The Limiting Sequence and Appropriate Amino Acid Ratio of Lysine, Methionine, and Threonine for Seven- to Nine-Month-Old Holstein Heifers Fed Corn–Soybean M-Based Diet


 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Experimental Design
2.2. Sampling and Analyses
2.3. Amino Acid Partial Partial Deletion Method
2.4. Statistical Analyses
3. Results
3.1. Growth Performance
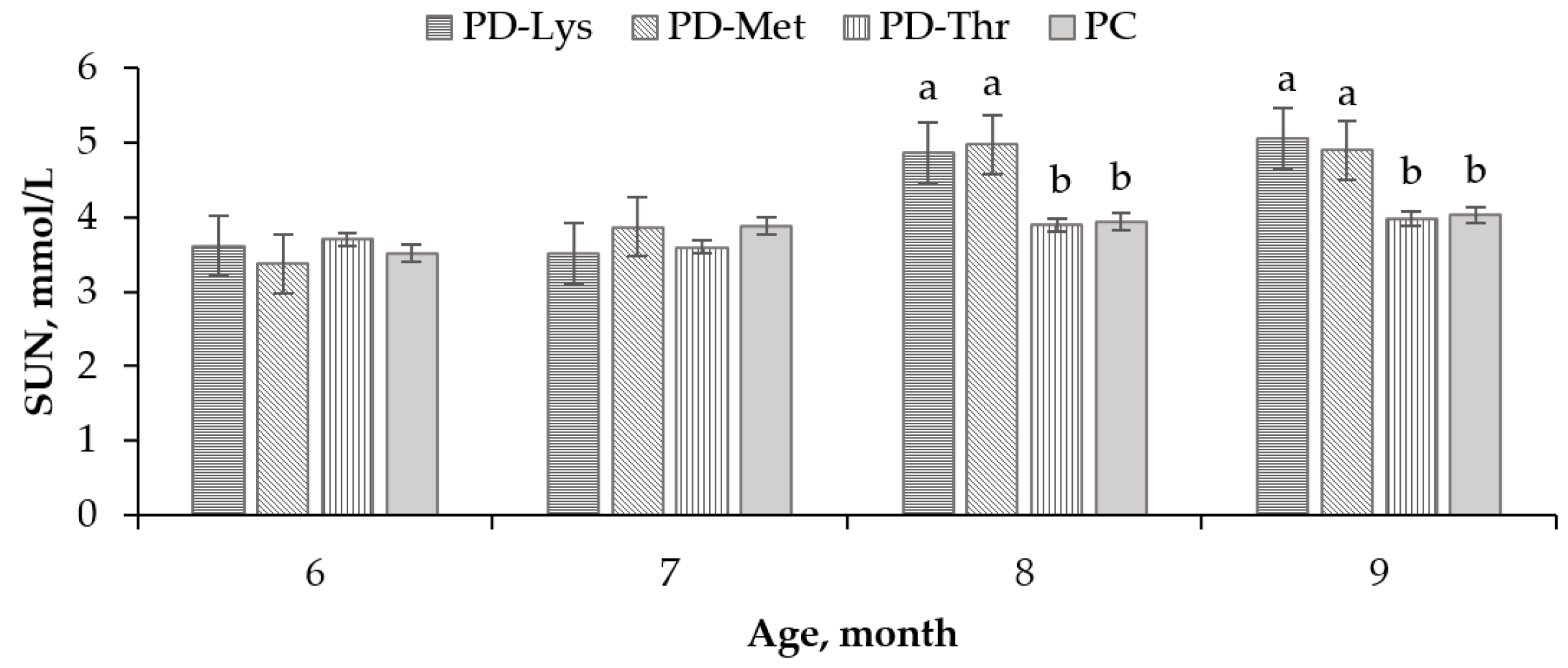
3.2. Serum Urea Nitrogen Levels
3.3. Nitrogen Metabolism
3.4. Appropriate Amino Acid Model
3.4.1. N Retention and Amino Acid Intake
3.4.2. Calculation and Model Diagram of the Effect on Nitrogen Retention
3.4.3. Appropriate Amino Acid Ratio
4. Discussion
4.1. Growth Performance and Body Size
4.2. Serum Urea Nitrogen and Nitrogen Retention
4.3. Limiting Sequence and Appropriate Ratio of Amino Acid
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kim, S.W.; Hurley, W.L.; Wu, G.; Fu, J. Ideal amino acid balance for sows during gestation and lactation. J. Anim. Sci. 2009, 87, E123–E132. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Hristov, A.N.; Cassidy, T.W.; Heyler, K.S.; Lapierre, H.; Varga, G.A.; Veth, M.D.; Patton, R.A.; Parys, C. Rumen-protected lysine, methionine, and histidine increase milk protein yield in dairy cows fed a metabolizable protein-deficient diet. J. Dairy Sci. 2012, 95, 6042–6056. [Google Scholar] [CrossRef]
- Lean, I.J.; Ondarza, M.B.; Sniffen, C.J.; Santos, J.P.; Griswold, K.E. Meta-analysis to predict the effects of metabolizable amino acids on dairy cattle performance. J. Dairy Sci. 2018, 101, 340–364. [Google Scholar] [CrossRef] [PubMed]
- Dorigam, J.C.; Sakomura, N.K.; Lima, M.B.; Sarcinelli, M.F.; Suzuki, R.M. Establishing an essential amino acid profile for maintenance in poultry using deletion method. J. Anim. Physiol. 2016, 100, 884–892. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Diao, Q.Y.; Xu, X.C.; Tu, Y.; Zhang, N.F.; Yun, Q. The limiting sequence and proper ratio of lysine, methionine and threonine for calves fed milk replacers containing soy protein. Asian-Australas. J. Anim. Sci. 2012, 25, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Ragland-Gray, K.K.; Amos, H.E.; Mccann, M.A.; Williams, C.C.; Sartin, J.L.; Barb, C.R.; Kautz, F.M. Nitrogen metabolism and hormonal responses of steers fed wheat silage and infused with amino acids or casein. J. Anim. Sci. 1997, 75, 3038–3045. [Google Scholar] [CrossRef]
- Hill, T.M.; Bateman, H.G.; Aldrich, J.M.; Schlotterbeck, R.L.; Tanan, K.G. Optimal concentrations of lysine, methionine, and threonine in milk replacers for calves less than five weeks of age. J. Dairy Sci. 2008, 91, 2433–2442. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Diao, Q.Y.; Xu, X.C.; Tu, Y.; Zhang, N.F.; Yun, Q. Effects of Dietary Addition Pattern of Lysine, Methionine and Threonine in the Diet on Growth Performance, Nutrient Digestion and Metabolism, and Serum Biochemical Parameters in Calves at the Ages of 0–2 Months. Sci. Agric. Sin. 2011, 44, 1898–1907. [Google Scholar]
- Wang, T.C.; Fuller, M.F. An Optimal Dietary Amino Acid Pattern for Growing Pigs. Br. J. Nutr. 1989, 62, 77–89. [Google Scholar] [CrossRef]
- Zinn, R.A.; Shen, Y. An evaluation of ruminally degradable intake protein and metabolizable amino acid requirements of feedlot calves. J. Anim. Sci. 1998, 76, 1280–1289. [Google Scholar] [CrossRef]
- Chung, T.K.; Baker, D.H. Ideal amino acid pattern for 10-kilogram pigs [L500, L600], pigs, amino acids, growth, nitrogen metabolism, nitrogen balance, diet IND92069907. J. Anim. Sci. 1992, 62, 77–89. [Google Scholar]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis, 15th ed.; AOAC: Washington, DC, USA, 1990. [Google Scholar]
- Soest, P.J.V.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Ludden, P.A.; Kerley, M.S. Amino acid and energy interrelationships in growing beef steers, II. Effects of energy intake and metabolizable lysine supply on growth. J. Anim. Sci. 1998, 76, 3157–3168. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Awawdeh, M.S.; Titgemeyer, E.C.; Schroeder, G.F.; Gnad, D.P. Excess amino acid supply improves methionine and leucine utilization by growing steers. J. Anim. Sci. 2006, 84, 1801–1809. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, H.Y.; Wang, Y.M.; Yang, Z.Q.; Liu, J.X.; Wu, Y.M.; Yan, T.; Ye, H.W. Effects of dietary supplementation of methionine and lysine on milk production and nitrogen utilization in dairy cows. J. Dairy Sci. 2010, 93, 3661–3670. [Google Scholar] [CrossRef]
- Awawdeh, M.S.; Titgemeyer, E.C.; Mccuistion, K.C.; Gnad, D.P. Effects of ammonia load on methionine utilization by growing steers. J. Anim. Sci. 2004, 82, 3537–3542. [Google Scholar] [CrossRef]
- Kohn, R.A.; Dinneen, M.M.; Russek-Cohen, E. Using blood urea nitrogen to predict nitrogen excretion and efficiency of nitrogen utilization in cattle, sheep, goats, horses, pigs, and rats. J. Anim. Sci. 2005, 83, 879–886. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Dixon, R.M. Intake, growth and carcass characteristics of lambs consuming low digestible hay and cereal grain. Anim. Feed Sci. Technol. 2004, 114, 31–41. [Google Scholar] [CrossRef]
- Jiang, S.Z.; Yang, W.R.; Liu, F.X.; Yang, Z.B. Study on Effects of Methionine Medicate into the Rumen on Apparent Digestibility of Amino Acid in the Small Intestine of the Steers. Acta Vet. Zootech. Sin. 2006, 12, 957–962. [Google Scholar]
- Wang, C.; Liu, J.X.; Yuan, Z.P.; Wu, Y.M.; Zhai, S.W.; Ye, H.W. Effect of level of metabolizable protein on milk production and nitrogen utilization in lactating dairy cows. J. Dairy Sci. 2007, 90, 2960–2965. [Google Scholar] [CrossRef]
- Socha, M.T.; Putnam, D.E.; Garthwaite, B.D.; Whitehouse, N.L.; Kierstead, N.A.; Schwab, C.G.; Ducharme, G.A.; Robert, J.C. Improving intestinal amino acid supply of pre- and postpartum dairy cows with rumen-protected methionine and lysine. J. Dairy Sci. 2005, 88, 1113–1126. [Google Scholar] [CrossRef]
- Goodband, B.; Tokach, M.; Dritz, S.; Derouchey, J.; Woodworth, J. Practical starter pig amino acid requirements in relation to immunity, gut health and growth performance. J. Anim. Sci. Biotechnol. 2014, 5, 251–261. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lin, X.J.; Jiang, S.Q.; Ding, F.Y.; Zheng, C.T.; Hong, P. Science IOA. Amino Acid Balance Pattern of a Low Protein Diet in Fast-Growing Yellow-Feathered Broilers Aged from 1 to 21 Days. Chin. J. Anim. Nutr. 2014, 26, 2542–2552. [Google Scholar]
- Haque, M.N.; Guinard-Flament, J.; Lamberton, P.; Mustière, C.; Lemosquet, S. Changes in mammary metabolism in response to the provision of an ideal amino acid profile at 2 levels of metabolizable protein supply in dairy cows, Consequences on efficiency. J. Dairy Sci. 2015, 98, 3951–3968. [Google Scholar] [CrossRef]
- Haque, M.N.; Rulquin, H.; Lemosquet, S. Milk protein responses in dairy cows to changes in postruminal supplies of arginine, isoleucine, and valine. J. Dairy Sci. 2013, 96, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.N.; Rulquin, H.; Lemosquet, S. Milk protein synthesis in response to the provision of an “ideal” amino acid profile at 2 levels of metabolizable protein supply in dairy cows. J. Dairy Sci. 2012, 95, 5876–5887. [Google Scholar] [CrossRef]
- Schingoethe, D.J. Balancing the amino acid needs of the dairy cow. Anim. Feed Sci. Technol. 1996, 60, 153–160. [Google Scholar] [CrossRef]
- Agle, M.; Hristov, A.N.; Zaman, S.; Schneider, C.; Ndegwa, P.; Vaddella, V.K. The effects of ruminally degraded protein on rumen fermentation and ammonia losses from manure in dairy cows. J. Dairy Sci. 2010, 93, 1625–1637. [Google Scholar] [CrossRef]
- Colmenero, J.J.O.; Broderick, G.A. Effect of dietary crude protein concentration on milk production and nitrogen utilization in lactating dairy cows. J. Dairy Sci. 2006, 89, 1704–1712. [Google Scholar] [CrossRef]
- Abd, P.; Whitehouse, N.L.; Aragona, K.M.; Schwab, C.S.; Reis, S.F.; Brito, A.F. Production and nitrogen utilization in lactating dairy cows fed ground field peas with or without ruminally protected lysine and methionine. J. Dairy Sci. 2017, 100, 6239–6255. [Google Scholar]
- Xue, F.; Zhou, Z.; Ren, L.; Meng, Q. Influence of rumen-protected lysine supplementation on growth performance and plasma amino acid concentrations in growing cattle offered the maize stalk silage/maize grain-based diet. Anim. Feed Sci. Technol. 2011, 169, 61–67. [Google Scholar] [CrossRef]
- Klemesrud, M.J.; Klopfenstein, T.J.; Lewis, A.J. Metabolize methionine and lysine requirements of growing cattle. J. Anim. Sci. 2000, 78, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Doepel, L.; Pacheco, D.; Kennelly, J.J.; Hanigan, M.D.; López, I.F.; Lapierre, H. Milk protein synthesis as a function of amino acid supply. J. Dairy Sci. 2004, 87, 1279–1297. [Google Scholar] [PubMed]
- NRC (Nutrient Requirements of Swine); National Academy Press: Washington, DC, USA, 2012.
- Christian, W.; Frank, L. Improving the reliability of optimal in-feed amino acid ratios based on individual amino acid efficiency data from N balance studies in growing chicken. Animals 2013, 3, 558–573. [Google Scholar]
- Gerrits, W.J.; France, J.; Dijkstra, J.; Bosch, M.W.; Tolman, G.H.; Tamminga, S. Evaluation of a model integrating protein and energy metabolism in preruminant calves. J. Nutr. 1997, 127, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- NRC (Nutrient Requirements of Beef Cattle); National Academy Press: Washington, DC, USA, 2016.




{kind=link}
{kind=link}
{kind=link}
| Ingredients | Contents, % | Nutrient Levels 2 | Levels |
|---|---|---|---|
| Corn | 45.67 | Metabolizable energy, (MJ/kg) | 10.13 |
| Soybean meal | 11.97 | Crude protein, % | 14.95 |
| Wheat bran | 15 | Ether extract, % | 3.04 |
| Alfalfa hay | 25 | Ash, % | 7.58 |
| Limestone | 1.06 | Neutral detergent fiber, % | 29.22 |
| Salt | 0.3 | Acid detergent fiber, % | 13.99 |
| Premix 1 | 1 | Calcium, % | 1.12 |
| Total | 100 | Phosphorus, % | 0.60 |
| Lysine, % | 0.51 | ||
| Methionine, % | 0.07 | ||
| Threonine, % | 0.49 |
| Items | Treatments 1 (%) | |||
|---|---|---|---|---|
| PC | PD–Lys | PD–Met | PD–Thr | |
| Total AA content | ||||
| Lysine | 1.00 | 0.66 | 1.00 | 1.00 |
| Methionine | 0.33 | 0.33 | 0.22 | 0.33 |
| Threonine | 0.72 | 0.72 | 0.72 | 0.45 |
| AA content in basal diet | ||||
| Lysine | 0.51 | 0.51 | 0.51 | 0.51 |
| Methionine | 0.07 | 0.07 | 0.07 | 0.07 |
| Threonine | 0.45 | 0.45 | 0.45 | 0.45 |
| Exogenously added AA | ||||
| Lysine | 0.49 | 0.15 | 0.49 | 0.49 |
| Methionine | 0.25 | 0.25 | 0.15 | 0.25 |
| Threonine | 0.23 | 0.23 | 0.23 | 0.00 |
| Items 1 | Treatments 2 | SEM | p Value 3 | |||||
|---|---|---|---|---|---|---|---|---|
| PD–Lys | PD–Met | PD–Thr | PC | T | M | T × M | ||
| BW, kg | ||||||||
| Average | 273.7 | 273.9 | 276.3 | 274.8 | 2.98 | 0.4798 | <0.0001 | <0.0001 |
| 6 mon | 227.5 | 227.0 | 229.9 | 228.5 | 4.08 | 0.4647 | ||
| 7 mon | 258.0 | 257.8 | 259.3 | 257.6 | 4.00 | 0.7016 | ||
| 8 mon | 282.7 | 284.3 | 282.5 | 280.3 | 4.52 | 0.3667 | ||
| 9 mon | 326.4 | 326.8 | 333.4 | 333.0 | 4.02 | 0.0997 | ||
| ADG, kg | ||||||||
| Average | 1.04 | 1.09 | 1.09 | 1.11 | 0.029 | 0.1566 | <0.0001 | <0.0001 |
| 6–7 mon | 0.98 | 1.02 | 0.94 | 0.95 | 0.075 | 0.2848 | ||
| 7–8 mon | 0.95 | 0.95 | 0.90 | 0.89 | 0.079 | 0.4946 | ||
| 8–9 mon | 1.21 c | 1.30 bc | 1.44 ab | 1.48 a | 0.080 | 0.0013 | ||
| DMI, kg | ||||||||
| Average | 7.15 | 7.16 | 7.16 | 7.05 | 0.050 | 0.1073 | <0.0001 | <0.0001 |
| 6–7 mon | 6.25 | 6.20 | 6.26 | 6.28 | 0.091 | 0.2942 | ||
| 7–8 mon | 6.99 | 6.98 | 6.94 | 6.84 | 0.089 | 0.0916 | ||
| 8–9 mon | 8.21 | 8.28 | 8.16 | 8.25 | 0.088 | 0.2297 | ||
| Feed conversion rate, G/F | ||||||||
| Average | 0.146 | 0.152 | 0.152 | 0.156 | 0.008 | 0.1452 | 0.0001 | 0.0002 |
| 6–7 mon | 0.156 | 0.164 | 0.151 | 0.155 | 0.011 | 0.2138 | ||
| 7–8 mon | 0.135 | 0.134 | 0.127 | 0.130 | 0.010 | 0.5009 | ||
| 8–9 mon | 0.147 c | 0.158 bc | 0.176 ab | 0.181 a | 0.012 | 0.0057 | ||
| Items 1 | Treatments 2 | SEM | p Value | |||
|---|---|---|---|---|---|---|
| PD–Lys | PD–Met | PD–Thr | PC | |||
| Intake N, g·(kg−1 BW0.75) d−1 | 2.92 | 2.97 | 2.90 | 2.87 | 0.020 | 0.2961 |
| Fecal N, g·(kg−1 BW0.75)·d−1 | 0.90 | 0.82 | 0.80 | 0.79 | 0.015 | 0.1223 |
| Urine N, g·(kg−1 BW0.75)·d−1 | 1.06 b | 1.16 a | 1.02 b | 0.88 c | 0.033 | 0.0011 |
| Total excrete N, g·(kg−1 BW0.75)·d−1 | 1.96 b | 1.98 a | 1.82 ab | 1.67 ab | 0.032 | 0.0208 |
| N retention, g·(kg−1 BW0.75)·d−1 | 0.96 b | 0.99 b | 1.08 ab | 1.20 a | 0.034 | 0.0324 |
| Digestible N, g·(kg−1 BW0.75)·d−1 | 2.02 | 2.15 | 2.06 | 2.08 | 0.020 | 0.2908 |
| N utilization, % | 33.08 b | 33.26 b | 34.96 b | 41.77 a | 1.210 | 0.0048 |
| N digestibility, % | 69.50 | 72.38 | 70.79 | 72.6 | 0.512 | 0.0798 |
| Items 1 | Based on Metabolic Body Weight, g·(kg−1 BW0.75)·d−1 | The Ratio to PC | ||||||
|---|---|---|---|---|---|---|---|---|
| NR 2 | AAI 2 | NR | AAI | |||||
| Lys | Met | Thr | Lys | Met | Thr | |||
| PD–Lys | 0.96 | 0.60 | 0.29 | 0.65 | 0.80 | 0.67 | 1.00 | 1.00 |
| PD–Met | 0.99 | 0.90 | 0.20 | 0.65 | 0.83 | 1.00 | 0.69 | 1.00 |
| PD–Thr | 1.02 | 0.90 | 0.29 | 0.40 | 0.85 | 1.00 | 1.00 | 0.62 |
| PC | 1.20 | 0.90 | 0.29 | 0.65 | 1.00 | 1.00 | 1.00 | 1.00 |
| Items 1 | S | P | C | R |
|---|---|---|---|---|
| Lys | 0.61 | 1.00 | 69.99 | 100.00 |
| Met | 0.57 | 0.98 | 22.36 | 31.95 |
| Thr | 0.26 | 0.79 | 39.78 | 56.84 |
| Stage | Index 1 | Lys: Met: Thr Ratio | Reference |
|---|---|---|---|
| Calves | NR | 100:26:66 | Gerrits et al., 1997 [37] |
| Calves | maximum ADG | 100:31:77 | Hill et al., 2008 [7] |
| Calves | maximum ADG | 100:35:63 | Wang et al., 2011 [8] |
| Beef cattle | body amino acids | 100:31:61 | NRC (2016) [38] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Bi, Y.; Diao, Q.; Piao, M.; Wang, B.; Kong, F.; Hu, F.; Tang, M.; Sun, Y.; Tu, Y. The Limiting Sequence and Appropriate Amino Acid Ratio of Lysine, Methionine, and Threonine for Seven- to Nine-Month-Old Holstein Heifers Fed Corn–Soybean M-Based Diet. Animals 2019, 9, 750. https://doi.org/10.3390/ani9100750
Li Y, Bi Y, Diao Q, Piao M, Wang B, Kong F, Hu F, Tang M, Sun Y, Tu Y. The Limiting Sequence and Appropriate Amino Acid Ratio of Lysine, Methionine, and Threonine for Seven- to Nine-Month-Old Holstein Heifers Fed Corn–Soybean M-Based Diet. Animals. 2019; 9(10):750. https://doi.org/10.3390/ani9100750
Chicago/Turabian StyleLi, Yuan, Yanliang Bi, Qiyu Diao, Minyu Piao, Bing Wang, Fanlin Kong, Fengming Hu, Mengqi Tang, Yu Sun, and Yan Tu. 2019. "The Limiting Sequence and Appropriate Amino Acid Ratio of Lysine, Methionine, and Threonine for Seven- to Nine-Month-Old Holstein Heifers Fed Corn–Soybean M-Based Diet" Animals 9, no. 10: 750. https://doi.org/10.3390/ani9100750
APA StyleLi, Y., Bi, Y., Diao, Q., Piao, M., Wang, B., Kong, F., Hu, F., Tang, M., Sun, Y., & Tu, Y. (2019). The Limiting Sequence and Appropriate Amino Acid Ratio of Lysine, Methionine, and Threonine for Seven- to Nine-Month-Old Holstein Heifers Fed Corn–Soybean M-Based Diet. Animals, 9(10), 750. https://doi.org/10.3390/ani9100750