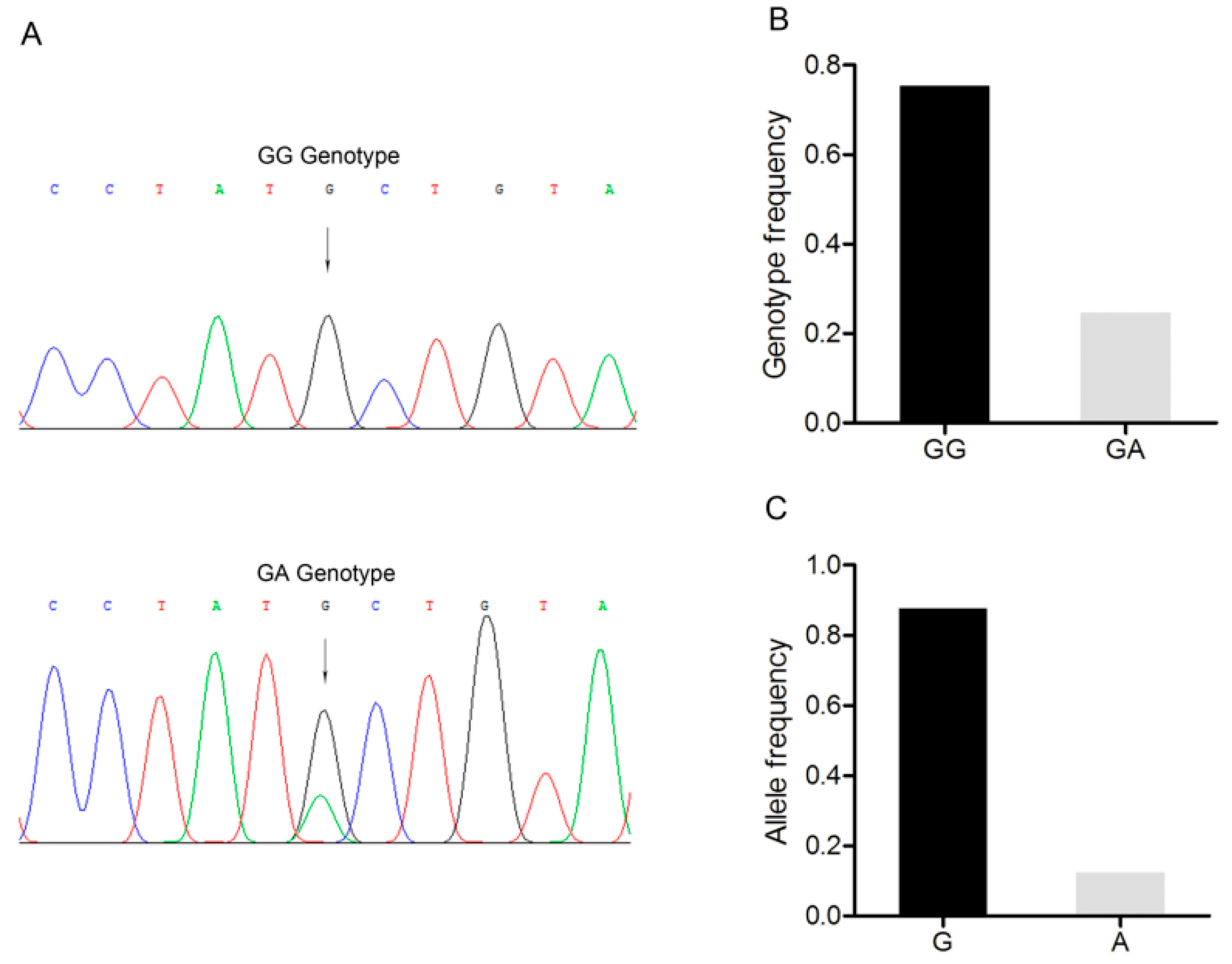
3.5. Detection of a G/A Point Mutation Site in the 3’-UTR of the BMP15 Gene
3.6. Association Analysis between the BMP15 c.2366G>A Polymorphism and Reproductive Traits
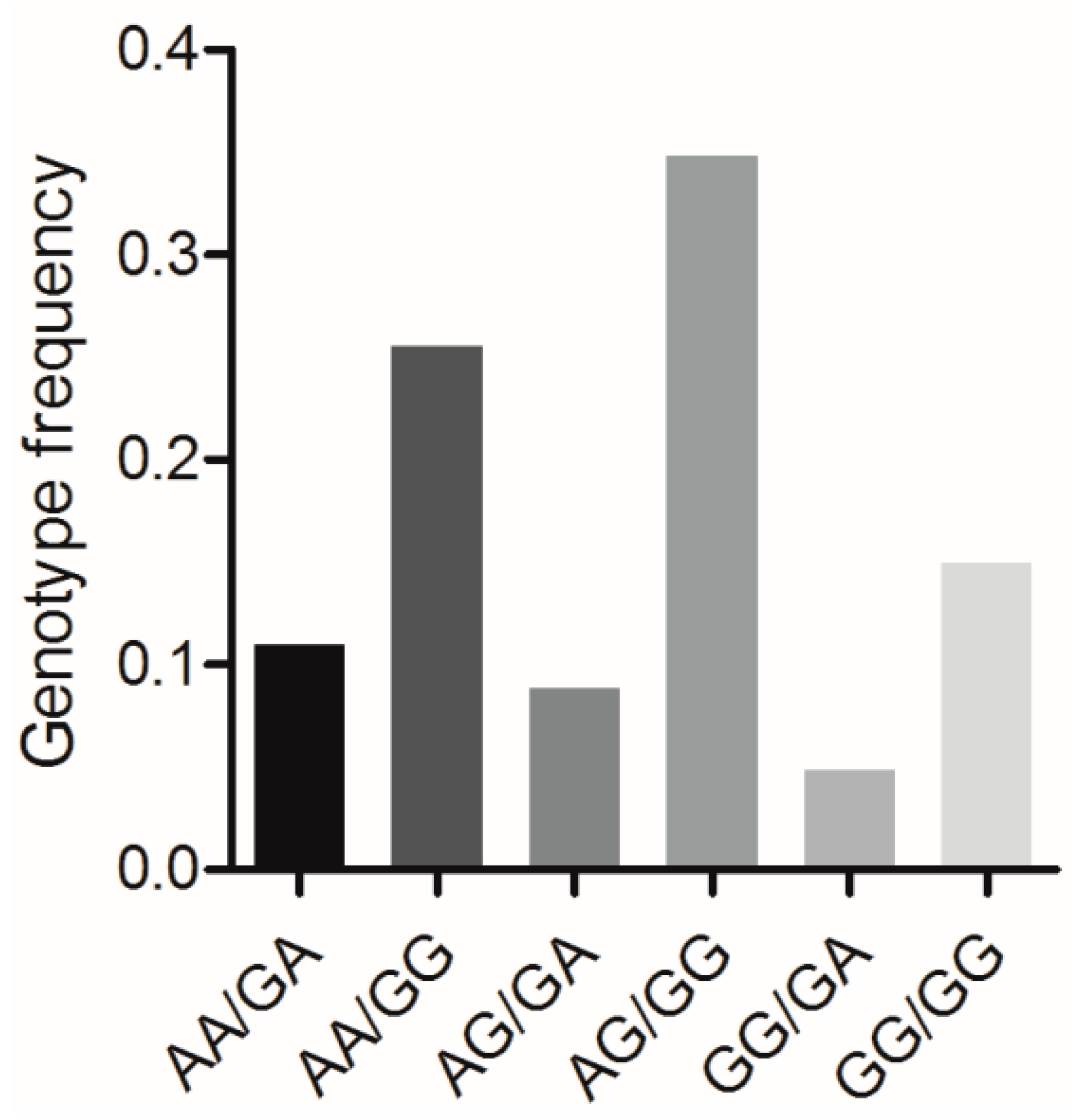
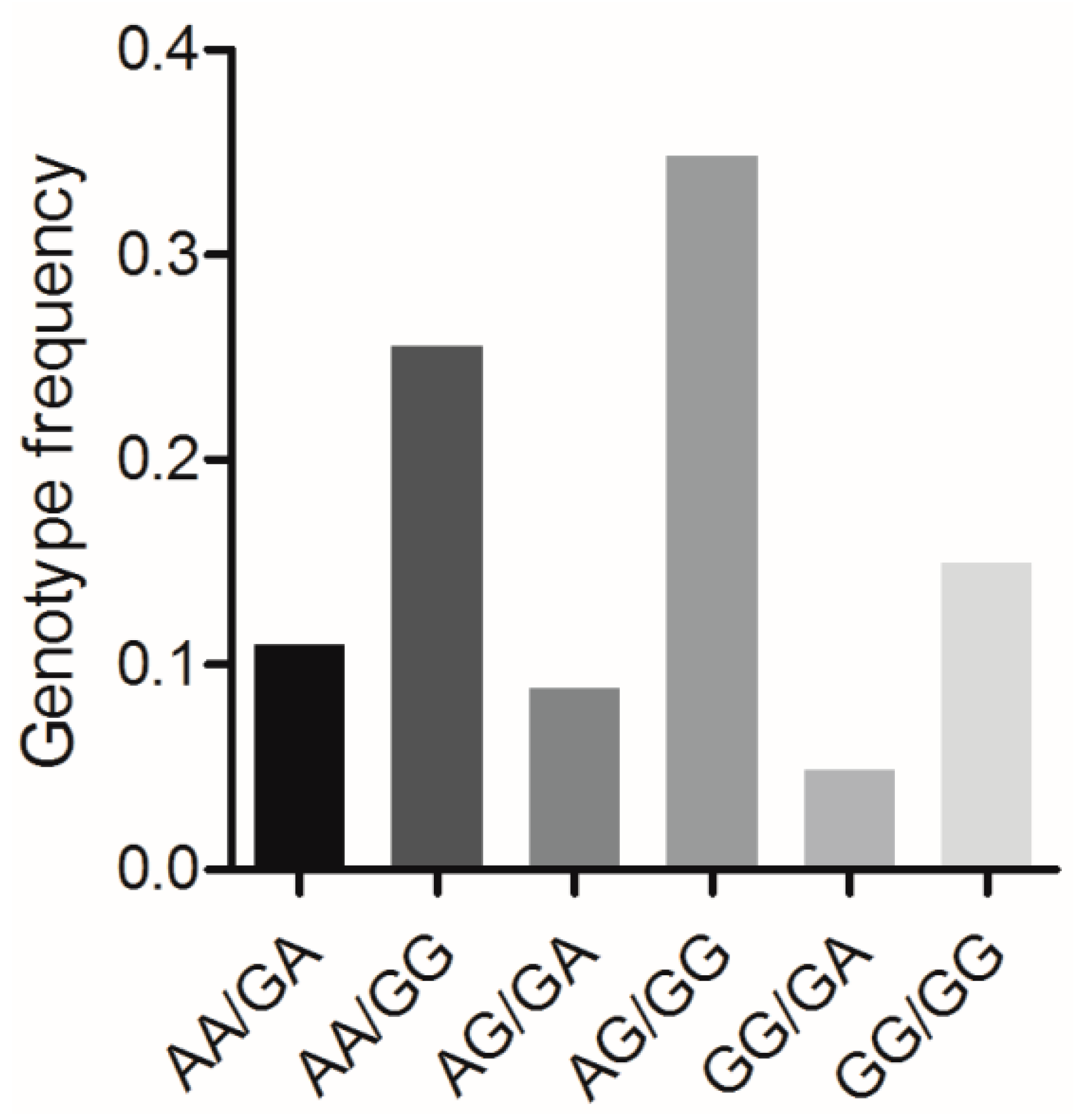
3.7. Association Analysis between the Combined Genotypes of BMP7 c.1569A>G and BMP15 c.2366G>A and Reproductive Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ongaro, L.; Schang, G.; Ho, C.C.; Zhou, X.; Bernard, D.J. TGF-beta superfamily regulation of follicle-stimulating hormone synthesis by gonadotrope cells: Is there a role for bone morphogenetic proteins? Endocrinology 2019, 3, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Abdurahman, A.; Du, X.; Yao, Y.; Sulaiman, Y.; Aniwashi, J.; Li, Q. Smad4 feedback enhances BMPR1B transcription in ovine granulosa cells. Int. J. Mol. Sci. 2019, 20, 2732. [Google Scholar] [CrossRef] [PubMed]
- Richani, D.; Gilchrist, R.B. The epidermal growth factor network: Role in oocyte growth, maturation and developmental competence. Hum. Reprod. Update 2018, 1, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fortin, J.; Lamba, P.; Bonomi, M.; Persani, L.; Roberson, M.S.; Bernard, D.J. Activator protein-1 and smad proteins synergistically regulate human follicle-stimulating hormone beta-promoter activity. Endocrinology 2008, 11, 5577–5591. [Google Scholar]
- Du, X.; Zhang, L.; Li, X.; Pan, Z.; Liu, H.; Li, Q. TGF-beta signaling controls FSHR signaling-reduced ovarian granulosa cell apoptosis through the SMAD4/miR-143 axis. Cell Death Dis. 2016, 11, e2476. [Google Scholar]
- Li, Q.; Du, X.; Pan, Z.; Zhang, L.; Li, Q. The transcription factor SMAD4 and miR-10b contribute to E2 release and cell apoptosis in ovarian granulosa cells by targeting CYP19A1. Mol. Cell Endocrinol. 2018, 476, 84–95. [Google Scholar] [CrossRef]
- Yan, C.; Wang, P.; DeMayo, J.; DeMayo, F.J.; Elvin, J.A.; Carino, C.; Prasad, S.V.; Skinner, S.S.; Dunbar, B.S.; Dube, J.L.; et al. Synergistic roles of bone morphogenetic protein 15 and growth differentiation factor 9 in ovarian function. Mol. Endocrinol. 2001, 6, 854–866. [Google Scholar] [CrossRef]
- Sanfins, A.; Rodrigues, P.; Albertini, D.F. GDF-9 and BMP-15 direct the follicle symphony. J. Assist. Reprod. Genet. 2018, 10, 1741–1750. [Google Scholar]
- Galloway, S.M.; McNatty, K.P.; Cambridge, L.M.; Laitinen, M.P.; Juengel, J.L.; Jokiranta, T.S.; McLaren, R.J.; Luiro, K.; Dodds, K.G.; Montgomery, G.W.; et al. Mutations in an oocyte-derived growth factor gene (BMP15) cause increased ovulation rate and infertility in a dosage-sensitive manner. Nat. Genet. 2000, 3, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Mulsant, P.; Lecerf, F.; Fabre, S.; Schibler, L.; Monget, P.; Lanneluc, I.; Pisselet, C.; Riquet, J.; Monniaux, D.; Callebaut, I.; et al. Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Merino ewes. Proc. Natl. Acad. Sci. USA 2001, 9, 5104–5109. [Google Scholar] [CrossRef] [PubMed]
- Abdoli, R.; Zamani, P.; Mirhoseini, S.Z.; Ghavi, H.N.; Nadri, S. A review on prolificacy genes in sheep. Reprod. Domest. Anim. 2016, 5, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Li, W.T.; Zhang, M.M.; Li, Q.G.; Tang, H.; Zhang, L.F.; Wang, K.J.; Zhu, M.Z.; Lu, Y.F.; Bao, H.G.; Zhang, Y.M.; et al. Whole-genome resequencing reveals candidate mutations for pig prolificacy. Proc. Biol. Sci. 2017, 248. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ye, J.; Han, X.; Qiao, R.; Li, X.; Lv, G.; Wang, K. Whole-genome sequencing identifies potential candidate genes for reproductive traits in pigs. Genomics 2019. [Google Scholar] [CrossRef] [PubMed]
- Mayr, C. Regulation by 3′-Untranslated Regions. Annu. Rev. Genet. 2017, 17–194. [Google Scholar] [CrossRef]
- Vejnar, C.E.; Abdel, M.M.; Takacs, C.M.; Yartseva, V.; Oikonomou, P.; Christiano, R.; Stoeckius, M.; Lau, S.; Lee, M.T.; Beaudoin, J.D.; et al. Genome wide analysis of 3’ UTR sequence elements and proteins regulating mRNA stability during maternal-to-zygotic transition in zebrafish. Genome Res. 2019, 7, 1100–1114. [Google Scholar] [CrossRef]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 1, 20–51. [Google Scholar] [CrossRef]
- Yang, L.; Du, X.; Liu, L.; Cao, Q.; Pan, Z.; Li, Q. miR-1306 Mediates the Feedback Regulation of the TGF-beta/SMAD Signaling Pathway in Granulosa Cells. Cells 2019, 8, 298. [Google Scholar] [CrossRef]
- Woo, I.; Christenson, L.K.; Gunewardena, S.; Ingles, S.A.; Thomas, S.; Ahmady, A.; Chung, K.; Bendikson, K.; Paulson, R.; McGinnis, L.K. Micro-RNAs involved in cellular proliferation have altered expression profiles in granulosa of young women with diminished ovarian reserve. J. Assist. Reprod Genet. 2018, 10, 1777–1786. [Google Scholar] [CrossRef]
- Donadeu, F.X.; Mohammed, B.T.; Ioannidis, J. A miRNA target network putatively involved in follicular atresia. Domest. Anim. Endocrinol. 2017, 58, 76–83. [Google Scholar] [CrossRef]
- Mao, Z.; Fan, L.; Yu, Q.; Luo, S.; Wu, X.; Tang, J.; Kang, G.; Tang, L. Abnormality of Klotho Signaling Is Involved in Polycystic Ovary Syndrome. Reprod. Sci. 2018, 3, 372–383. [Google Scholar] [CrossRef]
- Hu, Z.; Shen, W.J.; Kraemer, F.B.; Azhar, S. Regulation of adrenal and ovarian steroidogenesis by miR-132. J. Mol. Endocrinol. 2017, 3, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, Y.; Yuan, Y.; Nie, F.; Peng, R.; Li, Q.; Lyu, Z.; Mao, Z.; Huang, L.; Zhou, L.; et al. MiR542-3p Regulates the Epithelial-Mesenchymal Transition by Directly Targeting BMP7 in NRK52e. Int. J. Mol. Sci. 2015, 11, 27945–27955. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.; Sun, Y.; He, P. MicroRNA-137 inhibits BMP7 to enhance the epithelial-mesenchymal transition of breast cancer cells. Oncotarget 2017, 11, 18348–18358. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Maurino, S.M.; Rivero-Rodriguez, F.; Velazquez-Cruz, A.; Hernandez-Vellisca, M.; Diaz-Quintana, A.; De la Rosa, M.A.; Diaz-Moreno, I. RNA Binding Protein Regulation and Cross-Talk in the Control of AU-rich mRNA Fate. Front. Mol. Biosci. 2017, 71. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, H.; Fukao, A.; Funakami, Y.; Duncan, K.E.; Fujiwara, T. Emerging Evidence of Translational Control by AU-Rich Element-Binding Proteins. Front. Genet. 2019, 332. [Google Scholar] [CrossRef]
- Abdelmohsen, K.; Tominaga, K.; Lee, E.K.; Srikantan, S.; Kang, M.J.; Kim, M.M.; Selimyan, R.; Martindale, J.L.; Yang, X.; Carrier, F.; et al. Enhanced translation by Nucleolin via G-rich elements in coding and non-coding regions of target mRNAs. Nucleic Acids Res. 2011, 19, 8513–8530. [Google Scholar] [CrossRef]
- Lepeta, K.; Purzycka, K.J.; Pachulska-Wieczorek, K.; Mitjans, M.; Begemann, M.; Vafadari, B.; Bijata, K.; Adamiak, R.W.; Ehrenreich, H.; Dziembowska, M.; et al. A normal genetic variation modulates synaptic MMP-9 protein levels and the severity of schizophrenia symptoms. EMBO Mol. Med. 2017, 8, 1100–1116. [Google Scholar] [CrossRef]
- Rajesh, G.; Mishra, S.R.; Paul, A.; Punetha, M.; Vidyalakshmi, G.M.; Narayanan, K.; Bag, S.; Bhure, S.K.; Singh, C.V.; Maurya, V.P.; et al. Transcriptional and translational abundance of Bone morphogenetic protein (BMP) 2, 4, 6, 7 and their receptors BMPR1A, 1B and BMPR2 in buffalo ovarian follicle and the role of BMP4 and BMP7 on estrogen production and survival of cultured granulosa cells. Res. Vet. Sci. 2018, 118, 371–388. [Google Scholar] [CrossRef]
- Glister, C.; Regan, S.L.; Samir, M.; Knight, P. Gremlin, Noggin, Chordin and follistatin differentially modulate BMP induced suppression of androgen secretion by bovine ovarian theca cells. J. Mol. Endocrinol. 2018. [Google Scholar] [CrossRef]
- Zhang, H.; Klausen, C.; Zhu, H.; Chang, H.M.; Leung, P.C. BMP4 and BMP7 Suppress StAR and Progesterone Production via ALK3 and SMAD1/5/8-SMAD4 in Human Granulosa-Lutein Cells. Endocrinology 2015, 11, 4269–4280. [Google Scholar] [CrossRef]
- Rajesh, G.; Paul, A.; Mishra, S.R.; Bharati, J.; Thakur, N.; Mondal, T.; Soren, S.; Harikumar, S.; Narayanan, K.; Chouhan, V.S.; et al. Expression and functional role of Bone Morphogenetic Proteins (BMPs) in cyclical corpus luteum in buffalo (Bubalus bubalis). Gen. Comp. Endocrinol. 2017, 240, 198–213. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.S.; Otsuka, F.; Moore, R.K.; Shimasaki, S. Effect of bone morphogenetic protein-7 on folliculogenesis and ovulation in the rat. Biol. Reprod. 2001, 4, 994–999. [Google Scholar] [CrossRef] [PubMed]
- Bahire, S.V.; Rajput, P.K.; Kumar, V.; Kumar, D.; Kataria, M.; Kumar, S. Quantitative expression of mRNA encoding BMP/SMAD signaling genes in the ovaries of Booroola carrier and non-carrier GMM sheep. Reprod. Domest. Anim. 2019, 54, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, F.; Feng, X.; Yang, H.; Zhu, A.; Pang, J.; Han, L.; Zhang, T.; Yao, X.; Wang, F. Genome-wide analysis of DNA Methylation profiles on sheep ovaries associated with prolificacy using whole-genome Bisulfite sequencing. BMC Genom. 2017, 1, 759. [Google Scholar] [CrossRef]
- Feng, X.; Xie, S.Y.; Zhou, J.S.; Sun, G.R.; Lu, P.; Li, M. Polymorphisms of the bone morphogenetic protein 7 gene (BMP7) and association analysis with sow productive traits. Anim. Reprod. Sci. 2013, 1, 56–62. [Google Scholar] [CrossRef]
- Christoforou, E.R.; Pitman, J.L. Intrafollicular growth differentiation factor 9: Bone morphogenetic 15 ratio determines litter size in mammalsdagger. Biol. Reprod. 2019, 5, 1333–1343. [Google Scholar] [CrossRef]
- Garcia, P.; Aspee, K.; Ramirez, G.; Dettleff, P.; Palomino, J.; Peralta, O.A.; Parraguez, V.H.; De Los, R.M. Influence of growth differentiation factor 9 and bone morphogenetic protein 15 on in vitro maturation of canine oocytes. Reprod. Domest. Anim. 2019, 2, 373–380. [Google Scholar] [CrossRef]
- Velasquez, A.; Mellisho, E.; Castro, F.O.; Rodriguez-Alvarez, L. Effect of BMP15 and/or AMH during in vitro maturation of oocytes from involuntarily culled dairy cows. Mol. Reprod. Dev. 2019, 2, 209–223. [Google Scholar] [CrossRef]
- Hanrahan, J.P.; Gregan, S.M.; Mulsant, P.; Mullen, M.; Davis, G.H.; Powell, R.; Galloway, S.M. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol Reprod. 2004, 4, 900–909. [Google Scholar] [CrossRef]
- Roy, S.; Gandra, D.; Seger, C.; Biswas, A.; Kushnir, V.A.; Gleicher, N.; Kumar, T.R.; Sen, A. Oocyte-Derived Factors (GDF9 and BMP15) and FSH Regulate AMH Expression Via Modulation of H3K27AC in Granulosa Cells. Endocrinology 2018, 9, 3433–3445. [Google Scholar] [CrossRef]
- Tang, J.; Hu, W.; Di, R.; Liu, Q.; Wang, X.; Zhang, X.; Zhang, J.; Chu, M. Expression Analysis of the Prolific Candidate Genes, BMPR1B, BMP15, and GDF9 in Small Tail Han Ewes with Three Fecundity (FecB Gene) Genotypes. Animals 2018, 8, 166. [Google Scholar] [CrossRef] [PubMed]
- Demars, J.; Fabre, S.; Sarry, J.; Rossetti, R.; Gilbert, H.; Persani, L.; Tosser-Klopp, G.; Mulsant, P.; Nowak, Z.; Drobik, W.; et al. Genome-wide association studies identify two novel BMP15 mutations responsible for an atypical hyperprolificacy phenotype in sheep. PLoS Genet. 2013, 4, e1003482. [Google Scholar] [CrossRef] [PubMed]
- Lassoued, N.; Benkhlil, Z.; Woloszyn, F.; Rejeb, A.; Aouina, M.; Rekik, M.; Fabre, S.; Bedhiaf-Romdhani, S. FecX (Bar) a Novel BMP15 mutation responsible for prolificacy and female sterility in Tunisian Barbarine Sheep. BMC Genet. 2017, 1, 43. [Google Scholar] [CrossRef]
- Zak, L.J.; Gaustad, A.H.; Bolarin, A.; Broekhuijse, M.; Walling, G.A.; Knol, E.F. Genetic control of complex traits, with a focus on reproduction in pigs. Mol. Reprod Dev. 2017, 9, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Panasiewicz, G.; Bieniek-Kobuszewska, M.; Lipka, A.; Majewska, M.; Jedryczko, R.; Szafranska, B. Novel effects of identified SNPs within the porcine Pregnancy-Associated Glycoprotein gene family (pPAGs) on the major reproductive traits in Hirschmann hybrid-line sows. Res. Vet. Sci. 2017, 114, 123–130. [Google Scholar] [CrossRef]
- Pang, P.; Li, Z.; Hu, H.; Wang, L.; Sun, H.; Mei, S.; Li, F. Genetic effect and combined genotype effect of ESR, FSHbeta, CTNNAL1 and miR-27a loci on litter size in a Large White population. Anim. Biotechnol. 2018, 30, 287–292. [Google Scholar] [CrossRef]
- Haggman, J.; Uimari, P. Novel harmful recessive haplotypes for reproductive traits in pigs. J. Anim. Breed. Genet. 2017, 2, 129–135. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5’-3’) | Annealing Temp (°C) | Product Size (bp) | Accession No. | |
|---|---|---|---|---|---|
| P1 | BMP7 | F: GTGTTCCAGGTCCACTTCAT R: CCCAACTCCAAGCAGAAA | 54 °C | 822 | XM_005673044.3 |
| P2 | BMP15 | F: GTGCCTATTAGCATCCTCC R: CTAACTGAAGCCTCTACCC | 54 °C | 1063 | NM_001005155.2 |
| P3 | BMP15 | F: TTTGAGGGAAACAGAAGG R: GTGGCTAAAGGGAACAAA | 54 °C | 723 | NM_001005155.2 |
| Genotypes (n) | Traits (LSM ± SE) | |||
|---|---|---|---|---|
| TNB | NBA | NSB | LW | |
| AA (83) | 12.26(±0.53) a | 12.07(±0.52) a | 0.18(±0.15) a | 17.00(±0.70) b |
| AG (99) | 12.10(±0.54) a | 11.87(±0.54) a | 0.23(±0.16) a | 18.03(±0.72) a |
| GG (45) | 11.65(±0.63) a | 11.60(±0.62) a | 0.16(±0.17) a | 17.40(±0.82) ab |
| Genotypes (n) | Traits (LSM ± SE) | |||
|---|---|---|---|---|
| TNB | NBA | NSB | LW | |
| GA(56) | 12.56(±0.44) a | 12.24(±0.42) a | 0.21(±0.15) a | 17.20(±0.72) a |
| GG(171) | 12.63(±0.43) a | 12.34(±0.42) a | 0.20(±0.15) a | 17.57(±0.71) a |
| Genotypes (n) | Traits (LSM ± SE) | |||
|---|---|---|---|---|
| TNB | NBA | NSB | LW | |
| AA/GA(25) | 12.44(±0.60) a | 12.21(±0.59) a | 0.03(±0.20) a | 17.08(±0.95) a |
| AA/GG(58) | 12.02(±0.59) ab | 11.91(±0.58) ab | 0.07(±0.20) a | 17.06(±0.93) a |
| AG/GA(20) | 11.17(±0.67) b | 11.04(±0.66) b | 0.04(±0.22) a | 17.04(±1.09) a |
| AG/GG(79) | 12.31(±0.56) a | 12.08(±0.56) a | 0.14(±0.19) a | 18.25(±0.90) a |
| GG/GA(11) | 11.81(±0.80) ab | 11.78(±0.80) ab | 0.01(±0.26) a | 17.36(±1.32) a |
| GG/GG(34) | 11.49(±0.69) ab | 11.48(±0.68) ab | 0.04(±0.23) a | 16.97(±1.09) a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, H.; Du, X.; Li, Q.; Pan, Z.; Wu, W.; Liu, H.; Li, Q. Variants in BMP7 and BMP15 3’-UTRs Associated with Reproductive Traits in a Large White Pig Population. Animals 2019, 9, 905. https://doi.org/10.3390/ani9110905
Yin H, Du X, Li Q, Pan Z, Wu W, Liu H, Li Q. Variants in BMP7 and BMP15 3’-UTRs Associated with Reproductive Traits in a Large White Pig Population. Animals. 2019; 9(11):905. https://doi.org/10.3390/ani9110905
Chicago/Turabian StyleYin, Hang, Xing Du, Qiqi Li, Zengxiang Pan, Wangjun Wu, Honglin Liu, and Qifa Li. 2019. "Variants in BMP7 and BMP15 3’-UTRs Associated with Reproductive Traits in a Large White Pig Population" Animals 9, no. 11: 905. https://doi.org/10.3390/ani9110905
APA StyleYin, H., Du, X., Li, Q., Pan, Z., Wu, W., Liu, H., & Li, Q. (2019). Variants in BMP7 and BMP15 3’-UTRs Associated with Reproductive Traits in a Large White Pig Population. Animals, 9(11), 905. https://doi.org/10.3390/ani9110905