
Dingoes at the Doorstep: Home Range Sizes and Activity Patterns of Dingoes and Other Wild Dogs around Urban Areas of North-Eastern Australia

Abstract
:Simple summary
Abstract
1. Introduction
2. Methods
2.1. Ethics Statement
2.2. Study Area
2.3. Wild Dog Capture and Tracking
2.4. GPS Data Screening and Filtering
2.5. Home Range Size and Activity Patterns
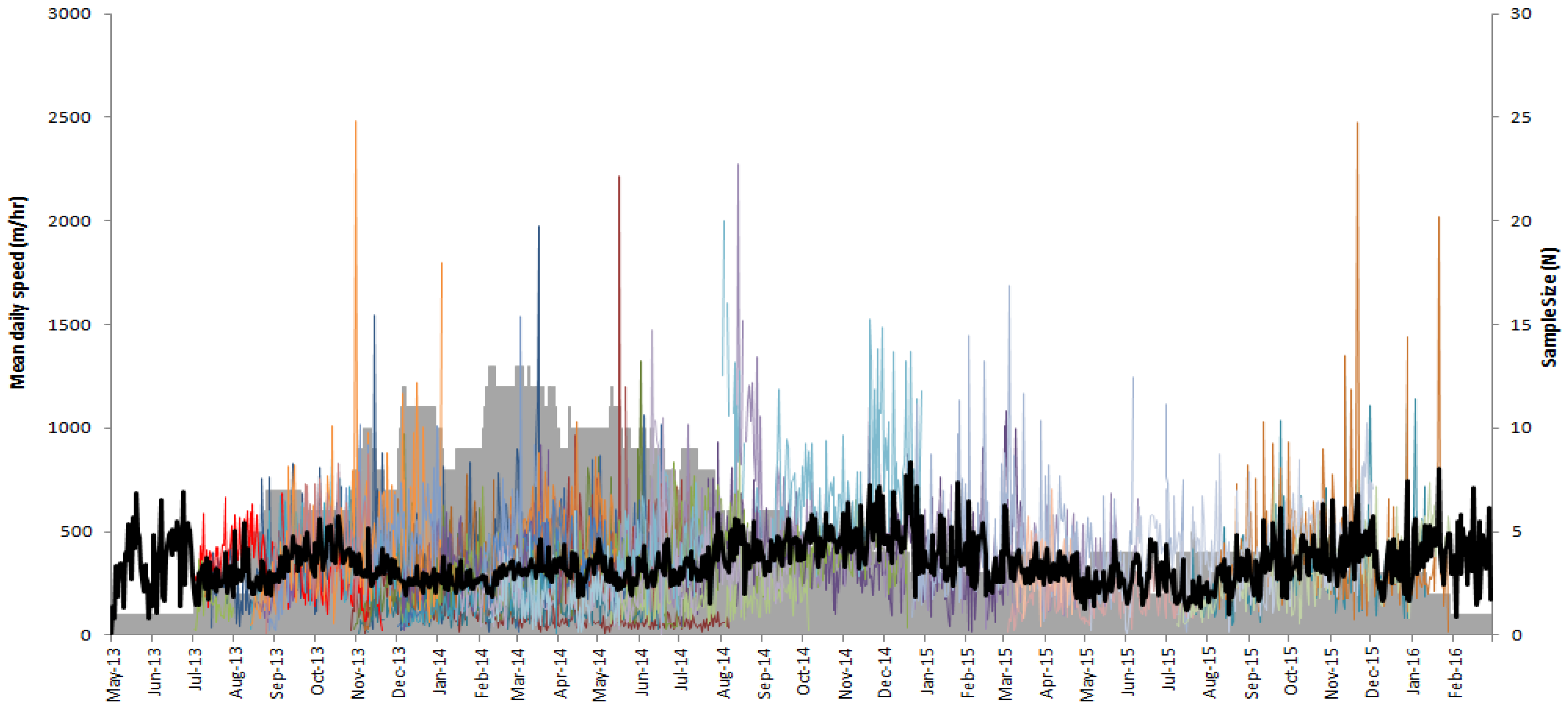
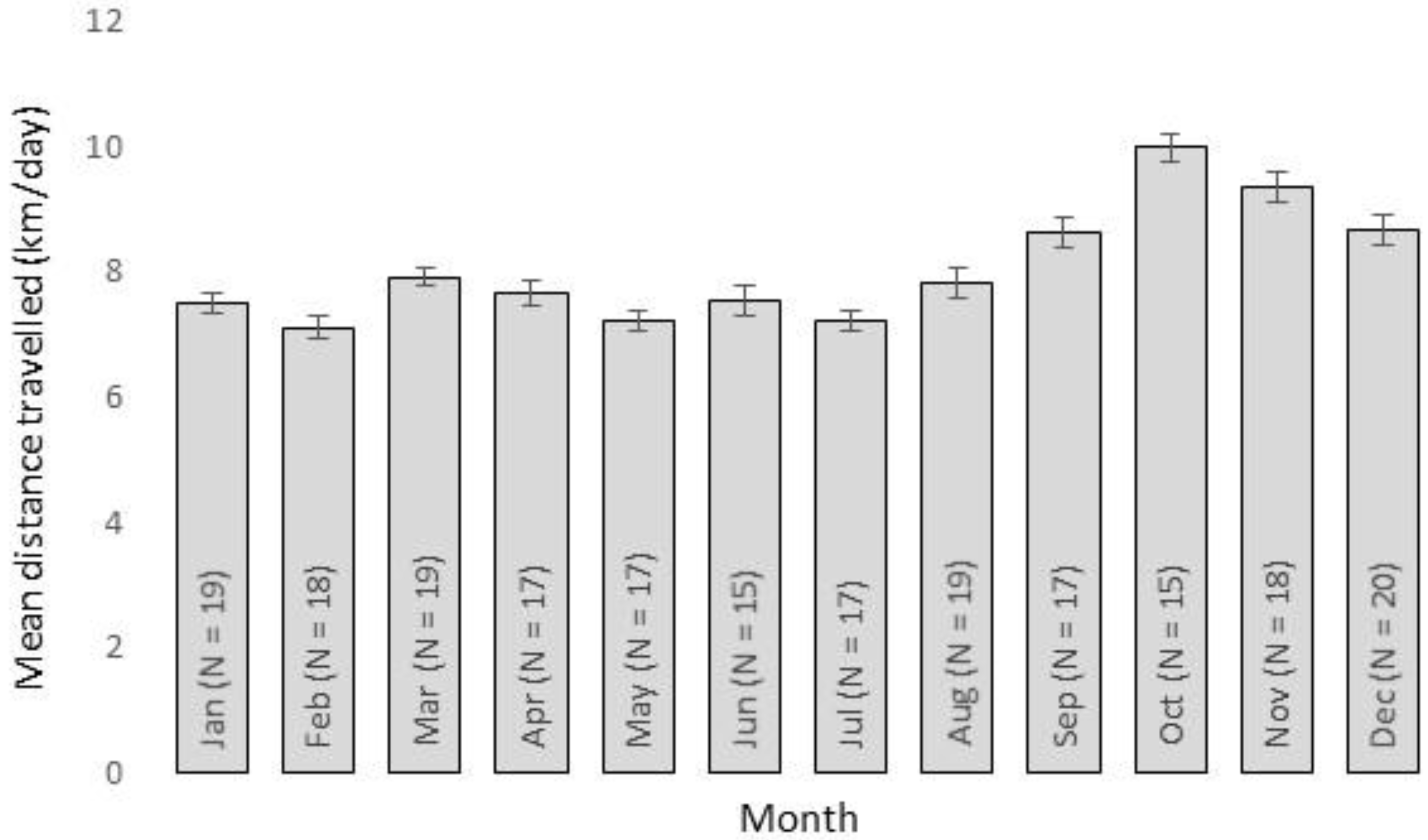
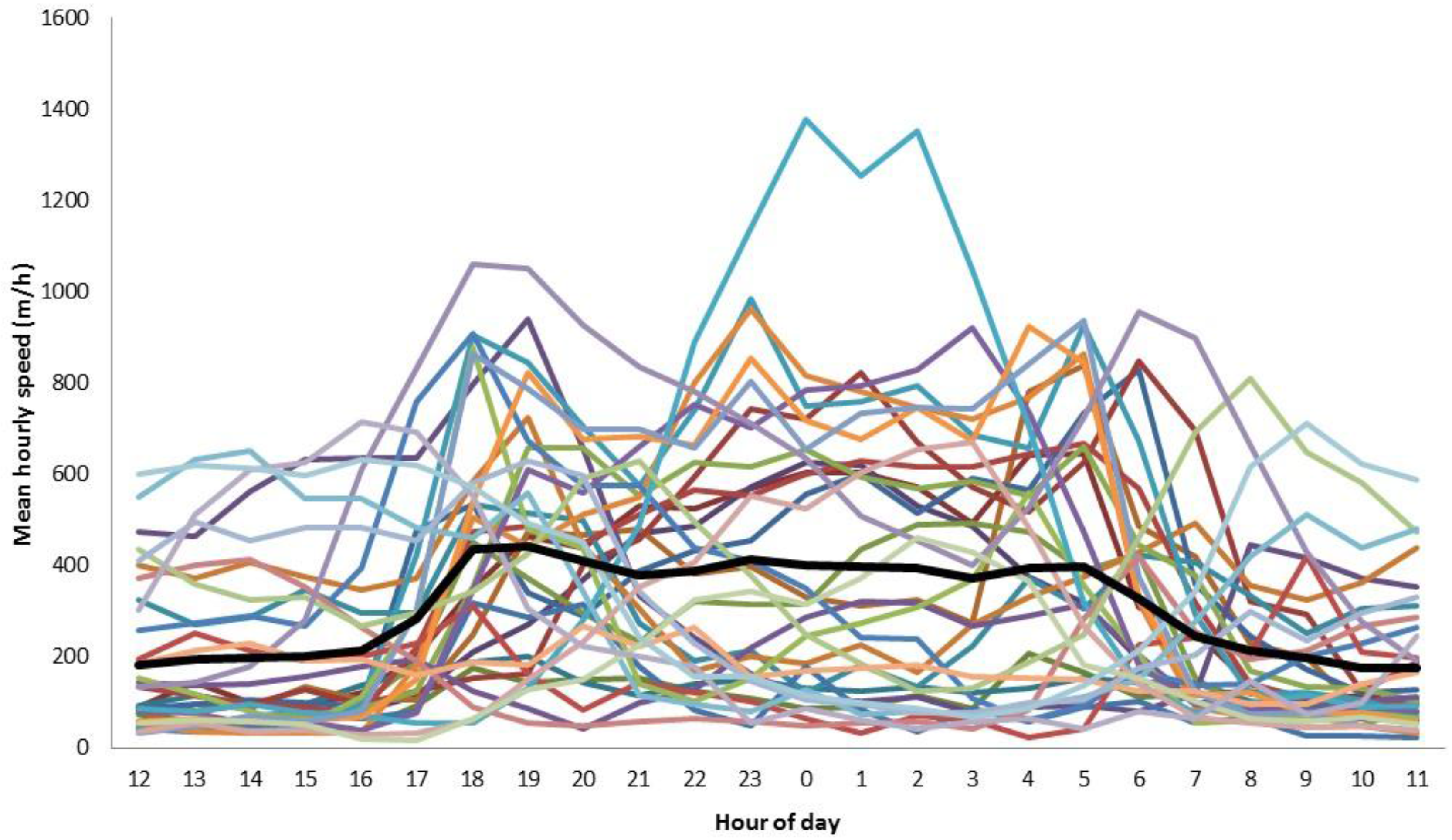
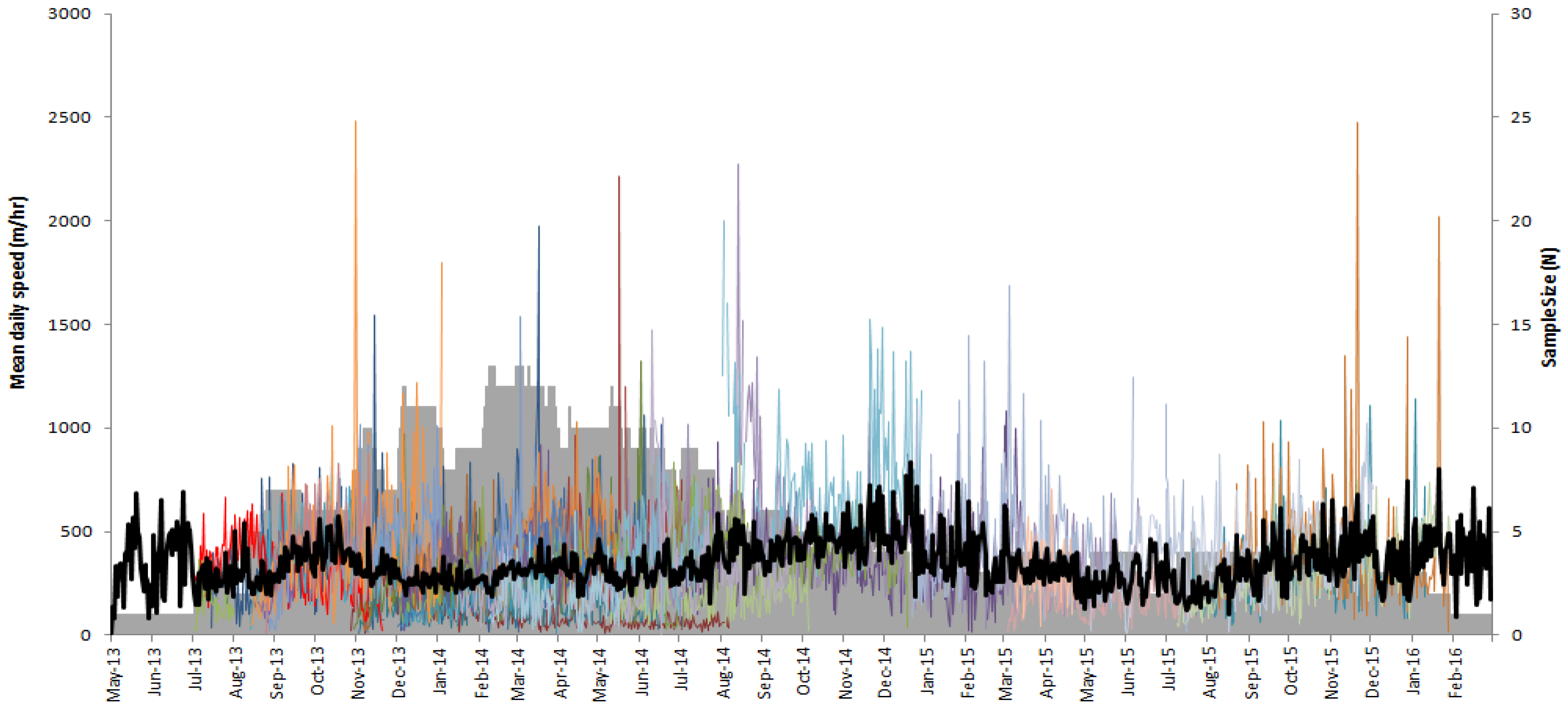
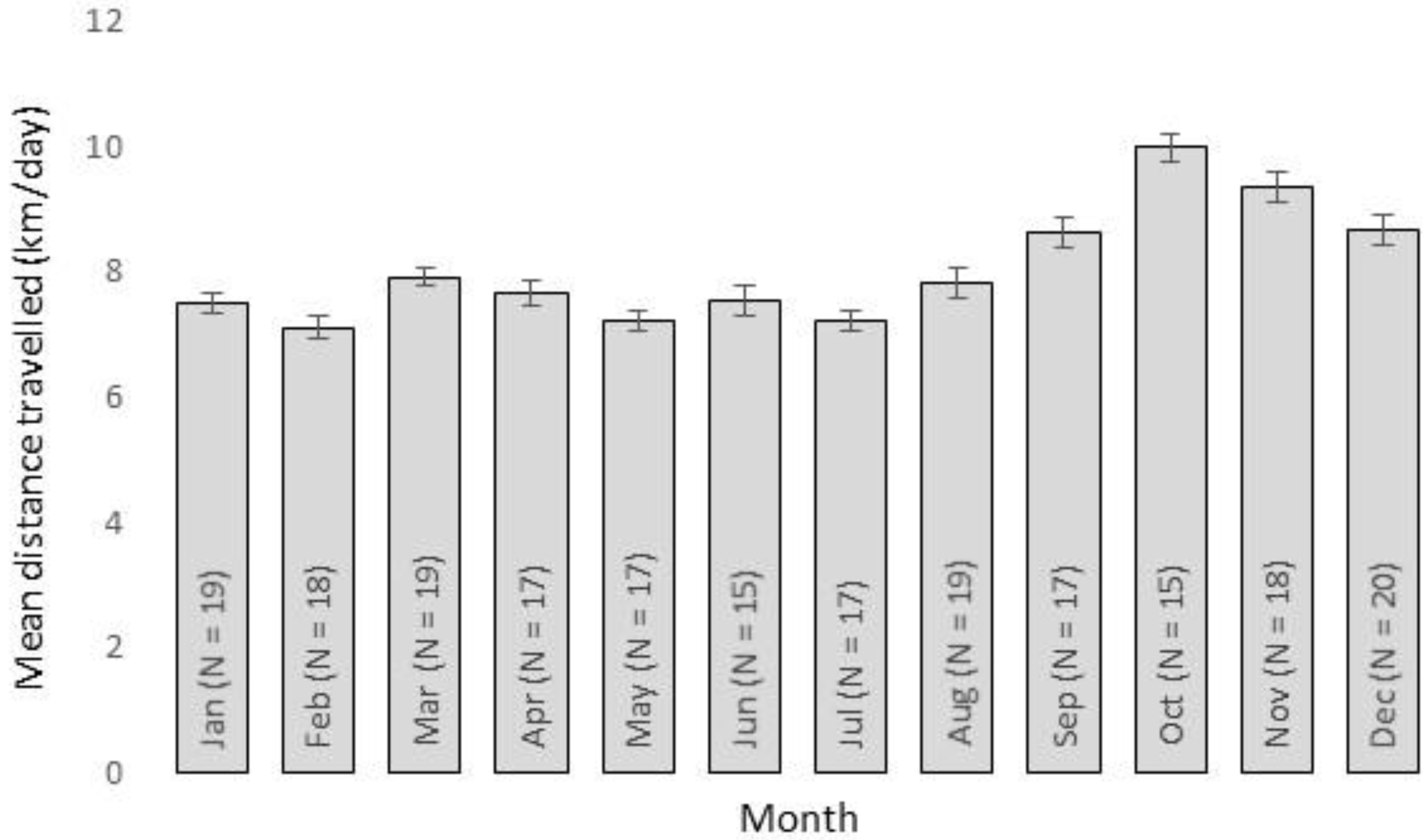
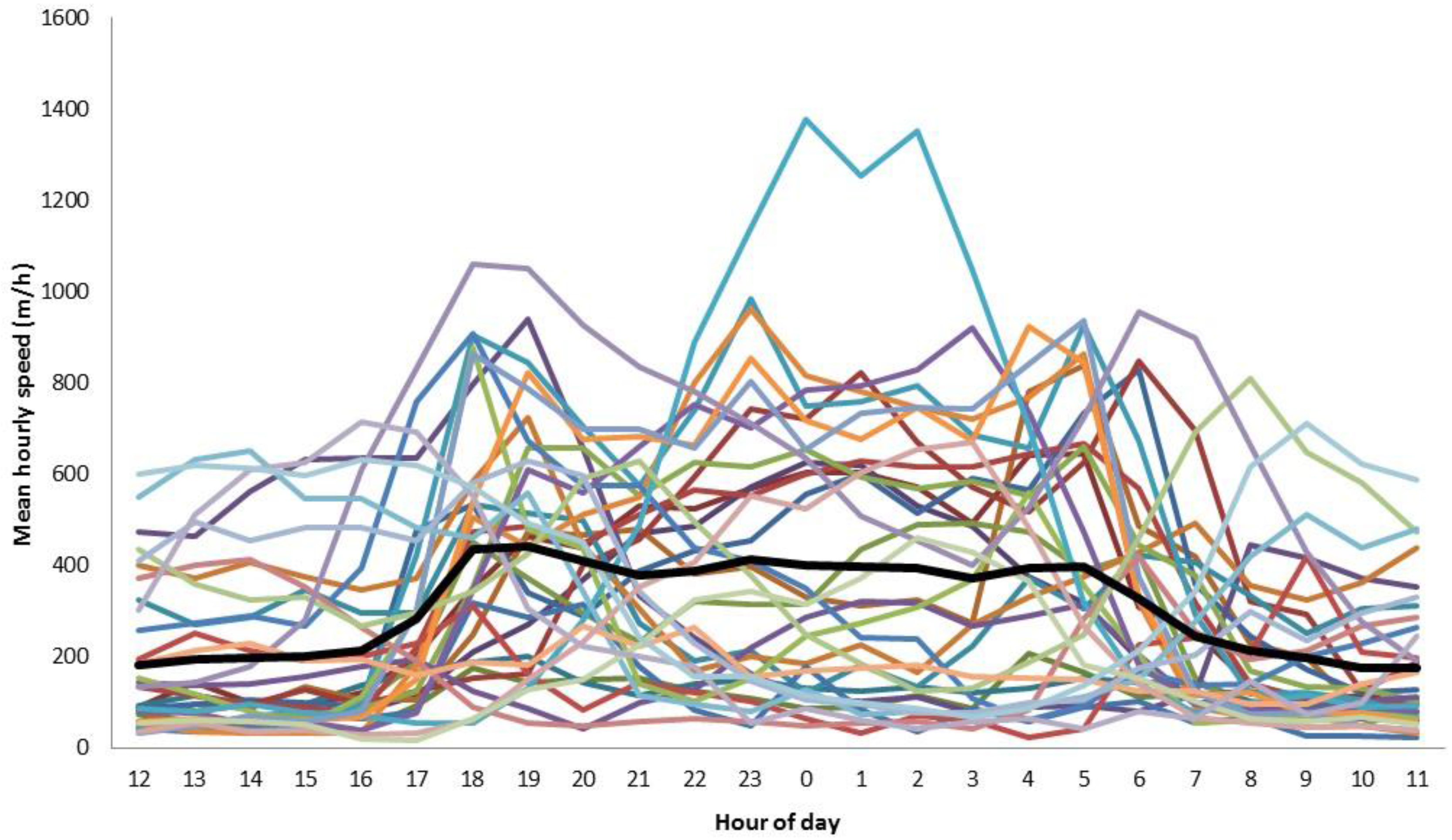
3. Results
4. Discussion
5. Management Conclusions
Supplementary Materials
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Urban Carnivores: Ecology, Conflict, and Conservation; Gehrt, S.D.; Riley, S.P.D.; Cypher, B.L. (Eds.) John Hopkins University Press: Baltimore, MD, USA, 2010.
- Bateman, P.W.; Fleming, P.A. Big city life: Carnivores in urban environments. J. Zool. 2012, 287, 1–23. [Google Scholar] [CrossRef]
- Baker, R.O.; Timm, R.M. Management of conflicts between urban coyotes and humans in southern California. In Proceedings of the 18th Vertebrate Pest Conference, Costa Mesa, CA, USA, 2–5 March 1998; Baker, R.O., Crabb, A.C., Eds.; University of California: Davis, CA, USA, 1998; pp. 299–312. [Google Scholar]
- Berger, K.M. Carnivore-livestock conflicts: Effects of subsidized predator control and economic correlates on the sheep industry. Conserv. Biol. 2006, 20, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Poessel, S.A.; Breck, S.W.; Teel, T.L.; Shwiff, S.; Crooks, K.R.; Angeloni, L. Patterns of human–coyote conflicts in the Denver Metropolitan Area. J. Wildl. Manag. 2013, 77, 297–305. [Google Scholar] [CrossRef]
- Fukuda, Y.; Manolis, C.; Appel, K. Management of human-crocodile conflict in the Northern Territory, Australia: Review of crocodile attacks and removal of problem crocodiles. J. Wildl. Manag. 2014, 78, 1239–1249. [Google Scholar] [CrossRef]
- Adkins, C.A.; Stott, P. Home ranges, movements and habitat associations of red foxes Vulpes vulpes in suburban Toronto, Ontario, Canada. J. Zool. 1998, 244, 335–346. [Google Scholar] [CrossRef]
- Atwood, T.C.; Weeks, H.P.; Gehring, T.M. Spatial ecology of coyotes along a suburban-to-rural gradient. J. Wildl. Manag. 2004, 68, 1000–1009. [Google Scholar] [CrossRef]
- Serieys, L.E.; Lea, A.; Pollinger, J.P.; Riley, S.P.; Wayne, R.K. Disease and freeways drive genetic change in urban bobcat populations. Evol. Appl. 2015, 8, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Denny, E.A.; Dickman, C.R. Review of Cat Ecology and Management Strategies in Australia; Invasive Animals Cooperative Research Centre: Canberra, Australia, 2010. [Google Scholar]
- Allen, B.L.; Goullet, M.; Allen, L.R.; Lisle, A.; Leung, L.K.-P. Dingoes at the doorstep: preliminary data on the ecology of dingoes in urban areas. Landsc. Urban Plan. 2013, 119, 131–135. [Google Scholar] [CrossRef]
- WoolProducers. National Wild Dog Action Plan: Promoting and Supporting Community-Driven Action for landscape-Scale Wild Dog Management; WoolProducers Australia: Barton, Australia, 2014. [Google Scholar]
- Allen, B.L.; Engeman, R.M.; Allen, L.R. Wild dogma II: The role and implications of wild dogma for wild dog management in Australia. Curr. Zool. 2011, 57, 737–740. [Google Scholar] [CrossRef] [Green Version]
- O’Keeffe, M.S.; Walton, C.S. Vertebrate Pests of Built-Up Areas in Queensland; Queensland Department of Natural Resources and Mines: Brisbane, Australia, 2001.
- Jenkins, D.J.; Allen, L.; Goullet, M. Encroachment of Echinococcus granulosus into urban areas in eastern Queensland, Australia. Aust. Vet. J. 2008, 86, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.L.; Engeman, R.M.; Leung, L.K. The short-term effects of a routine poisoning campaign on the movements and detectability of a social top-predator. Environ. Sci. Pollut. Res. Int. 2014, 21, 2178–2190. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.L.; Higginbottom, K.; Bracks, J.H.; Davies, N.; Baxter, G.S. Balancing dingo conservation with human safety on Fraser Island: The numerical and demographic effects of humane destruction of dingoes. Australas. J. Environ. Manag. 2015, 22, 197–215. [Google Scholar] [CrossRef]
- DEEDI. Wild Dog Management Strategy 2011–2016; QLD Department of Employment, Economic Development and Innovation, Biosecurity Queensland: Brisbane, Australia, 2011.
- Thompson, L.-J.; Aslin, H.J.; Ecker, S.; Please, P.; Trestrail, C. Social impacts of wild dogs: A review of literature. AWI Ltd.: Canberra, Australia, 2013. [Google Scholar]
- Meek, P.D. The movement, roaming behaviour and home range of free-roaming domestic dogs, Canis lupus familiaris, in coastal New South Wales. Wildl. Res. 1999, 26, 847–855. [Google Scholar] [CrossRef]
- Allen, L.R. Best practice baiting: Dispersal and seasonal movement of wild dogs (Canis lupus familiaris). In Technical Highlights: Invasive Plant and Animal Research 2008–2009; QLD Department of Employment, Economic Development and Innovation: Canberra, Australia, 2009; pp. 61–62. [Google Scholar]
- Marra, P.P.; Cohen, E.B.; Loss, S.R.; Rutter, J.E.; Tonra, C.M. A call for full annual cycle research in animal ecology. Biol. Lett. 2015, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Fleming, P.J.S.; Allen, B.L.; Ballard, G.; Allen, L.R. Wild Dog Ecology, Impacts and Management in Northern Australian Cattle Enterprises: A Review with Recommendations for RD&E Investments; Meat and Livestock Australia: Sydney, Australia, 2012. [Google Scholar]
- Thomson, P.C. The behavioural ecology of dingoes in north-western Australia: IV. Social and spatial organisation, and movements. Wildlife Res. 1992, 19, 543–563. [Google Scholar] [CrossRef]
- Corbett, L.K. The Dingo in Australia and Asia, 2nd ed.; J.B. Books: Marleston, Australia, 2001. [Google Scholar]
- Newsome, T.M. Ecology of the Dingo (Canis lupus dingo) in the Tanami Desert in Relation to Human-Resource Subsidies. Ph.D. Thesis, The University of Sydney, Sydney, Australia, 2011. [Google Scholar]
- Allen, B.L. Do desert dingoes drink daily? Visitation rates at remote waterpoints in the Strzelecki Desert. Aust. Mammal. 2012, 34, 251–256. [Google Scholar] [CrossRef]
- Matthews, A.; Ruykys, L.; Ellis, B.; FitzGibbon, S.; Lunney, D.; Crowther, M.S.; Glen, A.S.; Purcell, B.; Moseby, K.; Stott, J.; et al. The success of GPS collar deployments on mammals in Australia. Aust. Mammal. 2013, 35, 65–83. [Google Scholar] [CrossRef]
- Data East LLC. XTools Pro extension for ArcGIS. 2005. Available online: http://www.xtoolspro.com (accessed on 1 June 2006).
- Beyer, H.L. Hawth’s Analysis Tools for ArcGIS. 2004. Available online: http://www.spatialecology.com/htools (accessed on 1 June 2006).
- Rodgers, A.R.; Carr, A.P. HRE: The Home Range Extension for ArcView; Ontario Ministry of Natural Resources, Centre for Northern Forest Ecosystem Research: Thunder Bay, ON, Canada, 1998. [Google Scholar]
- Kie, J.G.; Matthiopoulos, J.; Fieberg, J.; Powell, R.A.; Cagnacci, F.; Mitchell, M.S.; Gaillard, J.-M.; Moorcroft, P.R. The home-range concept: are traditional estimators still relevant with modern telemetry technology? Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2221–2231. [Google Scholar] [CrossRef] [PubMed]
- Durr, S.; Ward, M. Roaming behaviour and home range estimation of domestic dogs in Aboriginal and Torres Strait Islander communities in northern Australia using four different methods. Prev. Vet. Med. 2014, 117, 340–357. [Google Scholar] [CrossRef] [PubMed]
- Goldingay, R.L. A review of home-range studies on Australian terrestrial vertebrates: Adequacy of studies, testing of hypotheses, and relevance to conservation and international studies. Aust. J. Zool. 2015, 63, 136–146. [Google Scholar] [CrossRef]
- Walter, W.; Onorato, D.; Fischer, J. Is there a single best estimator? Selection of home range estimators using area-under-the-curve. Mov. Ecol. 2015, 3, 10. [Google Scholar] [CrossRef] [PubMed]
- Rowcliffe, M.J.; Carbone, C.; Kays, R.; Kranstauber, B.; Jansen, P.A. Bias in estimating animal travel distance: The effect of sampling frequency. Methods Ecol. Evol. 2012, 3, 653–662. [Google Scholar] [CrossRef]
- Grinder, M.I.; Krausman, P.R. Home range, habitat use, and nocturnal activity of coyotes in an urban environment. J. Wildl. Manag. 2001, 65, 887–898. [Google Scholar] [CrossRef]
- Bandeira de Melo, L.F.; Lima S’abato, M.A.; Vaz Magni, E.M.; Young, R.J.; Coelho, C.M. Secret lives of maned wolves (Chrysocyon brachyurus): As revealed by GPS tracking collars. J. Zool. 2007, 271, 27–36. [Google Scholar] [CrossRef]
- Thomson, P.C. The behavioural ecology of dingoes in north-western Australia. II. Activity patterns, breeding season and pup rearing. Wildl. Res. 1992, 19, 519–530. [Google Scholar] [CrossRef]
- Allen, B.L.; Carmelito, E.; Amos, M.; Goullet, M.S.; Allen, L.R.; Speed, J.; Gentle, M.; Leung, L.K.P. Diet of dingoes and other wild dogs in peri-urban areas of north-eastern Australia. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.L.; Jones, M.E.; Jarman, P.J. Diurnal and nocturnal grouping and foraging behaviors of free-ranging eastern grey kangaroos. Aust. J. Zool. 1995, 43, 519–529. [Google Scholar] [CrossRef]
- Vernes, K.; Dennis, A.; Winter, J. Mammalian diet and broad hunting strategy of the dingo (Canis familiaris dingo) in the wet tropical rain forests of northeastern Australia. Biotropica 2001, 33, 339–345. [Google Scholar] [CrossRef]
- Newsome, T.M.; Ballard, G.; Dickman, C.R.; Fleming, P.J.S.; van de Ven, R. Home range, activity and sociality of a top predator, the dingo: A test of the Resource Dispersion Hypothesis. Ecography 2013, 36, 914–925. [Google Scholar] [CrossRef]
- Nouvellet, P.; Rasmussen, G.S.A.; Macdonald, D.W.; Courchamp, F. Noisy clocks and silent sunrises: Measurement methods of daily activity pattern. J. Zool. 2012, 286, 179–184. [Google Scholar] [CrossRef]
- APVMA. Review Findings for Sodium Monofluoroacetate: The Reconsideration of Registrations of Products Containing Sodium Monofluoroacetate and Approvals of Their Associated Labels, Environmental Assessment; Australian Pesticides and Veterinary Medicines Authority: Canberra, Australia, 2008.

{kind=link}
{kind=link}
{kind=link}
| Ecosystem | Home Range Size km2 (SE or Range) | N | Method | Source |
|---|---|---|---|---|
| Semi-arid tropics | 77.3 (22.1) | 19 | Pooled mean 95% MCP | [24] |
| Arid | 67 (32–126) | 5 | Not stated | [25] |
| Monsoonal | 39 (15–88) | 18 | Not stated | [25] |
| Arid monsoonal | 25 (7–110) | 24 | Not stated | [25] |
| Arid monsoonal | 414.9 (103.5) | 9 | 85% kernel | [26] |
| Arid | 24 (13–32) | 7 | 95% MCP | [27] |
| Arid | 63.5 (16.6–286.3) | 7 | 90% adaptive kernel | [16] |
| Urban | 17.72 (0.37–100.32) | 9 | 90% adaptive kernel | [11] |
| Urban | 17.47 (0.53–66.02)* | 37 | 90% adaptive kernel | Current Study |
| Dog ID | Estimated Age | Sex | Weight | Processing Time | Capture Date | Body Condition Score | N = Days Monitored | N = GPS Points Analysed | Mean HDOP | Home Range Size (km2) | Mean Daily Distance Travelled (km) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| GCDog01 | 36 | M | 21.0 | 23 | 23-Jul-2013 | 4.0 | 248 | 9268 | 1.88 | 12.17/10.13 ^ | 7.44 |
| GCDog02 | 16 | F | 13.7 | 24 | 5-Nov-2013 | 3.0 | 283 | 10,644 | 1.74 | 4.00 | 2.12 |
| GCDog03 | 28 | F | 17.3 | 43 | 7-Nov-2013 | 4.0 | 223 | 8021 | 1.91 | 14.59 | 5.96 |
| GCDog04 | 28 | F | 15.0 | 10 | 9-Nov-13 | 3.5 | 11 | 175 | 2.17 | 7.97 | 1.20 |
| GCDog05 | 52 | F | 19.6 | 15 | 10-Nov-13 | 3.0 | 225 | 8921 | 1.92 | 9.19 | 3.01 |
| GCDog06 | 28 | F | 14.5 | 21 | 14-Nov-2013 | 4.0 | 98 | 2985 | 1.89 | 30.94 | 6.86 |
| GCDog07 | 34 | F | 15.0 | 15 | 8-Mar-2014 | 4.0 | 110 | 4366 | 1.68 | 30.29 | 8.24 |
| GCDog09 | 45 | M | 15.5 | 15 | 2-Apr-2014 | 4.0 | 110 | 4247 | 1.87 | 45.28 | 8.93 |
| GCDog10 | 24 | F | 25.0 | 15 | 25-Jun-2014 | 3.5 | 42 | 1608 | 2.07 | 25.69 | 5.46 |
| GCDog11 | 60 | M | 23.0 | 20 | 9-Jul-2014 | - | 255 | 11,128 | 1.67 | 34.52 | 8.66 |
| GCdog12 | 24 | M | 19.0 | 11 | 28-Jul-2015 | 2.0 | 176 | 5115 | 1.50 | 20.48 | 5.48 |
| GCdog13 | 60 | F | 15.0 | 16 | 28-Jul-2015 | 2.0 | 192 | 5412 | 1.70 | 21.87 | 6.72 |
| NBDog01 | 17 | F | 16.3 | 13 | 10-Dec-2013 | 4.0 | 27 | 1212 | 1.81 | 0.53 | 2.28 |
| NBDog02 | 17 | M | 13.0 | 8 | 10-Dec-2013 | 3.0 | 17 | 764 | 1.71 | 0.26 | 2.50 |
| NBDog03 | 29 | F | 16.5 | 16 | 11-Dec-2013 | 3.5 | 381 | 17,465 | 1.64 | 4.59 | 8.79 |
| NBDog04 | 29 | M | 21.0 | 12 | 12-Dec-2013 | 4.0 | 127 | 5432 | 1.73 | 8.21 | 8.51 |
| NBDog05 | 29 | M | 19.7 | 15 | 13-Dec-2013 | 4.5 | 117 | 4816 | 1.76 | 1.08 | 3.89 |
| NBDog06 | 31 | M | 18.5 | 9 | 12-Feb-2014 | 4.0 | 99 | 4468 | 1.72 | 16.87 | 11.21 |
| NBDog07 | 31 | F | 15.3 | 15 | 14-Feb-2014 | 3.5 | 93 | 4117 | 1.69 | 8.95 | 9.27 |
| SCDog01 | 12 | F | 15.3 | 23 | 9-May-2013 | 3.5 | 203 | 9608 | 1.48 | 7.39 | 8.01 |
| SCDog02 | 12 | F | 13.8 | 18 | 10-Jul-2013 | 3.0 | 29 | 1322 | 1.66 | 9.41 | 3.95 |
| SCDog03 | 25 | F | 17.8 | 25 | 21-Aug-2013 | 3.0 | 37 | 1044 | 1.92 | 4.26 | 3.78 |
| SCDog04 | 37 | M | 19.0 | 16 | 21-Aug-2013 | 4.0 | 116 | 4913 | 1.84 | 13.76 | 8.76 |
| SCDog05 | 25 | F | 14.2 | 12 | 21-Aug-2013 | 3.0 | 144 | 5525 | 1.88 | 39.73 | 9.00 |
| SCDog06 | 42 | M | 15.7 | 15 | 1-Sep-2013 | 4.0 | 193 | 6967 | 1.80 | 21.82 | 9.40 |
| SCDog07 | 14 | F | 16.0 | 16 | 2-Sep-2013 | 4.0 | 81 | 3752 | 1.75 | 13.42 | 10.36 |
| SCDog08 | 10 | F | 21.4 | 150 | 18-May-2014 | 4.0 | 149 | 4857 | 1.83 | 3.21 | 4.05 |
| SCDog09 | 10 | F | 13.6 | 14 | 19-May-2014 | 4.0 | 394 | 16,962 | 1.26 | 24.82 | 9.19 |
| SCDog10 | 22 | F | 14.5 | 16 | 19-May-2014 | 4.0 | 232 | 9298 | 1.62 | 66.02 | 13.19 |
| SCDog11 | 24 | M | 15.0 | 14 | 17-Jul-2014 | 3.0 | 19 | 895 | 1.72 | 89.64 | 4.72 |
| SCDog14 | 5 | F | 13.4 | 20 | 12-Dec-2014 | 3.0 | 229 | 5950 | 1.73 | 13.25 | 6.47 |
| SCDog15 | 21 | M | 13.0 | 45 | 12-Mar-2015 | 4.0 | 156 | 5710 | 1.44 | 31.61 | 3.68 |
| SCDog16 | 60 | F | 18.0 | 45 | 17-Jul-2015 | 4.0 | 235 | 9849 | 1.62 | 15.89 | 7.08 |
| TCDog01 | 30 | F | 16.0 | 28 | 22-Jan-2014 | 3.0 | 123 | 4658 | 1.72 | 3.72 | 4.99 |
| TCDog02 | 7 | M | 16.0 | 28 | 16-Feb-2014 | 3.0 | 226 | 1560 | 1.69 | 3.93 | 3.53 |
| TCDog03 | 9 | F | 13.5 | 18 | 18-Mar-2015 | 3.0 | 42 | 1540 | 1.63 | 4.09 | 5.55 |
| TCDog04 | 21 | F | 17.5 | 25 | 6-May-2015 | 3.5 | 219 | 4956 | 1.42 | 35.64 | 9.37 |
| Total | Mean 27 | 24F/13M | Mean 16.7 | Mean 23 | Mean 3.5 | Mean 153 | Mean 5662 | 1.73 | Mean 17.47 * | Mean 6.86 # |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McNeill, A.T.; Leung, L.K.-P.; Goullet, M.S.; Gentle, M.N.; Allen, B.L. Dingoes at the Doorstep: Home Range Sizes and Activity Patterns of Dingoes and Other Wild Dogs around Urban Areas of North-Eastern Australia. Animals 2016, 6, 48. https://doi.org/10.3390/ani6080048
McNeill AT, Leung LK-P, Goullet MS, Gentle MN, Allen BL. Dingoes at the Doorstep: Home Range Sizes and Activity Patterns of Dingoes and Other Wild Dogs around Urban Areas of North-Eastern Australia. Animals. 2016; 6(8):48. https://doi.org/10.3390/ani6080048
Chicago/Turabian StyleMcNeill, Alice T., Luke K. -P. Leung, Mark S. Goullet, Matthew N. Gentle, and Benjamin L. Allen. 2016. "Dingoes at the Doorstep: Home Range Sizes and Activity Patterns of Dingoes and Other Wild Dogs around Urban Areas of North-Eastern Australia" Animals 6, no. 8: 48. https://doi.org/10.3390/ani6080048
APA StyleMcNeill, A. T., Leung, L. K.-P., Goullet, M. S., Gentle, M. N., & Allen, B. L. (2016). Dingoes at the Doorstep: Home Range Sizes and Activity Patterns of Dingoes and Other Wild Dogs around Urban Areas of North-Eastern Australia. Animals, 6(8), 48. https://doi.org/10.3390/ani6080048