Behaviour-Based Husbandry—A Holistic Approach to the Management of Abnormal Repetitive Behaviors


Simple Summary
Abstract
1. Introduction
2. Zoo Animal Welfare
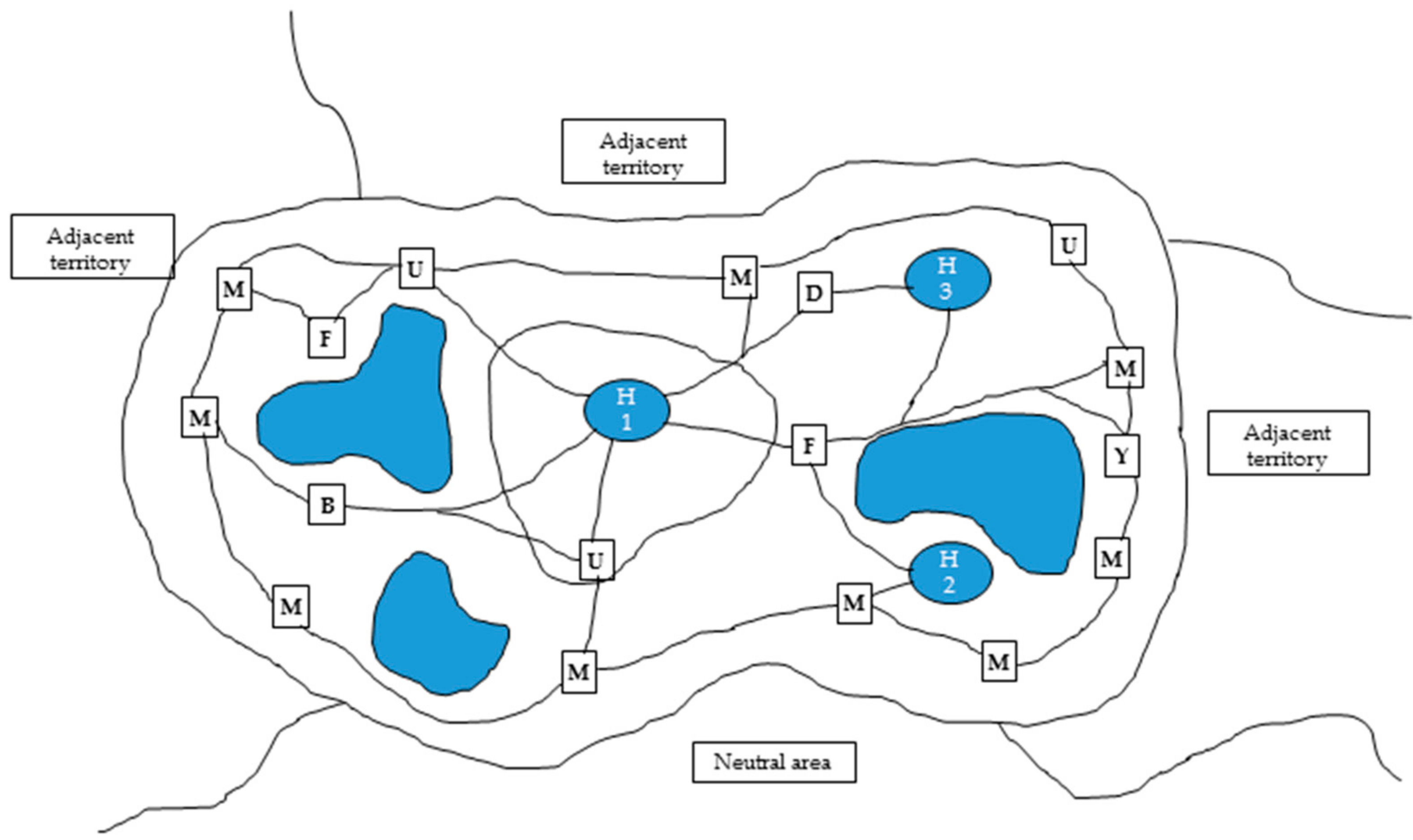
2.1. Zoo Animal Welfare: A Global Perspective
2.2. Zookeepers and the Importance of Husbandry
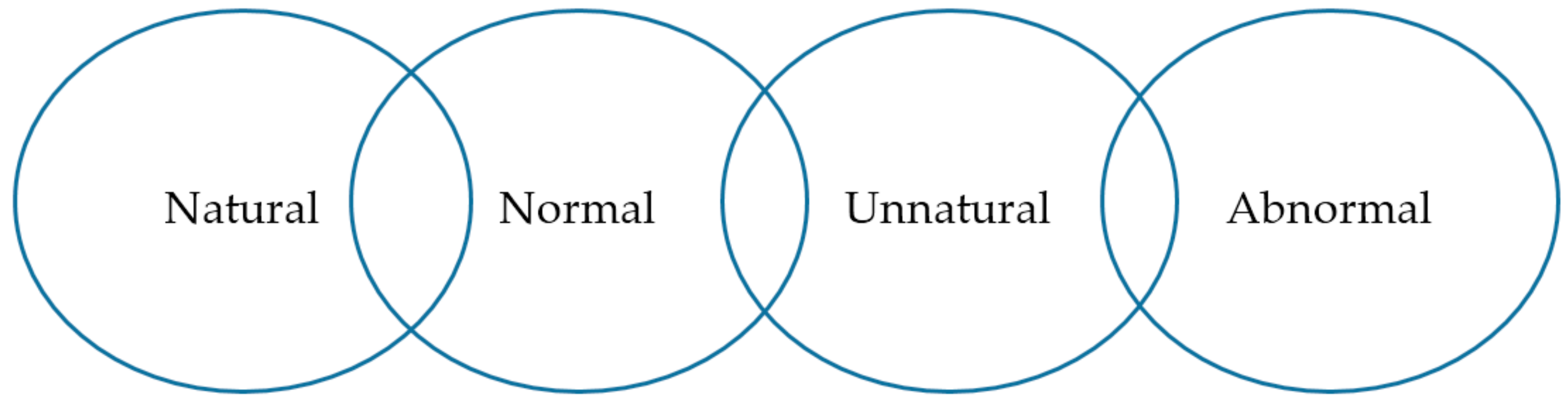
2.3. Back to Basics—The Importance of Behaviour in Zoo Animal Welfare
2.4. Addressing Behaviour Problems: Prevention Is Better than Cure
2.5. Environmental Enrichment—A Universal Panacea?
2.6. Behavioural Pharmacology
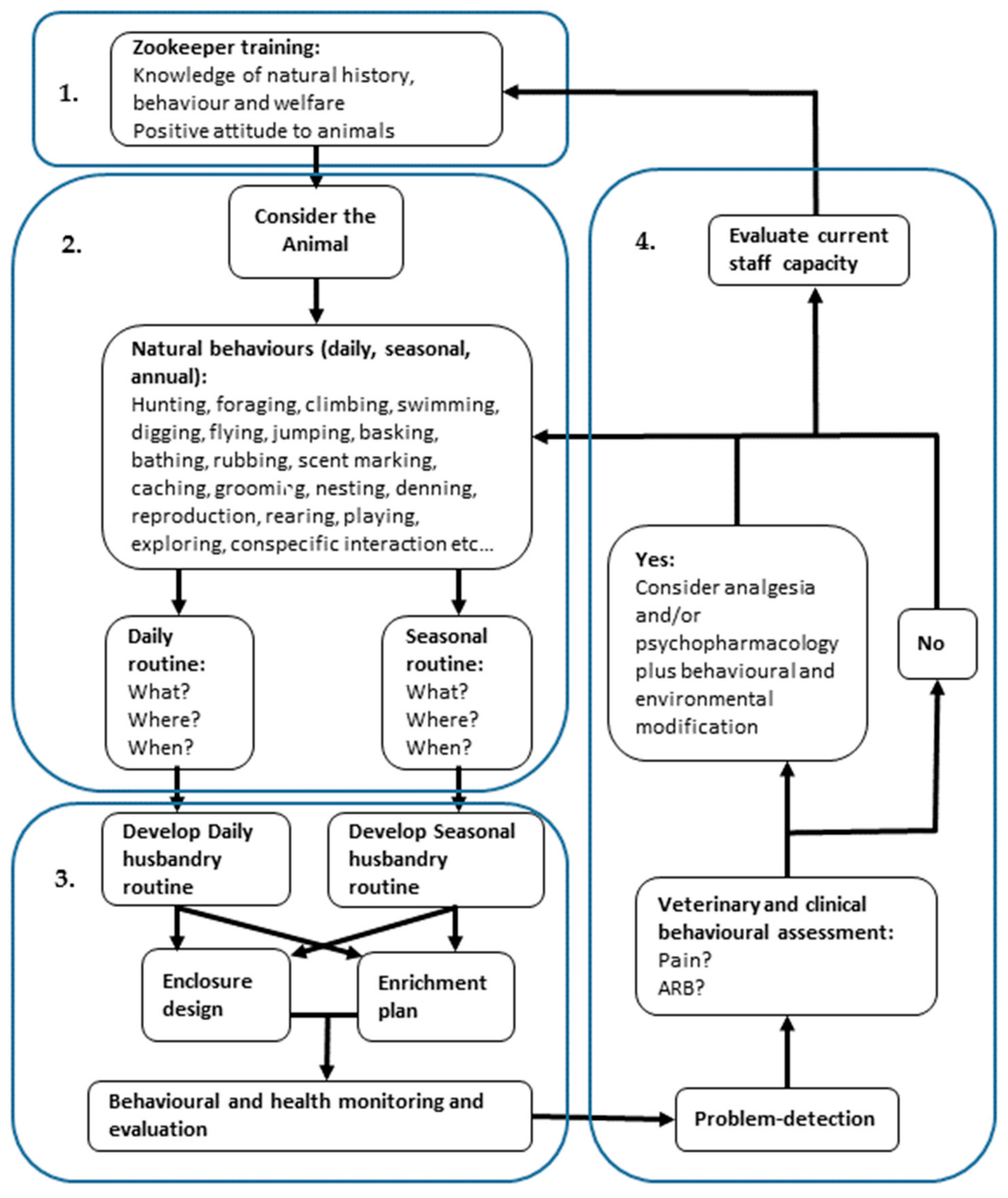
2.7. Framework for Preventing and Managing Behavioural Pathology
3. Conclusions
Funding
Conflicts of Interest
References
- Mellor, D.J.; Hunt, S.; Gusset, M. Caring for Wildlife: The World Zoo and Aquarium animal Welfare Strategy; WAZA Executive Office: Gland, Switzerland, 2015. [Google Scholar]
- Powell, D.M.; Watters, J.V. The evolution of the animal welfare movement in US. Zoos and aquariums. Der Zool. Gart. 2017, 86, 219–234. [Google Scholar] [CrossRef]
- Kagan, R.; Veasey, J. Challenges of Zoo Animal Welfare. In Wild Mammals in Captivity: Principles and Techniques for Zoo Management; University of Chicago Press: Chicago, IL, USA, 2010; pp. 11–21. [Google Scholar]
- Carlstead, K.; Shepherdson, D. Effects of environmental enrichment on reproduction. Zoo Biol. 1994, 13, 447–458. [Google Scholar] [CrossRef]
- Tetley, C.L.; O’Hara, S.J. Ratings of animal personality as a tool for improving the breeding, management and welfare of zoo mammals. Anim. Welf. 2012, 21, 463–476. [Google Scholar] [CrossRef]
- Zhang, G.; Swaisgood, R.R.; Zhang, H. Evaluation of behavioral factors influencing reproductive success and failure in captive giant pandas. Zoo Biol. 2004, 23, 15–31. [Google Scholar] [CrossRef]
- Jule, K.R.; Leaver, L.A.; Lea, S.E.G. The effects of captive experience on reintroduction survival in carnivores: A review and analysis. Biol. Conserv. 2008, 141, 355–363. [Google Scholar] [CrossRef]
- Reading, R.P.; Miller, B.; Shepherdson, D. The value of enrichment to reintroduction success. Zoo Biol. 2013, 32, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.E.; Nash, S.M.; Riley, L.M. To pace or not to pace? A review of what abnormal repetitive behavior tells us about zoo animal management. J. Vet. Behav. Clin. Appl. Res. 2017, 20, 11–21. [Google Scholar] [CrossRef]
- Kroshko, J.; Clubb, R.; Harper, L.; Mellor, E.; Moehrenschlager, A.; Mason, G. Stereotypic route tracing in captive carnivora is predicted by species-typical home range sizes and hunting styles. Anim. Behav. 2016, 117, 197–209. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; Shepherdson, D.J. Scientific approaches to enrichment and stereotypies in zoo animals: What’s been done and where should we go next? Zoo Biol. 2005, 24, 499–518. [Google Scholar] [CrossRef]
- World Association of Zoos and Aquariums. Building a Future for Wildlife: The World Zoo and Aquarium Conservation Strategy; Waza Executive Office: Gland, Switzerland, 2005. [Google Scholar]
- Catibog-Sinha, C. Zoo tourism: Biodiversity conservation through tourism. J. Ecotour. 2008, 7, 160–178. [Google Scholar] [CrossRef]
- Kagan, R.; Carter, S.; Allard, S. A universal animal welfare framework for zoos. J. Appl. Anim. Welf. Sci. 2015, 18, S1–S10. [Google Scholar] [CrossRef] [PubMed]
- Blackett, T.A.; McKenna, C.; Kavanagh, L.; Morgan, D.R. The welfare of wild animals in zoological institutions: Are we meeting our duty of care? Int. Zoo Yearb. 2017, 51, 187–202. [Google Scholar] [CrossRef]
- Melfi, V.; Hosey, G. Capacity building for better animal welfare. Int. Zoo Yearb. 2011, 45, 274–281. [Google Scholar] [CrossRef]
- Melfi, V.A. There are big gaps in our knowledge, and thus approach, to zoo animal welfare: A case for evidence-based zoo animal management. Zoo Biol. 2009, 28, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Goulart, V.D.; Azevedo, P.G.; Schepop, J.A.V.D.; Teixeira, C.P.; Barçante, L.; Azevedo, C.S.; Young, R.J. Gaps in the study of zoo and wild animal welfare. Zoo Biol. 2009, 28, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.P.; Broom, D.M. Measuring zoo animal welfare: Theory and practice. Zoo Biol. 2009, 28, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Wickins-Dražilová, D. Zoo animal welfare. J. Agric. Environ. Ethics 2006, 19, 27–36. [Google Scholar] [CrossRef]
- Waugh, D.R.; Wemmer, C. Training in zoo biology: Two approaches to enhance the conservation role of zoos in the tropics. In Creative Conservation: Interactive Management of Wild and Captive Animals; Olney, P.J.S., Mace, G.M., Feistner, A.T.C., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 207–240. [Google Scholar]
- BIAZA. Available online: https://biaza.org.uk/downloader/78 (accessed on 10 May 2018).
- Hosey, G.; Melfi, V. Are we ignoring neutral and negative human–animal relationships in zoos? Zoo Biol. 2015, 34, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.J.; Melfi, V. Keeper-animal interactions: Differences between the behaviour of zoo animals affect stockmanship. PLoS ONE 2015, 10, e0140237. [Google Scholar] [CrossRef] [PubMed]
- Alligood, C.; Leighty, K. Putting the “e” in spider: Evolving trends in the evaluation of environmental enrichment efficacy in zoological settings. Anim. Behav. Cognit. 2015, 2, 200–217. [Google Scholar] [CrossRef]
- Ross, S.R. Issues of choice and control in the behaviour of a pair of captive polar bears (ursus maritimus). Behav. Processes 2006, 73, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Owen, M.A.; Swaisgood, R.R.; Czekala, N.M.; Lindburg, D.G. Enclosure choice and well-being in giant pandas: Is it all about control? Zoo Biol. 2005, 24, 475–481. [Google Scholar] [CrossRef]
- Eilam, D.; Zor, R.; Szechtman, H.; Hermesh, H. Rituals, stereotypy and compulsive behavior in animals and humans. Neurosci. Biobehav. Rev. 2006, 30, 456–471. [Google Scholar] [CrossRef] [PubMed]
- Hediger, H. Wild Animals in Captivity. Dover Publications: New York, NY, USA, 1964. [Google Scholar]
- Réale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Integrating animal temperament within ecology and evolution. Biol. Rev. 2007, 82, 291–318. [Google Scholar] [CrossRef] [PubMed]
- Sergiel, A.; Naves, J.; Kujawski, P.; Maślak, R.; Serwa, E.; Ramos, D.; Fernández-Gil, A.; Revilla, E.; Zwijacz-Kozica, T.; Zięba, F.; et al. Histological, chemical and behavioural evidence of pedal communication in brown bears. Sci. Rep. 2017, 7, 1052. [Google Scholar] [CrossRef] [PubMed]
- Tattoni, C.; Bragalanti, N.; Groff, C.; Rovero, F. Patterns in the use of rub trees by the eurasian brown bear. Hystrix Ital. J. Mammal. 2015, 26, 118–124. [Google Scholar]
- Clapham, M.; Kitchin, J. Social play in wild brown bears of varying age-sex class. Acta Ethol. 2016, 19, 181–188. [Google Scholar] [CrossRef]
- Horner, M.A.; Powell, R.A. Internal structure of home ranges of black bears and analyses of home-range overlap. J. Mammal. 1990, 71, 402–410. [Google Scholar] [CrossRef]
- Penteriani, V.; López-Bao, J.V.; Bettega, C.; Dalerum, F.; Delgado, M.D.M.; Jerina, K.; Kojola, I.; Krofel, M.; Ordiz, A. Consequences of brown bear viewing tourism: A review. Biol. Conserv. 2017, 206, 169–180. [Google Scholar] [CrossRef]
- Maślak, R.; Sergiel, A.; Bowles, D.; Paśko, Ł. The welfare of bears in zoos: A case study of poland. J. Appl. Anim. Welf. Sci. 2016, 19, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.; Wilder, J.; Wilson, R.R. Polar bear–grizzly bear interactions during the autumn open-water period in alaska. J. Mammal. 2015, 96, 1317–1325. [Google Scholar] [CrossRef]
- Shepherdson, D.; Lewis, K.D.; Carlstead, K.; Bauman, J.; Perrin, N. Individual and environmental factors associated with stereotypic behavior and fecal glucocorticoid metabolite levels in zoo housed polar bears. Appl. Anim. Behav. Sci. 2013, 147, 268–277. [Google Scholar] [CrossRef]
- Heesen, R.; Genty, E.; Rossano, F.; Zuberbühler, K.; Bangerter, A. Social play as joint action: A framework to study the evolution of shared intentionality as an interactional achievement. Learn. Behav. 2017, 45, 390–405. [Google Scholar] [CrossRef] [PubMed]
- Mattiello, S.; Brignoli, S.M.; Cordedda, A.; Pedroni, B.; Colombo, C.; Rosi, F. Effect of the change of social environment on the behavior of a captive brown bear (Ursus arctos). J. Vet. Behav. Clin. Appl. Res. 2014, 9, 119–123. [Google Scholar] [CrossRef]
- Lührs, M.-L.; Dammhahn, M. An unusual case of cooperative hunting in a solitary carnivore. J. Ethol. 2010, 28, 379–383. [Google Scholar] [CrossRef]
- Hull, V.; Zhang, J.; Zhou, S.; Huang, J.; Li, R.; Liu, D.; Xu, W.; Huang, Y.; Ouyang, Z.; Zhang, H.; et al. Space use by endangered giant pandas. J. Mammal. 2015, 96, 230–236. [Google Scholar] [CrossRef]
- Winter, A.; Cooper, J.J. Observational Study on the Effects of Enclosure Size and Design on Diurnal Activity and Species-Specific Behaviours in Captive Polar Bear (Ursus maritimus); University of Lincoln: Lincoln, UK, 2015. [Google Scholar]
- Woods, K.; Cooper, J.J. Effects of Seasonal Changes and Climate on the Behaviour of the Polar Bear (Ursus maritimus) Population at the Yorkshire Wildlife Park; University of Lincoln: Lincoln, UK, 2017. [Google Scholar]
- Mellor, D.; Beausoleil, N. Extending the ‘five domains’ model for animal welfare assessment to incorporate positive welfare states. Anim. Welf. 2015, 24, 241–253. [Google Scholar] [CrossRef]
- Bashaw, M.J.; Gibson, M.D.; Schowe, D.M.; Kucher, A.S. Does enrichment improve reptile welfare? Leopard geckos (Eublepharis macularius) respond to five types of environmental enrichment. Appl. Anim. Behav. Sci. 2016, 184, 150–160. [Google Scholar] [CrossRef]
- Cordero-Rivera, A. Behavioral diversity (ethodiversity): A neglected level in the study of biodiversity. Front. Ecol. Evolut. 2017, 5. [Google Scholar] [CrossRef]
- Kirkwood, J.K. Welfare, husbandry and veterinary care of wild animals in captivity: Changes in attitudes, progress in knowledge and techniques. Int. Zoo Yearb. 2003, 38, 124–130. [Google Scholar] [CrossRef]
- Poole, T.B. Normal and abnormal behavior in captive primates. Primate Rep. 1988, 22, 3–11. [Google Scholar]
- Polverino, G.; Manciocco, A.; Vitale, A.; Alleva, E. Stereotypic behaviours in Melopsittacus undulatus: Behavioural consequences of social and spatial limitations. Appl. Anim. Behav. Sci. 2015, 165, 143–155. [Google Scholar] [CrossRef]
- Schuppli, C.; Fraser, D.; Bacon, H. Welfare of non-traditional pets. Rev. Sci. Tech. 2014, 33, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Martínez Silvestre, A. How to assess stress in reptiles. J. Exotic Pet Med. 2014, 23, 240–243. [Google Scholar] [CrossRef]
- Crane, A.L.; Ferrari, M.C.O. Learning of safety by a social fish: Applications for studying post-traumatic stress in humans. Anim. Behav. 2017, 132, 271–279. [Google Scholar] [CrossRef]
- Greenway, E.; Jones, K.S.; Cooke, G.M. Environmental enrichment in captive juvenile thornback rays, Raja clavata (linnaeus 1758). Appl. Anim. Behav. Sci. 2016, 182, 86–93. [Google Scholar] [CrossRef]
- Mason, G.J. Stereotypic behaviour in captive animals: Fundamentals and applications to welfare In Stereotypies in Captive Animals; Mason, G., Rushen, J., Eds.; CAB International: Wallingford, UK, 2006; pp. 325–356. [Google Scholar]
- Rutherford, K.M.D.; Piastowska-Ciesielska, A.; Donald, R.D.; Robson, S.K.; Ison, S.H.; Jarvis, S.; Brunton, P.J.; Russell, J.A.; Lawrence, A.B. Prenatal stress produces anxiety prone female offspring and impaired maternal behaviour in the domestic pig. Physiol. Behav. 2014, 129, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, M.S.; Poppe, T.T.; McCormick, M.; Damsgård, B.; Salte, R.; Braastad, B.O.; Bakken, M. Simulated maternal pre-spawning stress affects offspring’s attributes in farmed atlantic salmon salmo salar (linnaeus, 1758). Aquac. Res. 2015, 46, 1480–1489. [Google Scholar] [CrossRef]
- Panksepp, J. The basic emotional circuits of mammalian brains: Do animals have affective lives? Neurosci. Biobehav. Rev. 2011, 35, 1791–1804. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.J.; Tamborski, M.A.; Pickens, S.R.; Timberlake, W. Animal and visitor interactions in the modern zoo: Conflicts and interventions. Appl. Anim. Behav. Sci. 2009, 120, 1–8. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Magrath, M.J.L.; Butler, K.L.; Hemsworth, P.H. Little penguins, eudyptula minor, show increased avoidance, aggression and vigilance in response to zoo visitors. Appl. Anim. Behav. Sci. 2015, 168, 71–76. [Google Scholar] [CrossRef]
- Carlstead, K. A comparative approach to the study of keeper–animal relationships in the zoo. Zoo Biol. 2009, 28, 589–608. [Google Scholar] [CrossRef] [PubMed]
- Maslak, R.; Sergiel, A.; Hill, S.P. Some aspects of locomotory stereotypies in spectacled bears (tremarctos ornatus) and changes in behavior after relocation and dental treatment. J. Vet. Behav. Clin. Appl. Res. 2013, 8, 335–341. [Google Scholar] [CrossRef]
- Mellor, D.J. Positive animal welfare states and encouraging environment-focused and animal-to-animal interactive behaviours. N. Z. Vet. J. 2015, 63, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Latham, N.R.; Mason, G.J. Maternal deprivation and the development of stereotypic behaviour. Appl. Anim. Behav. Sci. 2008, 110, 84–108. [Google Scholar] [CrossRef]
- Wiedenmayer, C. Causation of the ontogenetic development of stereotypic digging in gerbils. Anim. Behav. 1997, 53, 461–470. [Google Scholar] [CrossRef]
- Dawkins, M.S. Behavioural deprivation: A central problem in animal welfare. Appl. Anim. Behav. Sci. 1988, 20, 209–225. [Google Scholar] [CrossRef]
- Hoy, J.M.; Murray, P.J.; Tribe, A. Thirty years later: Enrichment practices for captive mammals. Zoo Biol. 2010, 29, 303–316. [Google Scholar] [CrossRef] [PubMed]
- European Commission. EU Zoos Directive: Good Practices Document; European Commission, Publications Office of the European Union: Luxembourg, 2015. [Google Scholar]
- Cummings, D.; Brown, J.L.; Rodden, M.D.; Songsasen, N. Behavioral and physiologic responses to environmental enrichment in the maned wolf (chrysocyon brachyurus). Zoo Biol. 2007, 26, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Mehrkam, L.R.; Dorey, N.R. Preference assessments in the zoo: Keeper and staff predictions of enrichment preferences across species. Zoo Biol. 2015, 34, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Mellen, J.; MacPhee, M.S. Philosophy of environmental enrichment: Past, present, and future. Zoo Biol. 2001, 20, 211–226. [Google Scholar] [CrossRef]
- Tarou, L.R.; Kuhar, C.W.; Adcock, D.; Bloomsmith, M.A.; Maple, T.L. Computer-assisted enrichment for zoo-housed orangutans (pongo pygmaeus). Anim. Welf. 2004, 13, 445–453. [Google Scholar]
- Perdue Bonnie, M.; Clay Andrea, W.; Gaalema Diann, E.; Maple Terry, L.; Stoinski Tara, S. Technology at the zoo: The influence of a touchscreen computer on orangutans and zoo visitors. Zoo Biol. 2012, 31, 27–39. [Google Scholar] [CrossRef] [PubMed]
- French, F.; Mancini, C.; Sharp, H. High tech cognitive and acoustic enrichment for captive elephants. J. Neurosci. Methods 2018, 300, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Kim-McCormack, N.N.E.; Smith, C.L.; Behie, A.M. Is interactive technology a relevant and effective enrichment for captive great apes? Appl. Anim. Behav. Sci. 2016, 185, 1–8. [Google Scholar] [CrossRef]
- Dawkins, M.S. Behaviour as a tool in the assessment of animal welfare1. Zoology 2003, 106, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Bourne, D.C.; Cracknell, J.M.; Bacon, H.J. Veterinary issues related to bears (ursidae). Int. Zoo Yearb. 2010, 44, 16–32. [Google Scholar] [CrossRef]
- Goldberg, M.E. How to be a pain management advocate for exotic and zoo animals. Vet. Nurse 2017, 8, 389–397. [Google Scholar] [CrossRef]
- Kitchener, A.; Macdonald, A.A. The longevity legacy: The problem of old animals in zoos. Adv. Ethol. 2002, 7–10. [Google Scholar]
- Vachon, P.; Millecamps, M.; Low, L.; Thompsosn, S.J.; Pailleux, F.; Beaudry, F.; Bushnell, C.M.; Stone, L.S. Alleviation of chronic neuropathic pain by environmental enrichment in mice well after the establishment of chronic pain. Behav. Brain Funct. 2013, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Föllmi, J. Symptoms, Radiographic Examinations and Pathologies: Development of a Scoring System to Evaluate Physical Condition and Quality of Life in Geriatric Zoo Mammals. Uitgever Niet Vastgesteld. 2005. Available online: http://www.tierschutz.vetsuisse.unibe.ch/e191756/e224004/e224515/e239752/Diss_Foellmi_ger_eng.pdf (accessed on 27 June 2018).
- Föllmi, J.; Steiger, A.; Walzer, C.; Robert, N.; Geissbühler, U.; Doherr, M.; Wenker, C. A scoring system to evaluate physical condition and quality of life in geriatric zoo mammals. Anim. Welf. 2007, 16, 309–318. [Google Scholar]
- Overall, K.L. Pharmacological treatment in behavioural medicine: The importance of neurochemistry, molecular biology and mechanistic hypotheses. Vet. J. 2001, 162, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Overall, K.L. Proceedings of the dogs trust meeting on advances in veterinary behavioural medicine london; 4th–7th november 2004: Veterinary behavioural medicine: A roadmap for the 21st century. Vet. J. 2005, 169, 130–143. [Google Scholar] [CrossRef]
- Yalcin, E.; Aytug, N. Use of fluoxetine to treat stereotypical pacing behavior in a brown bear (Ursus arctos). J. Vet. Behav. Clin. Appl. Res. 2007, 2, 73–76. [Google Scholar] [CrossRef]
- Van Zeeland, Y. Medication for behavior modification in birds. Vet. Clin. Exot. Anim. Pract. 2018, 21, 115–149. [Google Scholar] [CrossRef] [PubMed]
- Wielebnowski, N. Stress and distress: Evaluating their impact for the well-being of zoo animals. J. Am. Vet. Med. Assoc. 2003, 223, 973–977. [Google Scholar] [CrossRef] [PubMed]
- McAuley, M.T.; Kenny, R.A.; Kirkwood, T.B.; Wilkinson, D.J.; Jones, J.J.; Miller, V.M. A mathematical model of aging-related and cortisol induced hippocampal dysfunction. BMC Neurosci. 2009, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Lowenstine, L.J.; McManamon, R.; Terio, K.A. Comparative pathology of aging great apes: Bonobos, chimpanzees, gorillas, and orangutans. Vet. Pathol. 2015, 53, 250–276. [Google Scholar] [CrossRef] [PubMed]
- Schütt, T.; Helboe, L.; Pedersen, L.Ø.; Waldemar, G.; Berendt, M.; Pedersen, J.T. Dogs with cognitive dysfunction as a spontaneous model for early alzheimer’s disease: A translational study of neuropathological and inflammatory markers. J. Alzheimer's Dis. 2016, 52, 433–449. [Google Scholar] [CrossRef] [PubMed]
- Head, E.; Gunn-Moore, D. Neuropathology of feline dementia. In Canine and Feline Dementia: Molecular Basis, Diagnostics and Therapy; Landsberg, G., Maďari, A., Žilka, N., Eds.; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- European Professional Zookeeper Qualification Framework. Available online: https://www.zookeepers.eu/framework/area-2-animal-management/2-2-animal-behaviour/#2.2.4 (accessed on 10 May 2018).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
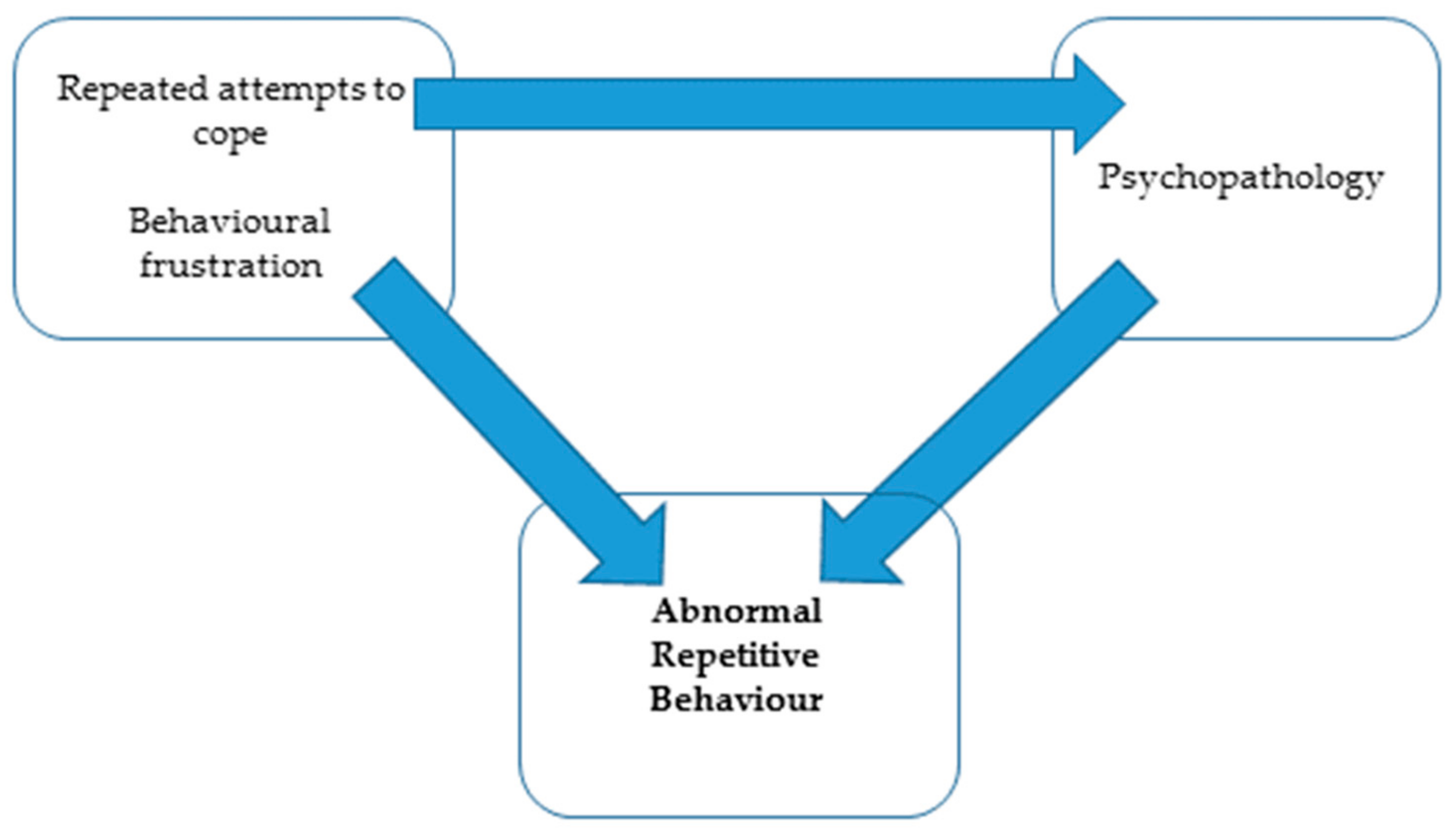
| Potential Triggers | Suggested Mechanism | Aetiology of ARBs [9,55] | Symptom |
|---|---|---|---|
| Negative human–animal interactions e.g., zoo visitors, zoo staff, chronic pain [59,60,61,62]. | The animal does not have the evolved behaviours or physiology to cope with stressors. | Repeated attempts to cope | Abnormal repetitive behaviours, e.g., stereotypy, self-directed behaviours or externally directed behaviours |
| Frustrated reproductive, maternal, foraging, predatory, browsing, grazing behaviours [10,58,63]. | A lack of provisions prevents the animal from performing its evolved behaviours or physiological needs. | Behavioural frustration | |
| Early maternal separation, barren environment during development [56,57,64]. | Impaired cognitive development. | Psychopathology |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bacon, H. Behaviour-Based Husbandry—A Holistic Approach to the Management of Abnormal Repetitive Behaviors. Animals 2018, 8, 103. https://doi.org/10.3390/ani8070103
Bacon H. Behaviour-Based Husbandry—A Holistic Approach to the Management of Abnormal Repetitive Behaviors. Animals. 2018; 8(7):103. https://doi.org/10.3390/ani8070103
Chicago/Turabian StyleBacon, Heather. 2018. "Behaviour-Based Husbandry—A Holistic Approach to the Management of Abnormal Repetitive Behaviors" Animals 8, no. 7: 103. https://doi.org/10.3390/ani8070103
APA StyleBacon, H. (2018). Behaviour-Based Husbandry—A Holistic Approach to the Management of Abnormal Repetitive Behaviors. Animals, 8(7), 103. https://doi.org/10.3390/ani8070103