Age-Dependent Expression of MyHC Isoforms and Lipid Metabolism-Related Genes in the Longissimus Dorsi Muscle of Wild and Domestic Pigs


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Muscle Samples
2.2. RNA Isolation, cDNA Synthesis and Quantitative Polymerase Chain Reaction (qPCR)
2.3. Histochemistry
2.4. Histological Examination
2.5. Statistical Analysis
3. Results
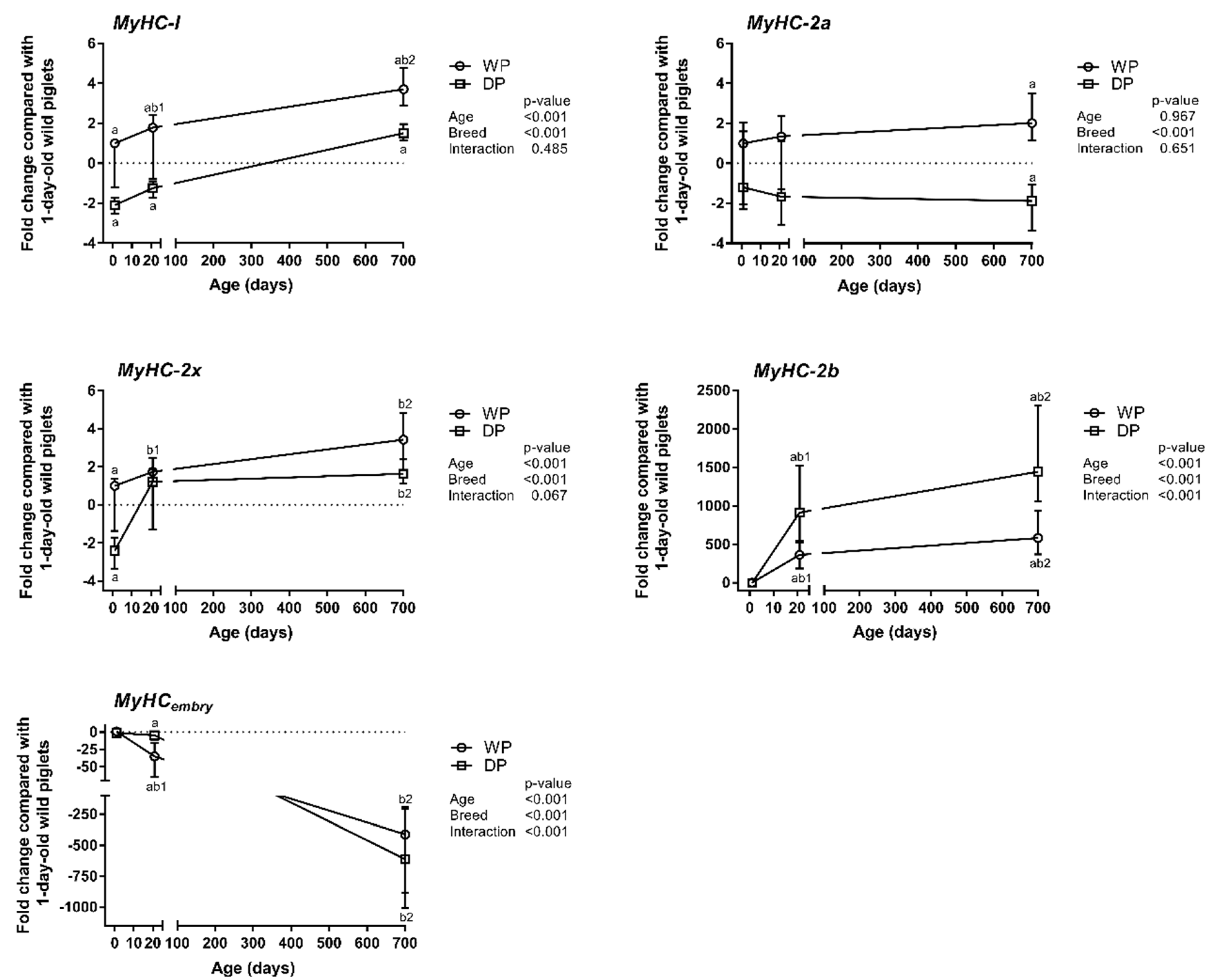
3.1. MyHC Isoform Expression Patterns
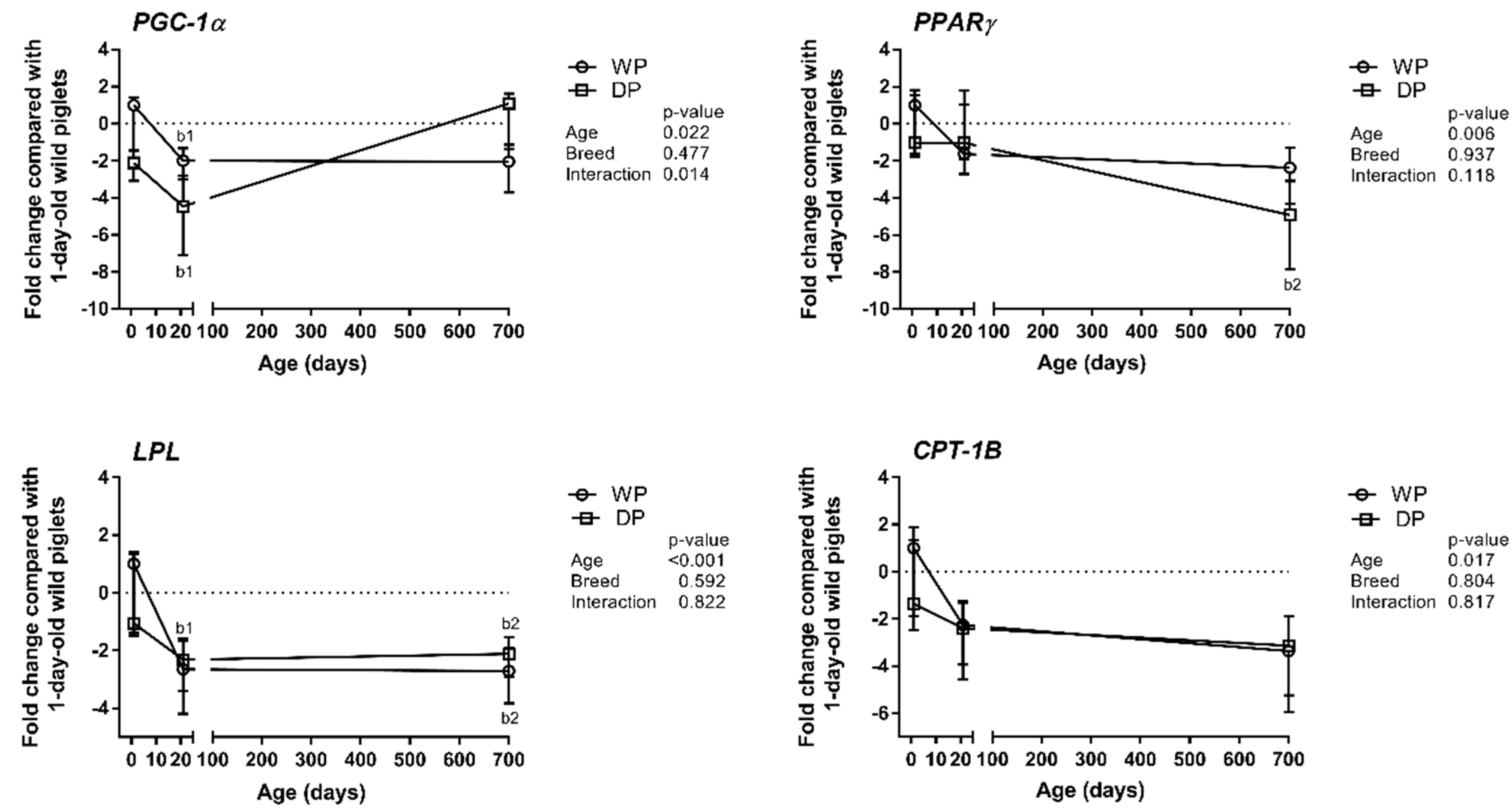
3.2. Expression Patterns of Lipid Metabolism-Related Genes
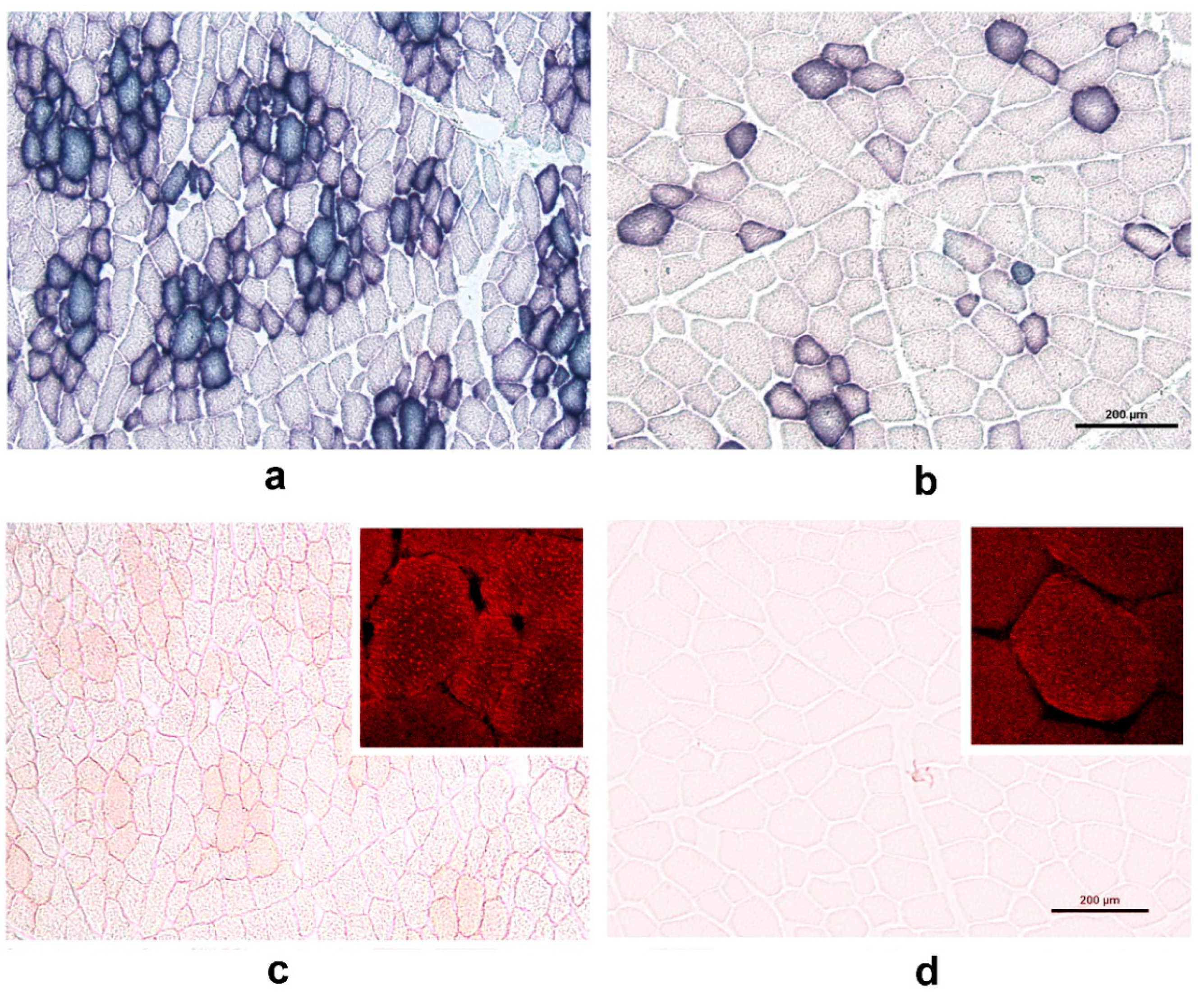
3.3. Histological Evaluation
4. Discussion
4.1. Contractile Phenotype—MyHCs Expression Patterns
4.2. Expression Patterns of Lipid Metabolism-Related Genes—Metabolic vs. Contractile Phenotype
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rehfeldt, C.; Henning, M.; Fiedler, I. Consequences of pig domestication for skeletal muscle growth and cellularity. Livest. Sci. 2008, 116, 30–41. [Google Scholar] [CrossRef]
- Lefaucheur, L. A second look into fibre typing-relation to meat quality. Meat. Sci. 2010, 84, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Gunawan, A.M.; Scheffler, T.L.; Grant, A.L.; Gerrard, D.E. Myosin heavy chain isoform content and energy metabolism can be uncoupled in pig skeletal muscle. J. Anim. Sci. 2009, 87, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Muller, E.; Rutten, M.; Moser, G.; Reiner, G.; Bartenschlager, H.; Geldermann, H. Fibre structure and metabolites in M. longissimus dorsi of wild boar, Pietrain and Meishan pigs as well as their crossbred generations. J. Anim. Breed. Genet. 2002, 119, 125–137. [Google Scholar] [CrossRef]
- Lefaucheur, L.; Hoffman, R.K.; Gerrard, D.E.; Okamura, C.S.; Rubinstein, N.; Kelly, A. Evidence for three adult fast myosin heavy chain isoforms in type II skeletal muscle fibers in pigs. J. Anim. Sci. 1998, 76, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.M.; Da Costa, N.; Chang, K.C. Cluster characterisation and temporal expression of porcine sarcomeric myosin heavy chain genes. J. Muscle Res. Cell Motil. 2003, 24, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Lefaucheur, L.; Edom, F.; Ecolan, P.; Butler-Browne, G.S. Pattern of muscle fiber type formation in the pig. Dev. Dyn. 1995, 203, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.C.; Fernandes, K. Developmental expression and 5′ end cDNA cloning of the porcine 2x and 2b myosin heavy chain genes. DNA Cell Biol. 1997, 16, 1429–1437. [Google Scholar] [CrossRef]
- Lefaucheur, L.; Ecolan, P.; Plantard, L.; Gueguen, N. New insights into muscle fiber types in the pig. J. Histochem. Cytochem. 2002, 50, 719–730. [Google Scholar] [CrossRef]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How muscle structure and composition influence meat and flesh quality. Sci. World J. 2016, 2016, 3182746. [Google Scholar] [CrossRef] [PubMed]
- Fazarinc, G.; Vrecl, M.; Skorjanc, D.; Cehovin, T.; Candek-Potokar, M. Dynamics of myosin heavy chain isoform transition in the longissimus muscle of domestic and wild pigs during growth: A comparative study. Animal 2017, 11, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.N.; Ao, Q.W.; Jiang, Q.Y.; Guo, Y.F.; Lan, G.Q.; Jiang, H.S. Comparisons of different myosin heavy chain types, AMPK, and PGC-1α gene expression in the longissimus dorsi muscles in Bama Xiang and Landrace pigs. Genet. Mol. Res. 2016, 15, gmr.15028379. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Luo, J.Q.; Zheng, P.; Yu, B.; Huang, Z.Q.; Mao, X.B.; He, J.; Yu, J.; Chen, J.L.; Chen, D.W. Differential expression of lipid metabolism-related genes and myosin heavy chain isoform genes in pig muscle tissue leading to different meat quality. Animal 2015, 9, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Palma-Granados, P.; Haro, A.; Seiquer, I.; Lara, L.; Aguilera, J.F.; Nieto, R. Similar effects of lysine deficiency in muscle biochemical characteristics of fatty and lean piglets. J. Anim. Sci. 2017, 95, 3025–3036. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Wu, H.; Tarr, P.T.; Zhang, C.Y.; Wu, Z.; Boss, O.; Michael, L.F.; Puigserver, P.; Isotani, E.; Olson, E.N.; et al. Transcriptional co-activator PGC-1 alpha drives the formation of slow-twitch muscle fibres. Nature 2002, 418, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhao, Y.; Li, R.; Gong, J.; Zheng, Y.; Wang, Y. PGC-1α is associated with C2C12 myoblast differentiation. Cent. Eur. J. Biol. 2014, 9, 1030–1036. [Google Scholar] [CrossRef]
- Erkens, T.; Vandesompele, J.; Van Zeveren, A.; Peelman, L.J. Correlation between porcine PPARGC1A mRNA expression and its downstream target genes in backfat and longissimus dorsi muscle. J. Appl. Genet. 2009, 50, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Fleige, S.; Walf, V.; Huch, S.; Prgomet, C.; Sehm, J.; Pfaffl, M.W. Comparison of relative mRNA quantification models and the impact of RNA integrity in quantitative real-time RT-PCR. Biotechnol. Lett. 2006, 28, 1601–1613. [Google Scholar] [CrossRef] [PubMed]
- da Costa, N.; McGillivray, C.; Chang, K.C. Postnatal myosin heavy chain isoforms in prenatal porcine skeletal muscles: Insights into temporal regulation. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2003, 273, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Wimmers, K.; Ngu, N.T.; Jennen, D.G.; Tesfaye, D.; Murani, E.; Schellander, K.; Ponsuksili, S. Relationship between myosin heavy chain isoform expression and muscling in several diverse pig breeds. J. Anim. Sci. 2008, 86, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Nachlas, M.M.; Tsou, K.C.; de Souza, E.; Cheng, C.S.; Seligman, A.M. Cytochemical demonstration of succinic dehydrogenase by the use of a new p-nitrophenyl substituted ditetrazole. J. Histochem. Cytochem. 1957, 5, 420–436. [Google Scholar] [CrossRef] [PubMed]
- Koopman, R.; Schaart, G.; Hesselink, M.K. Optimisation of oil red O staining permits combination with immunofluorescence and automated quantification of lipids. Histochem. Cell Biol. 2001, 116, 63–68. [Google Scholar] [PubMed]
- Chang, K.C.; da Costa, N.; Blackley, R.; Southwood, O.; Evans, G.; Plastow, G.; Wood, J.D.; Richardson, R.I. Relationships of myosin heavy chain fibre types to meat quality traits in traditional and modern pigs. Meat Sci. 2003, 64, 93–103. [Google Scholar] [CrossRef]
- da Costa, N.; Blackley, R.; Alzuherri, H.; Chang, K.C. Quantifying the temporospatial expression of postnatal porcine skeletal myosin heavy chain genes. J. Histochem. Cytochem. 2002, 50, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, J.; Zhu, R.; Guo, J.; Wu, Y. Effect of myosin heavy chain composition of muscles on meat quality in Laiwu pigs and Duroc. Sci. China C Life Sci. 2008, 51, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Men, X.M.; Deng, B.; Tao, X.; Qi, K.K.; Xu, Z.W. Association analysis of myosin heavy-chain genes mRNA transcription with the corresponding proteins expression of longissimus muscle in growing pigs. Asian-Aust. J. Anim. Sci. 2016, 29, 457–463. [Google Scholar] [CrossRef]
- Men, X.M.; Deng, B.; Xu, Z.W.; Tao, X.; Qi, K.K. Age-related changes and nutritional regulation of myosin heavy-chain composition in longissimus dorsi of commercial pigs. Animal 2013, 7, 1486–1492. [Google Scholar] [CrossRef]
- Ruusunen, M.; Puolanne, E. Histochemical properties of fibre types in muscles of wild and domestic pigs and the effect of growth rate on muscle fibre properties. Meat Sci. 2004, 67, 533–539. [Google Scholar] [CrossRef]
- Rehfeldt, C.; Te Pas, M.F.; Wimmers, K.; Brameld, J.M.; Nissen, P.M.; Berri, C.; Valente, L.M.; Power, D.M.; Picard, B.; Stickland, N.C.; et al. Advances in research on the prenatal development of skeletal muscle in animals in relation to the quality of muscle-based food. I. Regulation of myogenesis and environmental impact. Animal 2011, 5, 703–717. [Google Scholar] [CrossRef]
- Lefaucheur, L.; Ecolan, P.; Lossec, G.; Gabillard, J.C.; Butler-Browne, G.S.; Herpin, P. Influence of early postnatal cold exposure on myofiber maturation in pig skeletal muscle. J. Muscle Res. Cell Motil. 2001, 22, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef]
- Bee, G. Effect of early gestation feeding, birth weight, and gender of progeny on muscle fiber characteristics of pigs at slaughter. J. Anim. Sci. 2004, 82, 826–836. [Google Scholar] [CrossRef] [PubMed]
- Lefaucheur, L.; Ecolan, P.; Barzic, Y.M.; Marion, J.; Le Dividich, J. Early postnatal food intake alters myofiber maturation in pig skeletal muscle. J. Nutr. 2003, 133, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.T.; Kim, G.D.; Hwang, Y.H.; Ryu, Y.C. Control of fresh meat quality through manipulation of muscle fiber characteristics. Meat Sci. 2013, 95, 828–836. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| MyHC Isoform | Gene | Primer Sequence (5′→ 3′) | Amplicon Length |
| embryonal | MyHCembry | F: CGGGTCCTTCCCATCTGA R: GCAGCAGCCGTGAGAAATC P: FAM-CTGCCCGGCTTTGGTC | 75 |
| I | MyHC-I | F: CGTGCTCCGTCTTCTTTCCTT R: GAAGAAAGAGGCTCAAGCTGGAA P: FAM-CTGCTCTCAGGCCCC | 60 |
| 2a | MyHC-2a | F: GTCCTGCTTTAAAAAGCTCCAAGAA R: AGAGGTCCCTCTTAGCAAGTGA P: FAM-CAGGCTGCATCTTC | 73 |
| 2b | MyHC-2b | F: AACACTTTAAGTAGTTGTCTGCCTTGA R: AGGGCACTAGATGTACCTCTTATGT P: FAM-CCTGCCACCGTCTTC | 76 |
| 2x | MyHC-2x | F: GCTTCAAGTTCTGCCCCACTT R: TGGCAGCCCAGTCAAAGAC P: FAM-CTTTGAGATGCAACCTTG | 59 |
| Full Gene Name | Gene | Amplicon Length | Assay ID |
| Peroxisome proliferator-activated receptor gamma, coactivator 1 alpha | PGC-1α | 96 | Ss03393114_u1 |
| Peroxisome proliferator-activated receptor gamma | PPARγ | 72 | Ss03394829_m1 |
| Lipoprotein-lipase | LPL | 66 | Ss03394612_m1 |
| Carnitine palmitoyltransferase 1B | CPT1B | 60 | Ss03378792_u1 |
| Eukaryotic ribosomal (r) 18S rRNA | 18S rRNA | 69 | Hs03003631_g1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrecl, M.; Cotman, M.; Uršič, M.; Čandek-Potokar, M.; Fazarinc, G. Age-Dependent Expression of MyHC Isoforms and Lipid Metabolism-Related Genes in the Longissimus Dorsi Muscle of Wild and Domestic Pigs. Animals 2019, 9, 10. https://doi.org/10.3390/ani9010010
Vrecl M, Cotman M, Uršič M, Čandek-Potokar M, Fazarinc G. Age-Dependent Expression of MyHC Isoforms and Lipid Metabolism-Related Genes in the Longissimus Dorsi Muscle of Wild and Domestic Pigs. Animals. 2019; 9(1):10. https://doi.org/10.3390/ani9010010
Chicago/Turabian StyleVrecl, Milka, Marko Cotman, Matjaž Uršič, Marjeta Čandek-Potokar, and Gregor Fazarinc. 2019. "Age-Dependent Expression of MyHC Isoforms and Lipid Metabolism-Related Genes in the Longissimus Dorsi Muscle of Wild and Domestic Pigs" Animals 9, no. 1: 10. https://doi.org/10.3390/ani9010010
APA StyleVrecl, M., Cotman, M., Uršič, M., Čandek-Potokar, M., & Fazarinc, G. (2019). Age-Dependent Expression of MyHC Isoforms and Lipid Metabolism-Related Genes in the Longissimus Dorsi Muscle of Wild and Domestic Pigs. Animals, 9(1), 10. https://doi.org/10.3390/ani9010010