
Intermittent Suckling Causes a Transient Increase in Cortisol That Does Not Appear to Compromise Selected Measures of Piglet Welfare and Stress †

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, Diets and Housing
2.2. Measurements
2.3. Blood and Plasma Analyses
2.4. Statistical Methods
3. Results
3.1. Piglet Mortality
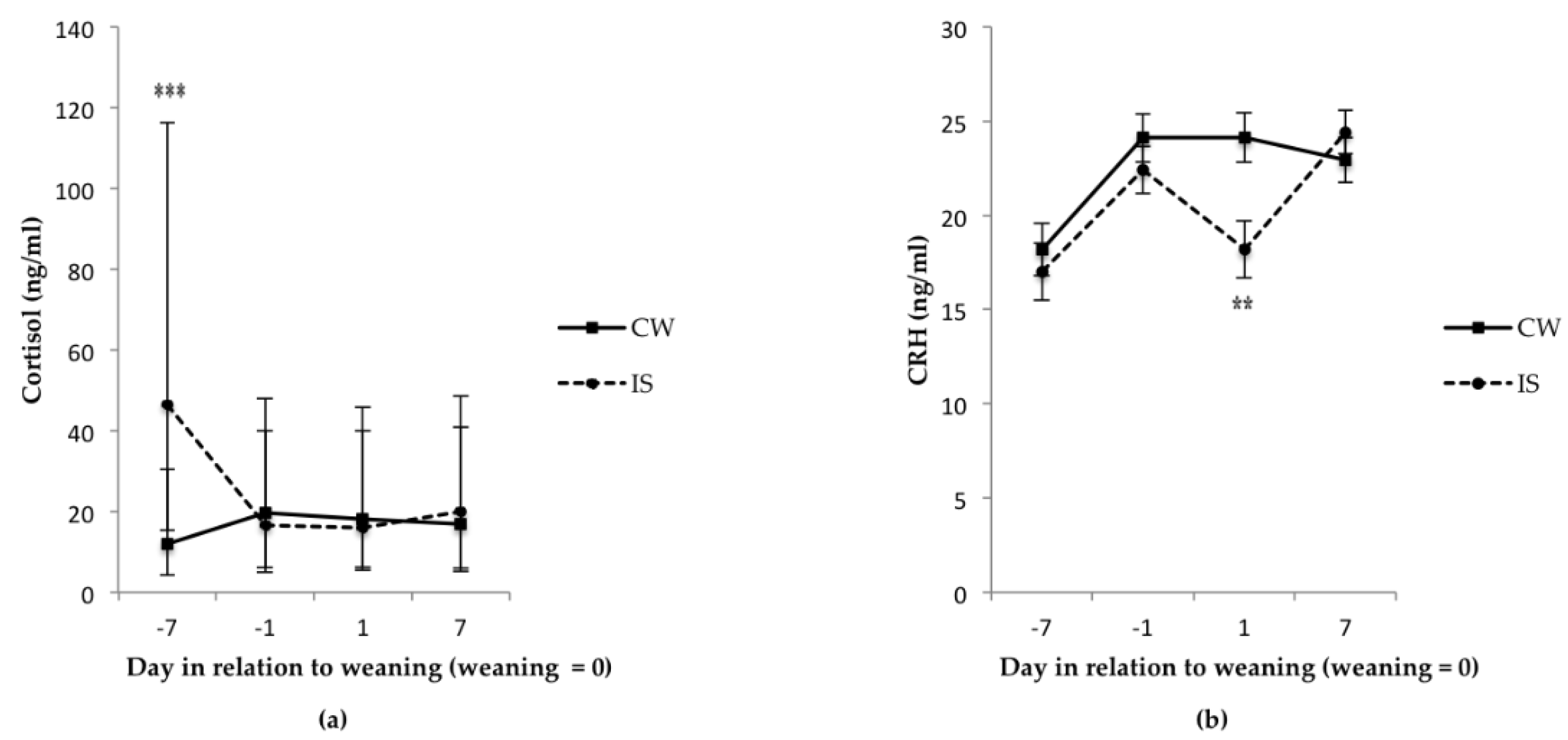
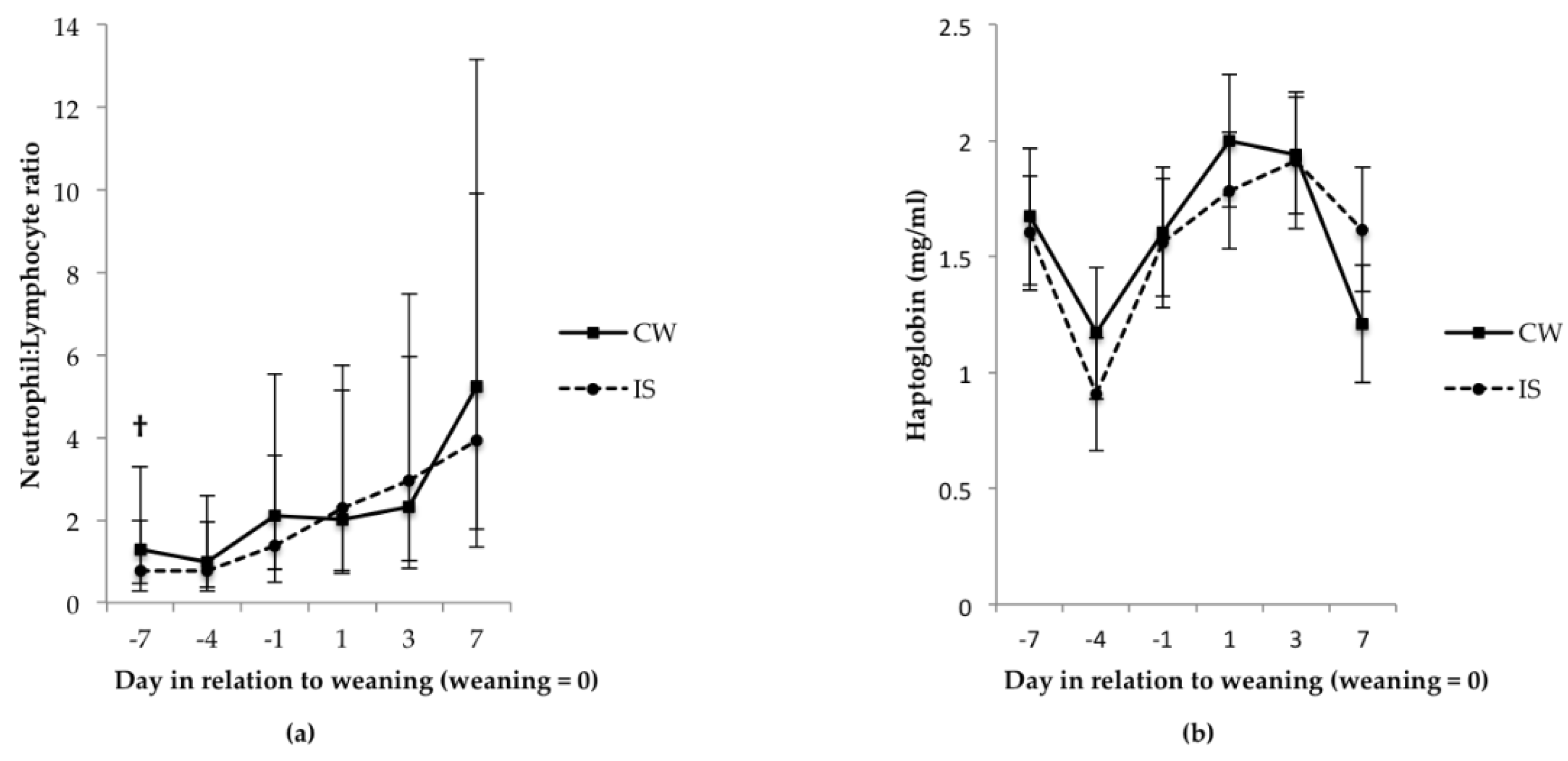
3.2. Blood Measures Indicative of Stress and Welfare, and Injury Scores
3.3. Production Measures
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CRH | corticotrophin releasing hormone |
| CW | corticotrophin releasing hormone |
| IS | Intermittent suckling |
References
- Gerritsen, R.; Soede, N.; Langendijk, P.; Hazeleger, W.; Kemp, B. The intermittent suckling regimen in pigs: Consequences for reproductive performance of sows. Reprod. Domest. Anim. 2008, 43, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Downing, J.; Brock, D.; Smits, R.; Giles, L. Induction of Oestrus in Lactation Results in Normal Mating and Farrowing Performance, Manipulating Pig Production XII, Cairns, Queensland, Australia, 2009; Van Barneveld, R., Ed.; Australasian Pig Science Association (Inc.): Cairns, Australia, 2009; p. 144. [Google Scholar]
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors influencing the structure and function of the small intestine in the weaned pig: A review. Livest. Prod. Sci. 1997, 51, 215–236. [Google Scholar] [CrossRef]
- Bøe, K. The process of weaning in pigs: When the sow decides. Appl. Anim. Behav. Sci. 1991, 30, 47–59. [Google Scholar] [CrossRef]
- Weary, D.M.; Pajor, E.A.; Bonenfant, M.; Fraser, D.; Kramer, D.L. Alternative housing for sows and litters: Part 4. Effects of sow-controlled housing combined with a communal piglet area on pre-and post-weaning behaviour and performance. Appl. Anim. Behav. Sci. 2002, 76, 279–290. [Google Scholar] [CrossRef]
- Kuller, W.; Soede, N.; van Beers-Schreurs, H.; Langendijk, P.; Taverne, M.; Verheijden, J.; Kemp, B. Intermittent suckling: Effects on piglet and sow performance before and after weaning. J. Anim. Sci. 2004, 82, 405–413. [Google Scholar] [PubMed]
- Kuller, W.; Soede, N.; van Beers-Schreurs, H.; Langendijk, P.; Taverne, M.; Kemp, B.; Verheijden, J. Effects of intermittent suckling and creep feed intake on pig performance from birth to slaughter. J. Anim. Sci. 2007, 85, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Berkeveld, M.; Langendijk, P.; van Beers-Schreurs, H.M.; Koets, A.P.; Taverne, M.A.; Verheijden, J.H. Postweaning growth check in pigs is markedly reduced by intermittent suckling and extended lactation. J. Anim. Sci. 2007, 85, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Berkeveld, M.; Langendijk, P.; Soede, N.M.; Kemp, B.; Taverne, M.A.; Verheijden, J.H.; Kuijken, N.; Koets, A.P. Improving adaptation to weaning: Effect of intermittent suckling regimens on piglet feed intake, growth, and gut characteristics. J. Anim. Sci. 2009, 87, 3156–3166. [Google Scholar] [CrossRef] [PubMed]
- Dybkjær, L. The identification of behavioural indicators of “stress” in early weaned piglets. Appl. Anim. Behav. Sci. 1992, 35, 135–147. [Google Scholar] [CrossRef]
- Mormède, P.; Andanson, S.; Aupérin, B.; Beerda, B.; Guémené, D.; Malmkvist, J.; Manteca, X.; Manteuffel, G.; Prunet, P.; van Reenen, C.G.; et al. Exploration of the hypothalamic-pituitary-adrenal function as a tool to evaluate animal welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef] [PubMed]
- Widowski, T.; Cottrell, T.; Dewey, C.; Friendship, R. Observations of piglet-directed behavior patterns and skin lesions in eleven commercial swine herds. J. Swine Health Prod. 2003, 11, 181–185. [Google Scholar]
- Stevenson, J.S.; Davis, D.L. Influence of reduced litter size and daily litter separation on fertility of sows at 2 to 5 weeks postpartum. J. Anim. Sci. 1984, 59, 284–293. [Google Scholar] [PubMed]
- Newton, E.A.; Stevenson, J.S.; Davis, D.L. Influence of duration of litter separation and boar exposure on estrous expression of sows during and after lactation. J. Anim. Sci. 1987, 65, 1500–1506. [Google Scholar] [PubMed]
- Moberg, G. Biological response to stress: Implications for animal welfare. In The Biology of Animal Stress: Basic Principles and Implications for Animal Welfare; CABI Publishing: Wallingford, UK, 2000; pp. 1–21. [Google Scholar]
- Hemsworth, P.; Mellor, D.; Cronin, G.; Tilbrook, A. Scientific assessment of animal welfare. N. Z. Vet. J. 2015, 63, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Klemcke, H.G.; Pond, W.G. Porcine adrenal adrenocorticotropic hormone receptors: Characterization, changes during neonatal development, and response to a stressor. Endocrinology 1991, 128, 2476–2488. [Google Scholar] [CrossRef] [PubMed]
- Kanitz, E.; Tuchscherer, M.; Puppe, B.; Tuchscherer, A.; Stabenow, B. Consequences of repeated early isolation in domestic piglets (Sus scrofa) on their behavioural, neuroendocrine, and immunological responses. Brain Behav. Immun. 2004, 18, 35–45. [Google Scholar] [CrossRef]
- McCormick, C.M.; Kehoe, P.; Kovacs, S. Corticosterone release in response to repeated, short episodes of neonatal isolation: Evidence of sensitization. Int. J. Dev. Neurosci. 1998, 16, 175–185. [Google Scholar] [CrossRef]
- Berkeveld, M.; Langendijk, P.; Bolhuis, J.E.; Koets, A.P.; Verheijden, J.H.; Taverne, M.A. Intermittent suckling during an extended lactation period: Effects on piglet behavior. J. Anim. Sci. 2007, 85, 3415–3424. [Google Scholar] [CrossRef] [PubMed]
- Van der Meulen, J.; Koopmans, S.; Dekker, R.; Hoogendoorn, A. Increasing weaning age of piglets from 4 to 7 weeks reduces stress, increases post-weaning feed intake but does not improve intestinal functionality. Animal 2010, 4, 1653–1661. [Google Scholar] [CrossRef] [PubMed]
- Hay, M.; Orgeur, P.; Lévy, F.; Le Dividich, J.; Concordet, D.; Nowak, R.; Schaal, B.S.; Mormède, P. Neuroendocrine consequences of very early weaning in swine. Physiol. Behav. 2001, 72, 263–269. [Google Scholar] [CrossRef]
- Mason, S.P.; Jarvis, S.; Lawrence, A.B. Individual differences in responses of piglets to weaning at different ages. Appl. Anim. Behav. Sci. 2003, 80, 117–132. [Google Scholar] [CrossRef]
- Moeser, A.J.; Vander Klok, C.; Ryan, K.A.; Wooten, J.G.; Little, D.; Cook, V.L.; Blikslager, A.T. Stress signaling pathways activated by weaning mediate intestinal dysfunction in the pig. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G173–G181. [Google Scholar] [CrossRef] [PubMed]
- Worsaae, H.; Schmidt, M. Plasma cortisol and behaviour in early weaned piglets. Acta Vet. Scand. 1980, 21, 640–657. [Google Scholar] [PubMed]
- Worobec, E.K.; Duncan, I.J.H.; Widowski, T.M. The effects of weaning at 7, 14 and 28 days on piglet behaviour. Appl. Anim. Behav. Sci. 1999, 62, 173–182. [Google Scholar] [CrossRef]
- Sales, N.; Collins, D.; Collins, A.; McKenna, T.; Bauer, M.; Parke, C.; Hermesch, S. Porcine haptoglobin levels measured at 7–14 days after weaning were independent of age, weight or gender. Anim. Prod. Sci. 2015, 55, 1457–1457. [Google Scholar]
- Piñeiro, C.; Piñeiro, M.; Morales, J.; Carpintero, R.; Campbell, F.; Eckersall, P.; Toussaint, M.; Alava, M.; Lampreave, F. Pig acute-phase protein levels after stress induced by changes in the pattern of food administration. Animal 2007, 1, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Eckersall, P.; Bell, R. Acute phase proteins: Biomarkers of infection and inflammation in veterinary medicine. Vet. J. 2010, 185, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Hicks, T.A.; McGlone, J.J.; Whisnant, C.S.; Kattesh, H.G.; Norman, R.L. Behavioral, endocrine, immune, and performance measures for pigs exposed to acute stress. J. Anim. Sci. 1998, 76, 474–483. [Google Scholar] [PubMed]
- Sorensen, N.; Tegtmeier, C.; Andresen, L.O.; Pineiro, M.; Toussaint, M.; Campbell, F.; Lampreave, F.; Heegaard, P.M. The porcine acute phase protein response to acute clinical and subclinical experimental infection with streptococcus suis. Vet. Immunol. Immunopathol. 2006, 113, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Heegaard, P.M.; Klausen, J.; Nielsen, J.P.; González-Ramón, N.; Piñeiro, M.; Lampreave, F.; Alava, M.A. The porcine acute phase response to infection with Actinobacillus pleuropneumoniae. Haptoglobin, c-reactive protein, major acute phase protein and serum amyloid a protein are sensitive indicators of infection. Compar. Biochem. Physiol. Part B: Biochem. Mol. Biol. 1998, 119, 365–373. [Google Scholar] [CrossRef]
- Jungersen, G.; Jensen, L.; Riber, U.; Heegaard, P.M.; Petersen, E.; Poulsen, J.; Bille-Hansen, V.; Lind, P. Pathogenicity of selected Toxoplasma gondii isolates in young pigs. Int. J. Parasitol. 1999, 29, 1307–1319. [Google Scholar] [CrossRef]
- Sauerwein, H.; Schmitz, S.; Hiss, S. The acute phase protein haptoglobin and its relation to oxidative status in piglets undergoing weaning-induced stress. Redox Rep. 2005, 10, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Vente-Spreeuwenberg, M.; Verdonk, J.; Verstegen, M.; Beynen, A. Villus height and gut development in weaned piglets receiving diets containing either glucose, lactose or starch. Br. J. Nutr. 2003, 90, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Piñeiro, C.; Piñeiro, M.; Morales, J.; Andrés, M.; Lorenzo, E.; del Pozo, M.; Alava, M.A.; Lampreave, F. Pig-map and haptoglobin concentration reference values in swine from commercial farms. Vet. J. 2009, 179, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Petersen, H.; Nielsen, J.; Jensen, A.; Heegaard, P.M. Evaluation of an enzyme-linked immunosorbent assay for determination of porcine haptoglobin. J. Vet. Med. 2001, 48, 513–523. [Google Scholar] [CrossRef]
- Blecha, F.; Pollman, D.; Nichols, D. Weaning pigs at an early age decreases cellular immunity. J. Anim. Sci. 1983, 56, 396–400. [Google Scholar] [PubMed]
- Blecha, F.; Pollmann, D.; Nichols, D. Immunologic reactions of pigs regrouped at or near weaning. Am. J. Vet. Res. 1985, 46, 1934–1937. [Google Scholar] [PubMed]
- Pié, S.; Lalles, J.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [PubMed]
- Moeser, A.J.; Ryan, K.A.; Nighot, P.K.; Blikslager, A.T. Gastrointestinal dysfunction induced by early weaning is attenuated by delayed weaning and mast cell blockade in pigs. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G413–G421. [Google Scholar] [CrossRef] [PubMed]
- Puppe, B.; Tuchscherer, M.; Tuchscherer, A. The effect of housing conditions and social environment immediately after weaning on the agonistic behaviour, neutrophil/lymphocyte ratio, and plasma glucose level in pigs. Livest. Prod. Sci. 1997, 48, 157–164. [Google Scholar] [CrossRef]
- Metz, J.H.M.; Gonyou, H.W. Effect of age and housing conditions on the behavioural and haemolytic reaction of piglets to weaning. Appl. Anim. Behav. Sci. 1990, 27, 299–309. [Google Scholar] [CrossRef]
- Kiliaan, A.J.; Saunders, P.R.; Bijlsma, P.B.; Berin, M.C.; Taminiau, J.A.; Groot, J.A.; Perdue, M.H. Stress stimulates transepithelial macromolecular uptake in rat jejunum. Am. J. Physiol. Gastrointest. Liver Physiol. 1998, 275, G1037–G1044. [Google Scholar]
- Meddings, J.B.; Swain, M. Environmental stress-induced gastrointestinal permeability is mediated by endogenous glucocorticoids in the rat. Gastroenterology 2000, 119, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.; Blisklager, A.T.; Moeser, A.J. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G352–G363. [Google Scholar] [CrossRef] [PubMed]
- Nabuurs, M.; Hoogendoorn, A.; Van Zijderveld-Van Bemmel, A. Effect of supplementary feeding during the sucking period on net absorption from the small intestine of weaned pigs. Res. Vet. Sci. 1996, 61, 72–77. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Day 1 | Weaning Regimen 2 | SEM | p-Value | |
|---|---|---|---|---|
| CW | IS | |||
| −14 | 4.8 | 4.5 | 0.26 | 0.42 |
| −7 | 6.4 | 6.0 | 0.34 | 0.44 |
| −4 | 7.0 | 6.5 | 0.38 | 0.33 |
| 0 | 8.2 | 7.4 | 0.45 | 0.22 |
| 3 | 8.2 | 7.3 | 0.23 | 0.23 |
| 7 | 8.7 | 8.1 | 0.26 | 0.93 |
| Item | Weaning Regimen 1 | SEM | p-Value | |
|---|---|---|---|---|
| CW | IS | |||
| ADG (g) | ||||
| Before weaning | ||||
| Day −14 2 to day −7 | 223 | 222 | 14.8 | 0.63 |
| Day −7 to day −4 | 210 | 184 | 31.4 | 0.38 |
| Day −4 to weaning | 281 | 212 | 38.9 | 0.20 |
| After weaning | ||||
| Weaning to day 3 3 | 2 | −26 | 43.1 | 0.63 |
| Day 3 to day 7 3 | 134 | 209 | 28.7 | 0.07 |
| ADFI (g) 4 | ||||
| Day −14 to day −7 | 6 (3.81–10.59) | 8 (4.85–12.22) | 0.57 | |
| Day −7 to day −4 | 14 (7.24–26.42) | 42 (22.54–77.09) | 0.02 | |
| Day −4 to weaning | 25 (12.30–50.12) | 57 (30.27–106.41) | 0.08 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turpin, D.L.; Langendijk, P.; Chen, T.-Y.; Lines, D.; Pluske, J.R. Intermittent Suckling Causes a Transient Increase in Cortisol That Does Not Appear to Compromise Selected Measures of Piglet Welfare and Stress. Animals 2016, 6, 24. https://doi.org/10.3390/ani6030024
Turpin DL, Langendijk P, Chen T-Y, Lines D, Pluske JR. Intermittent Suckling Causes a Transient Increase in Cortisol That Does Not Appear to Compromise Selected Measures of Piglet Welfare and Stress. Animals. 2016; 6(3):24. https://doi.org/10.3390/ani6030024
Chicago/Turabian StyleTurpin, Diana L., Pieter Langendijk, Tai-Yuan Chen, David Lines, and John R. Pluske. 2016. "Intermittent Suckling Causes a Transient Increase in Cortisol That Does Not Appear to Compromise Selected Measures of Piglet Welfare and Stress" Animals 6, no. 3: 24. https://doi.org/10.3390/ani6030024
APA StyleTurpin, D. L., Langendijk, P., Chen, T.-Y., Lines, D., & Pluske, J. R. (2016). Intermittent Suckling Causes a Transient Increase in Cortisol That Does Not Appear to Compromise Selected Measures of Piglet Welfare and Stress. Animals, 6(3), 24. https://doi.org/10.3390/ani6030024