The Effect of Virtual Fencing Stimuli on Stress Responses and Behavior in Sheep


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Habituation
2.3. Experimental Design and Treatments
- Control animals entered the testing area for 1 min and were not subjected to stimuli.
- Beep treatment animals were fitted with remotely controlled Garmin collars prior to entry to the testing area and the beep tone (45–55 dB, 2.7 kHz) was manually applied to each of the four collared animals simultaneously three times for two seconds with a two-second interval in between, two experimenters each operated two collar control devices. The duration was based upon the virtual fencing protocol developed by Marini, Meuleman, Belson, Rodenburg, Llewellyn, and Lee [5].
- Bark treatment involved playing the sound of a dog barking (58–68 dB, 6.1 kHz) through a speaker placed centrally in the testing area, with 3 s played for 2 s each with an interval of 2 s in between.
- Restraint treatment involved handlers entering the test area and each catching one sheep by hand (one handler per sheep). The sheep was then placed in an inverted restraint position (ventral belly exposed) for a period of 60 s, such as sheep would experience undergoing normal shearing practices.
- Electrical stimuli (ES) treatment animals were fitted with Garmin dog control collars (Garmin TT15, Garmin Ltd., Kansas, KS, USA) placed snugly around the neck below the jaw, ensuring that the probes were in close contact with the skin. To ensure good contact the animals in this treatment group had their necks shorn more extensively the day prior to testing. Three electrical stimuli were applied to each of the four collared animals simultaneously for approximately 500–600 milliseconds each, with intervals of two seconds in between each application, with two operators as per beep treatment. The electrical stimulus was set to level 4 (320 V, 20 us, 16 pulses per/sec) out of a possible 18. The level chosen has been identified in previous studies as being sufficiently aversive to achieve the desired conditioned response, but not so aversive as to cause negative behavioural responses such as jumping and running around [5]. The level chosen is much lower than traditional electric fencing, in which industry standards recommend 4000–5000V for sheep [22], and lower than that used on sheep in previous research by Brunberg et al. [23] at 4000V and Martin et al. [24] at 6500V. The intensity of the electrical stimulus used was also lower than that used in a similar experiment conducted by Lee, Fisher, Reed, and Henshall [9] on cattle at 600V.
2.4. Body Temperature and Blood Sampling
2.5. Behavioral Analysis
2.6. Statistical Analysis
3. Results
3.1. Physiological Measures
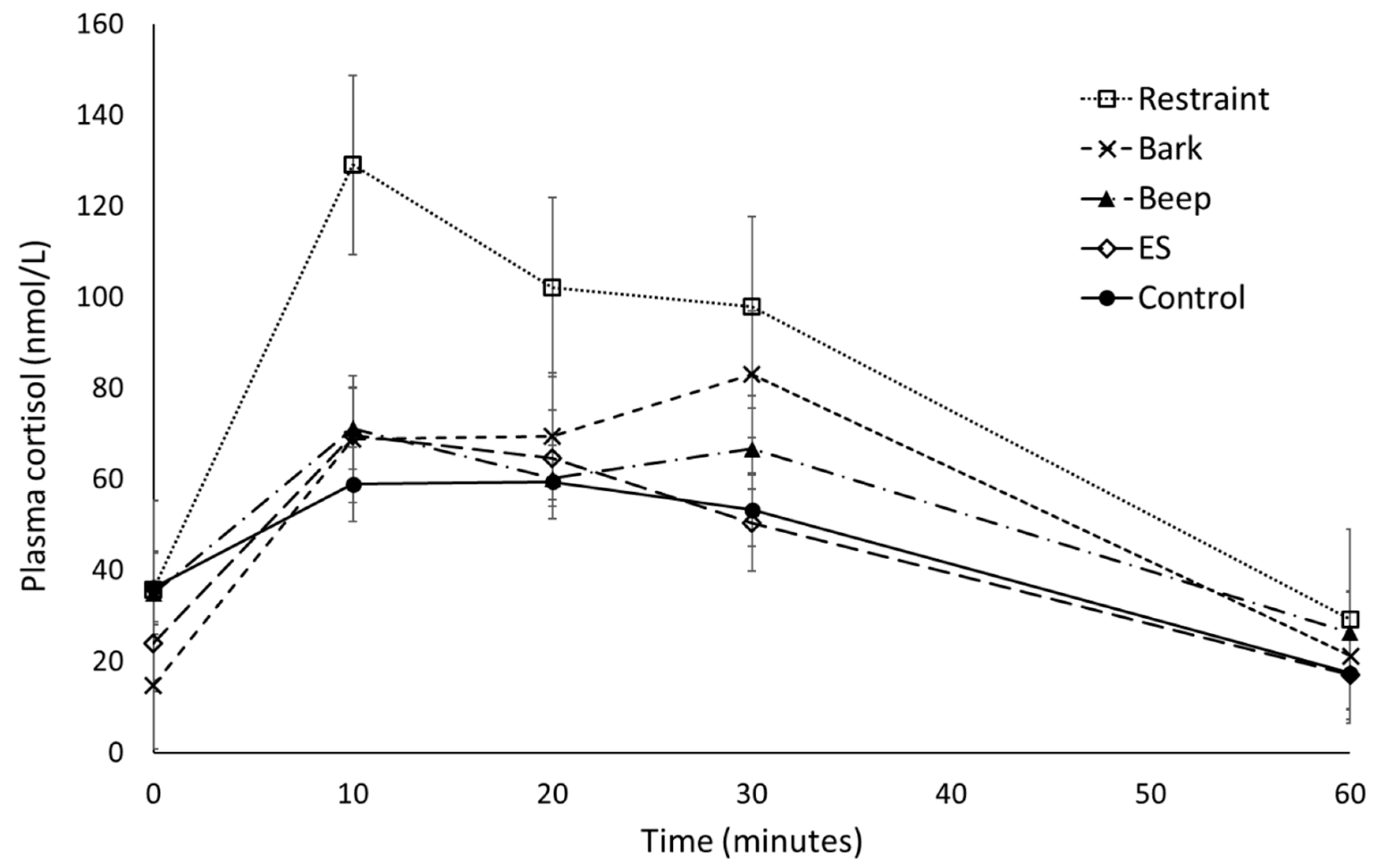
3.1.1. Plasma Cortisol
3.1.2. Vaginal Temperature
3.2. Behavioral Measures (Excluding Restraint Treatment)
3.2.1. Vigilance Behaviors
3.2.2. Ear Positions
3.2.3. Point Behaviors
4. Discussion
4.1. Physiological Responses
4.1.1. Audio Stimuli
4.1.2. Physical Stimuli
4.2. Behavioral Responses
4.2.1. Audio Stimuli
4.2.2. Physical Stimuli
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, C.; Henshall, J.M.; Wark, T.J.; Crossman, C.C.; Reed, M.T.; Brewer, H.G.; O’Grady, J.; Fisher, A.D. Associative learning by cattle to enable effective and ethical virtual fences. Appl. Anim. Behav. Sci. 2009, 119, 15–22. [Google Scholar] [CrossRef]
- Lee, C. An Apparatus and Method for the Virtual Fencing of an Animal. Available online: https://patentscope.wipo.int/search/en/detail.jsf?docId=WO2006007643 (accessed on 9 January 2019).
- Campbell, D.L.M.; Lea, J.M.; Farrer, W.J.; Haynes, S.J.; Lee, C. Tech-savvy beef cattle? How heifers respond to moving virtual fence lines. Animals 2017, 7, 72. [Google Scholar] [CrossRef]
- Campbell, D.L.M.; Lea, J.M.; Haynes, S.J.; Farrer, W.J.; Leigh-Lancaster, C.J.; Lee, C. Virtual fencing of cattle using an automated collar in a feed attractant trial. Appl. Anim. Behav. Sci. 2018, 200, 71–77. [Google Scholar] [CrossRef]
- Marini, D.; Meuleman, M.; Belson, S.; Rodenburg, T.; Llewellyn, R.; Lee, C. Developing an ethically acceptable virtual fencing system for sheep. Animals 2018, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Colditz, I.G.; Campbell, D.L.M. A framework to assess the impact of new animal management technologies on welfare: A case study of virtual fencing. Front. Vet. Sci. 2018, 5, 187. [Google Scholar] [CrossRef] [PubMed]
- Destrez, A.; Deiss, V.; Leterrier, C.; Boivin, X.; Boissy, A. Long-term exposure to unpredictable and uncontrollable aversive events alters fearfulness in sheep. Animal 2013, 7, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.M. Psychological factors in stress and disease. Sci. Am. 1972, 226, 104–113. [Google Scholar] [CrossRef]
- Lee, C.; Fisher, A.D.; Reed, M.T.; Henshall, J.M. The effect of low energy electric shock on cortisol, β-endorphin, heart rate and behaviour of cattle. Appl. Anim. Behav. Sci. 2008, 113, 32–42. [Google Scholar] [CrossRef]
- Hargreaves, A.L.; Hutson, G.D. The stress response in sheep during routine handling procedues. Appl. Anim. Behav. Sci. 1990, 26, 83–90. [Google Scholar] [CrossRef]
- Heffner, H.E. Auditory awareness. Appl. Anim. Behav. Sci. 1998, 57, 259–268. [Google Scholar] [CrossRef]
- Andersson, I. Behaviour and Heart Rate in Sheep When Herded by Border Collies with Different Background; Student report; Swedish University of Agricultural Sciences: Upsala, Sweden, 2015. [Google Scholar]
- Beausoleil, N.J. Behavioural and Physiological Responses of Domestic Sheep (Ovis aries) to the Presence of Humans and Dogs. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 2006. [Google Scholar]
- Harlow, H.J.; Thorne, E.T.; Williams, E.S.; Belden, E.L.; Gern, W.A. Adrenal responsiveness in domestic sheep (Ovis aries) to acute and chronic stressors as predicted by remote monitoring of cardiac frequency. Can. J. Zool. 1987, 65, 2021–2027. [Google Scholar] [CrossRef]
- Hargreaves, A.L.; Hutson, G.D. Some effects of repeated handling on stress responses in sheep. Appl. Anim. Behav. Sci. 1990, 26, 253–263. [Google Scholar] [CrossRef]
- Hargreaves, A.L.; Hutson, G.D. Changes in heart rate, plasma cortisol and haematocrit of sheep during a shearing procedure. Appl. Anim. Behav. Sci. 1990, 26, 91–101. [Google Scholar] [CrossRef]
- Adriaan Bouwknecht, J.; Olivier, B.; Paylor, R.E. The stress-induced hyperthermia paradigm as a physiological animal model for anxiety: A review of pharmacological and genetic studies in the mouse. Neurosci. Biobehav. Rev. 2007, 31, 41–59. [Google Scholar] [CrossRef]
- Pedernera-Romano, C.; Ruiz De La Torre, J.L.; Badiella, L.; Manteca, X. Associations between open-field behaviour and stress-induced hyperthermia in two breeds of sheep. Anim. Welf. 2011, 20, 339–346. [Google Scholar]
- Sanger, M.E.; Doyle, R.E.; Hinch, G.N.; Lee, C. Sheep exhibit a positive judgement bias and stress-induced hyperthermia following shearing. Appl. Anim. Behav. Sci. 2011, 131, 94–103. [Google Scholar] [CrossRef]
- Boissy, A.; Aubert, A.; Desire, L.; Greiveldinger, L.; Delval, E.; Veissier, I. Cognitive sciences to relate ear postures to emotions in sheep. Anim. Wel. 2011, 20, 47–56. [Google Scholar]
- Frid, A. Vigilance by female dall’s sheep: Interactions between predation risk factors. Anim. Behav. 1997, 53, 799–808. [Google Scholar] [CrossRef]
- Staffix. Animal Voltage Requirements. Available online: https://www.stafix.com/en-us/helpful-information/animal-voltage-requirements (accessed on 9 January 2019).
- Brunberg, E.I.; Bergslid, I.K.; Boe, K.E.; Sorheim, K.M. The ability of ewes with lambs to learn a virtual fencing system. Animal 2017, 11, 2045–2050. [Google Scholar] [CrossRef]
- Martin, M.; Pritchard, D.; Stephenson, R. Electric shock conditioning of sheep to avoid feed supplement troughs in field investigations of diet supplementation. Rangeland J. 1989, 11, 44–46. [Google Scholar] [CrossRef]
- Lea, J.M.; Niemeyer, D.D.O.; Reed, M.T.; Fisher, A.D.; Ferguson, D.M. Development and validation of a simple technique for logging body temperature in free-ranging cattle. Aust. J. Exp. Agr. 2008, 48, 741–745. [Google Scholar] [CrossRef]
- Paull, D.R.; Lee, C.; Colditz, I.G.; Atkinson, S.J.; Fisher, A.D. The effect of a topical anaesthetic formulation, systemic flunixin and carprofen, singly or in combination, on cortisol and behavioural responses of merino lambs to mulesing. Aust. Vet. J. 2007, 85, 98–106. [Google Scholar] [CrossRef]
- Greiveldinger, L.; Veissier, I.; Boissy, A. Behavioural and physiological responses of lambs to controllable vs. Uncontrollable aversive events. Psychoneuroendocrinology 2009, 34, 805–814. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; 3.3.1; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Nlme: Linear and Nonlinear Mixed Effects Models; R package, 3.1-137; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Jackman, S. Pscl: Classes and methods for r developed in the political science computational laboratory; 1.5.2; United States Studies Centre, University of Sydney: Sydney, New South Wales, Australia, 2017. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Mass r package. Modern Applied Statistics with S, Fourth ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Mangiafico, S. Rcompanion: Functions to Support Extension Education Program Evaluation; R package, 1.13.2; Rutgers: Camden, NJ, USA, 2018. [Google Scholar]
- Peters, G. Userfriendlyscience: Quantitative Analysis Made Accessible, 2018. Available online: https://cran.r-project.org/web/packages/userfriendlyscience/userfriendlyscience.pdf (accessed on 9 January 2019).
- Niezgoda, J.; Bobek, S.; Wronska-Fortuna, D.; Wierzchos, E. Response of sympatho-adrenal axis and adrenal cortex to short-term restraint stress in sheep. J. Vet. Med. 1993, 40, 631–638. [Google Scholar] [CrossRef]
- Gliner, J.A. Predictable vs. Unpredictable shock: Preference behavior and stomach ulceration. Physiol. Behav. 1972, 9, 693–698. [Google Scholar] [CrossRef]
- Greiveldinger, L.; Veissier, I.; Boissy, A. Emotional experience in sheep: Predictability of a sudden event lowers subsequent emotional responses. Physiol. Behav. 2007, 92, 675–683. [Google Scholar] [CrossRef]
- De, K.; Saxena, V.K.; Balaganur, K.; Kumar, D.; Naqvi, S.M.K. Effect of short-term seclusion of sheep on their welfare indicators. J. Vet. Behav. 2018, 27, 1–7. [Google Scholar] [CrossRef]
- Ciechanowska, M.; Lapot, M.; Antkowiak, B.; Mateusiak, K.; Paruszewska, E.; Malewski, T.; Paluch, M.; Przekop, F. Effect of short-term and prolonged stress on the biosynthesis of gonadotropin-releasing hormone (gnrh) and gnrh receptor (gnrhr) in the hypothalamus and gnrhr in the pituitary of ewes during various physiological states. Anim. Reprod. Sci. 2016, 174, 65–72. [Google Scholar] [CrossRef]
- Veissier, I.; Boissy, A. Stress and welfare: Two complementary concepts that are intrinsically related to the animal’s point of view. Physiol. Behav. 2007, 92, 429–433. [Google Scholar] [CrossRef]
- Lee, T.K.; Lee, C.; Bischof, R.; Lambert, G.W.; Clarke, I.J.; Henry, B.A. Stress-induced behavioral and metabolic adaptations lead to an obesity-prone phenotype in ewes with elevated cortisol responses. Psychoneuroendocrinology 2014, 47, 166–177. [Google Scholar] [CrossRef]
- Beausoleil, N.J.; Stafford, K.J.; Mellor, D.J. Sheep show more aversion to a dog than to a human in an arena test. Appl. Anim. Behav. Sci. 2005, 91, 219–232. [Google Scholar] [CrossRef]
- Morris, J.E.; Fisher, A.D.; Doyle, R.E.; Bush, R.D. Determination of sheep learning responses to a directional audio cue. J. Appl. Anim. Welf. Sci. 2010, 13, 347–360. [Google Scholar] [CrossRef]
- Hopewell, L.; Rossiter, R.; Blower, E.; Leaver, L.; Goto, K. Grazing and vigilance by soay sheep on lundy island: Influence of group size, terrain and the distribution of vegetation. Behav. Processes 2005, 70, 186–193. [Google Scholar] [CrossRef]
- Mooring, M.S.; Fitzpatrick, T.A.; Nishihira, T.T.; Reisig, D.D. Vigilance, predation risk, and the allee effect in desert bighorn sheep. J. Wildl. Manage. 2004, 68, 519–532. [Google Scholar] [CrossRef]
- Quenette, P.Y. Functions of vigilance behavior in mammals―A review. Acta Oecol. 1990, 11, 801–818. [Google Scholar]
- Lee, C.; Verbeek, E.; Doyle, R.; Bateson, M. Attention bias to threat indicates anxiety differences in sheep. Biol. Lett. 2016, 12. [Google Scholar] [CrossRef]
- Monk, J.E.; Doyle, R.E.; Colditz, I.G.; Belson, S.; Cronin, G.M.; Lee, C. Towards a more practical attention bias test to assess affective state in sheep. PLoS ONE 2018, 13, e0190404. [Google Scholar] [CrossRef]
- Bassett, L.; Buchanan-Smith, H.M. Effects of predictability on the welfare of captive animals. Appl. Anim. Behav. Sci. 2007, 102, 223–245. [Google Scholar] [CrossRef]
- Desire, L.; Veissier, I.; Despres, G.; Boissy, A. On the way to assess emotions in animals: Do lambs (Ovis aries) evaluate an event through its suddenness, novelty, or unpredictability? J. Comp. Psychol. 2004, 118, 363–374. [Google Scholar] [CrossRef]
- Reefmann, N.; Bütikofer Kaszàs, F.; Wechsler, B.; Gygax, L. Ear and tail postures as indicators of emotional valence in sheep. Appl. Anim. Behav. Sci. 2009, 118, 199–207. [Google Scholar] [CrossRef]
- Tamioso, P.R.; Rucinque, D.S.; Taconeli, C.A.; da Silva, G.P.; Molento, C.F.M. Behavior and body surface temperature as welfare indicators in selected sheep regularly brushed by a familiar observer. J. Vet. Behav. Clin. Appl. Res. 2017, 19, 27–34. [Google Scholar] [CrossRef]
- Reefmann, N.; Muehlemann, T.; Wechsler, B.; Gygax, L. Housing induced mood modulates reactions to emotional stimuli in sheep. Appl. Anim. Behav. Sci. 2012, 136, 146–155. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameter of Behavior | Definition of Behavior (Characterisation) | Point or Continuous Sampling |
|---|---|---|
| Vigilance | Vigilant—head above shoulder (Vigilance) | Continuous |
| Not vigilant—head parallel to or below shoulder height | ||
| Ear posture | Ears in different directions (Axial) | Continuous |
| Both ears facing backward (Backward) | ||
| Both ears neutral position (Planar) | ||
| Both ears facing forward (Forward) | ||
| Elimination | Urination (Elimination) | Point |
| Defecation (Elimination) | ||
| Movement | One step of each leg or pair of legs (Step) | Point |
| Rapid movement | Run—rapid movement (Avoidance) | Point |
| Step back—step backwards (Avoidance) | ||
| Sniffing | Sniffing other sheep (Exploratory) | Point |
| Sniffing ground (Exploratory) | ||
| Sniffing surroundings (Exploratory) | ||
| Huddling | Huddling—grouping together so they are in close body contact (Avoidance) | Point |
| Hiding—lowered head in corner or under other sheep (Avoidance) | ||
| Avoidance responses | Jump—all four feet off the ground (Avoidance) | Point |
| Rear—two feet off the ground (Avoidance) | ||
| Fall—quarters touching the ground (Avoidance) | ||
| Shaking | Shaking head (Shaking) | Point |
| Shaking body (Shaking) | ||
| Avoidance novel behaviors | Stiff neck—observed as a stretching and rigidity of the neck posture (Avoidance) | Point |
| Hunching—hunched back posture (Avoidance) |
| Treatment | Vaginal Temperature (°C) | Plasma Cortisol (nmol/L) | ||||
|---|---|---|---|---|---|---|
| Mean | Peak | Change from Baseline | Mean | Peak | Change from Baseline | |
| Mean ± s.e.m. | Mean ± s.e.m. | Mean ± s.e.m. | Mean ± s.e.m. | Mean ± s.e.m. | Mean ± s.e.m. | |
| Control | 39.2 ± 0.07 a | 39.4 ± 0.09 a | 0.5 ± 0.07 a | 53.5 ± 6.75 a | 93.9 ± 12.30 a | 77.0 ± 10.85 a |
| Beep | 39.2 ± 0.06 a | 39.4 ± 0.06 a | 0.5 ± 0.03 a | 49.7 ± 7.71 a | 84.4 ± 13.01 a | 71. 8 ± 12.14 a |
| Bark | 39.1 ± 0.07 a | 39.4 ± 0.09 a | 0.5 ± 0.05 a | 50.7 ± 7.86 a | 97.6 ± 15.16 a | 87.9 ± 13.90 a |
| Restraint | 39.1 ± 0.10 a | 39.3 ± 0.11 a | 0.4 ± 0.08 a | 84.5 ± 13.28 a | 155.4 ± 22.56 b | 130. 8 ± 17.97 b |
| ES | 39.3 ± 0.07 a | 39.5 ± 0.07 a | 0.4 ± 0.06 a | 47.9 ± 6.96 a | 80.2 ± 9.59 a | 66.9 ± 8.58 a |
| p | ns | ns | ns | ns | 0.08 | ns |
| Behavior | Treatment | ||||
| Control | Beep | Bark | ES | p | |
| µ ± s.e.m. | µ ± s.e.m. | µ ± s.e.m. | µ ± s.e.m. | ||
| Vigilance (%) | 62 ± 9.8 a | 100 ± 0.3 b | 99 ± 0.7 b | 82 ± 8.0 a | *** |
| Ears axial (%) | 15 ± 5.0 a | 16 ± 4.6 a | 22 ± 3.4 a | 17 ± 3.7 a | ns |
| Ears planar (%) | 23 ± 8.9 a | 6 ± 2.6 a | 4 ± 1.5 a | 9 ± 2.5 a | ns |
| Ears forward (%) | 51 ± 11.2 ab | 70 ± 7.6 b | 58 ± 6.3 ab | 31 ± 8.0 a | * |
| Ears backward (%) | 12 ± 6.4 a | 9 ± 3. 8 a | 16 ± 4.3 ab | 43 ± 7.3 b | *** |
| Ear posture changes (count) | 3 ± 0.5 a | 3 ± 0.5 a | 5 ± 0.5 a | 5 ± 0. 7 a | ns |
| Steps taken (count) | 1 ± 0.4 a | 1 ± 0.7 b | 5 ± 0.7 b | 5 ± 0.8 b | *** |
| Behavior | Post-treatment | ||||
| Control | Beep | Bark | ES | p | |
| µ ± s.e.m. | µ ± s.e.m. | µ ± s.e.m. | µ ± s.e.m. | ||
| Vigilance (%) | 73 ± 7.8 ac | 98 ± 1.5 bc | 100 ± 0.0 b | 55 ± 10.3 a | *** |
| Ears axial (%) | 14 ± 3.1 a | 9 ± 2.6 a | 13 ± 2.9 a | 15 ± 4.1 a | ns |
| Ears planar (%) | 13 ± 2.6 b | 2 ± 1.7 a | 1 ± 0.6 a | 6 ± 2.3 ab | *** |
| Ears forward (%) | 57 ± 5.7 a | 82 ± 4.8 b | 75 ± 4.4 ab | 59 ± 8.6 ab | * |
| Ears backward (%) | 17 ± 4.5 a | 7 ± 3.1 a | 12 ± 2.7 a | 21 ± 7.1 a | ns |
| Ear posture changes (count) | 6 ± 0.7 a | 3 ± 0.6 b | 5 ± 0.7 a | 4 ± 0.8 a | * |
| Steps taken (count) | 4 ± 0.8 b | 2 ± 0.8 a | 2 ± 0.6 b | 4 ± 0.5 b | * |
| Treatment | Exploratory | Avoidance | Elimination | Shaking | ||||
|---|---|---|---|---|---|---|---|---|
| Treatment | Post-treatment | Treatment | Post-treatment | Treatment | Post-treatment | Treatment | Post-treatment | |
| Control | 2 a | 6 a | 1 a | 1 b | 5 a | 1 a | 1 a | 2 a |
| Beep | 0 a | 0 a | 1 a | 1 b | 0 a | 1 a | 0 a | 0 a |
| Bark | 0 a | 0 a | 5 a | 9 a | 0 a | 1 a | 0 a | 0 a |
| ES | 0 a | 5 a | 14 b | 9 a | 1 a | 1 a | 4 a | 0 a |
| p | n.s | ** | *** | *** | * | n.s | * | n.s |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kearton, T.; Marini, D.; Cowley, F.; Belson, S.; Lee, C. The Effect of Virtual Fencing Stimuli on Stress Responses and Behavior in Sheep. Animals 2019, 9, 30. https://doi.org/10.3390/ani9010030
Kearton T, Marini D, Cowley F, Belson S, Lee C. The Effect of Virtual Fencing Stimuli on Stress Responses and Behavior in Sheep. Animals. 2019; 9(1):30. https://doi.org/10.3390/ani9010030
Chicago/Turabian StyleKearton, Tellisa, Danila Marini, Frances Cowley, Susan Belson, and Caroline Lee. 2019. "The Effect of Virtual Fencing Stimuli on Stress Responses and Behavior in Sheep" Animals 9, no. 1: 30. https://doi.org/10.3390/ani9010030
APA StyleKearton, T., Marini, D., Cowley, F., Belson, S., & Lee, C. (2019). The Effect of Virtual Fencing Stimuli on Stress Responses and Behavior in Sheep. Animals, 9(1), 30. https://doi.org/10.3390/ani9010030