
Selecting a Conservation Surrogate Species for Small Fragmented Habitats Using Ecological Niche Modelling

Simple Summary
Abstract
1. Introduction
2. Experimental Section
2.1. The Study Area and Its Inhabitants
2.2. Shortlist Criteria and Locally Appropriate Flagship Characteristics
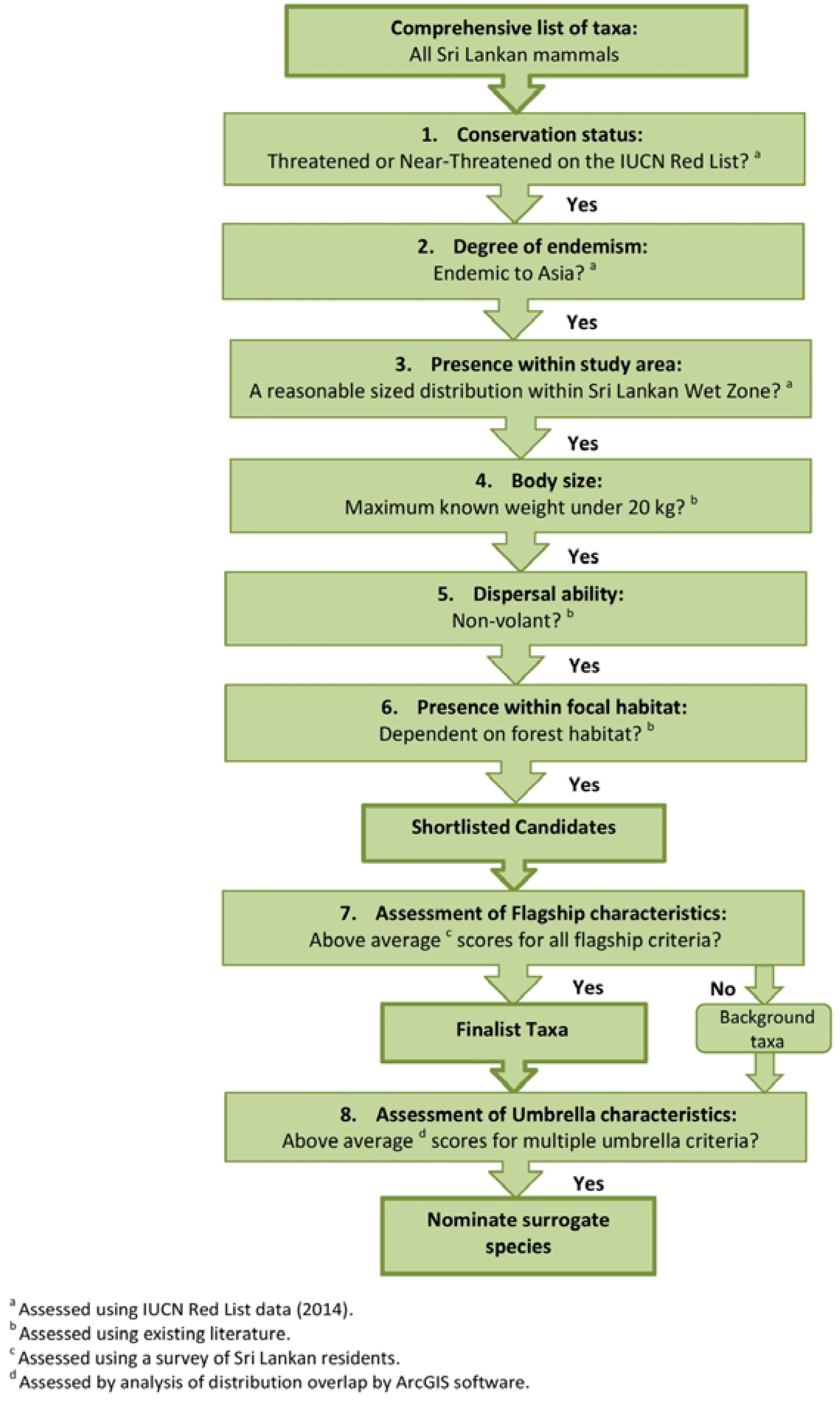
2.3. Ecological Niche Modelling (ENM)
2.4. Assessment of Umbrella Characteristics
- (1)
- Overlap in geographic range: For each species acting as umbrella, we calculated the percentage of the beneficiary species’ range in the study area that occurred within the range of the umbrella [42].
- (2)
- Number of species protected: If more than 75% of the beneficiary species’ range within the study area was afforded cover, we counted it as “protected”.
- (3)
- Umbrella efficiency was defined as the level of overlap per km2 of the umbrella’s range. We calculated this for each umbrella by dividing the mean area of overlap from all beneficiary species, by the umbrella’s total range within the study area.
3. Results
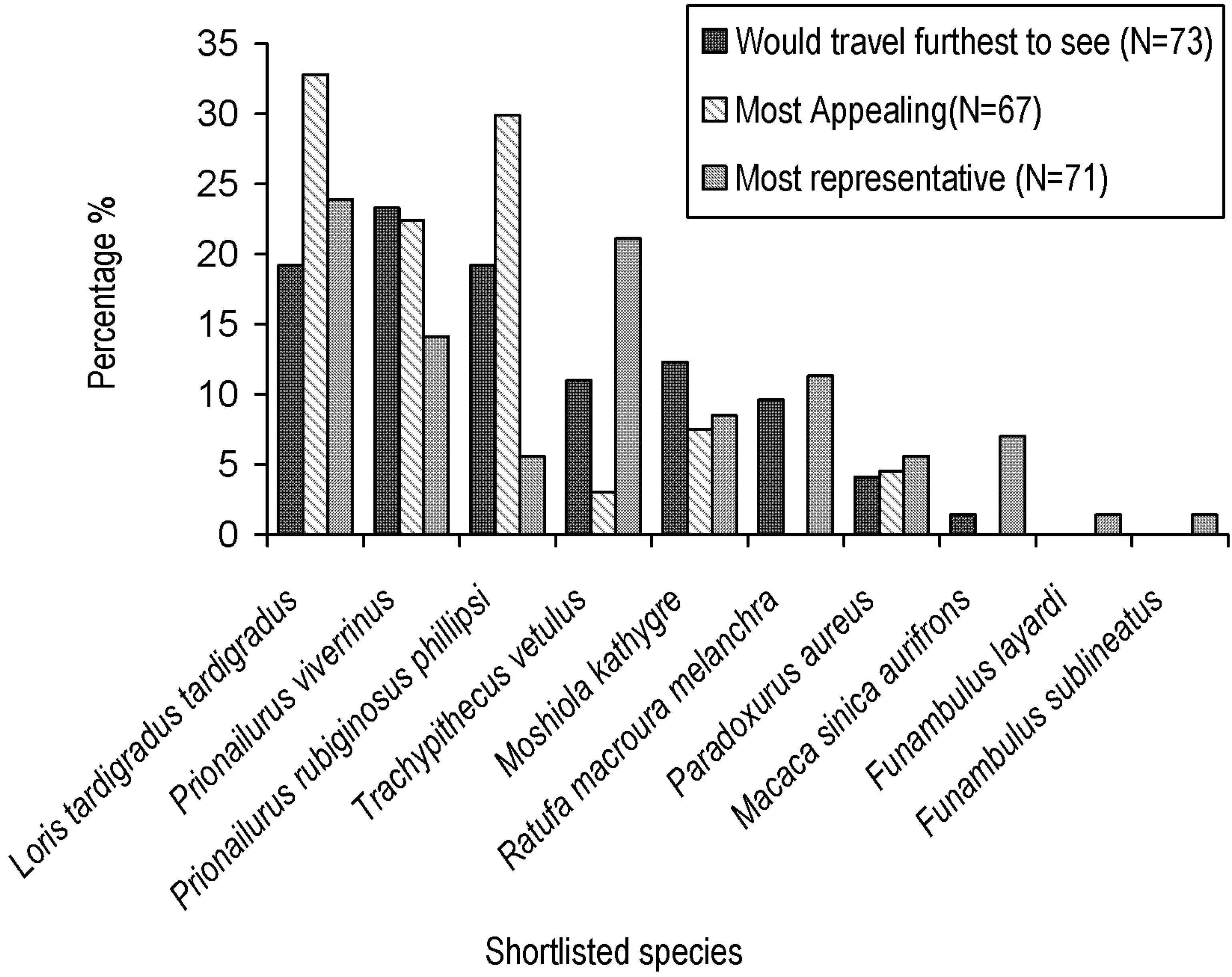
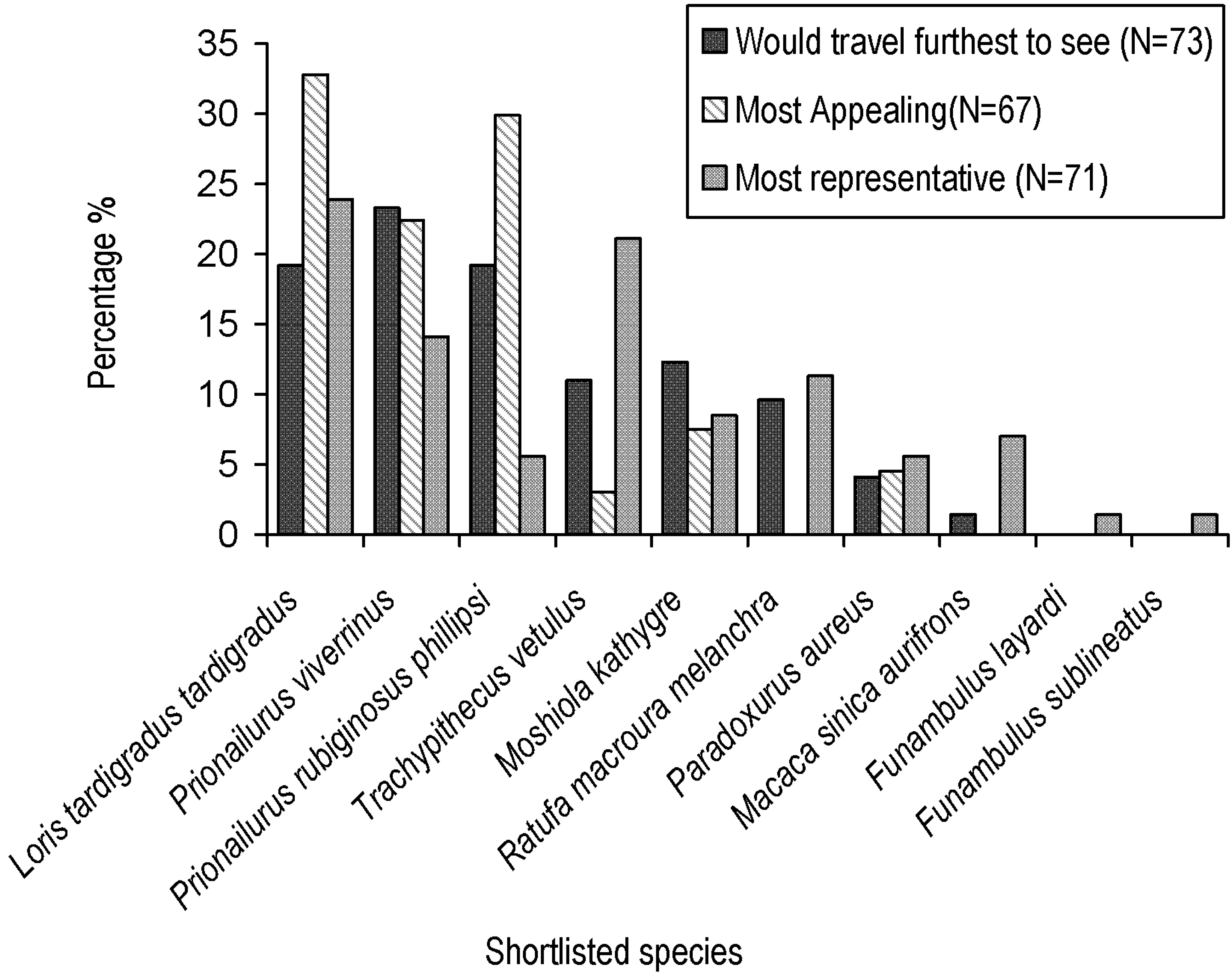
3.1. Shortlist Criteria and Locally Appropriate Flagship Characteristics
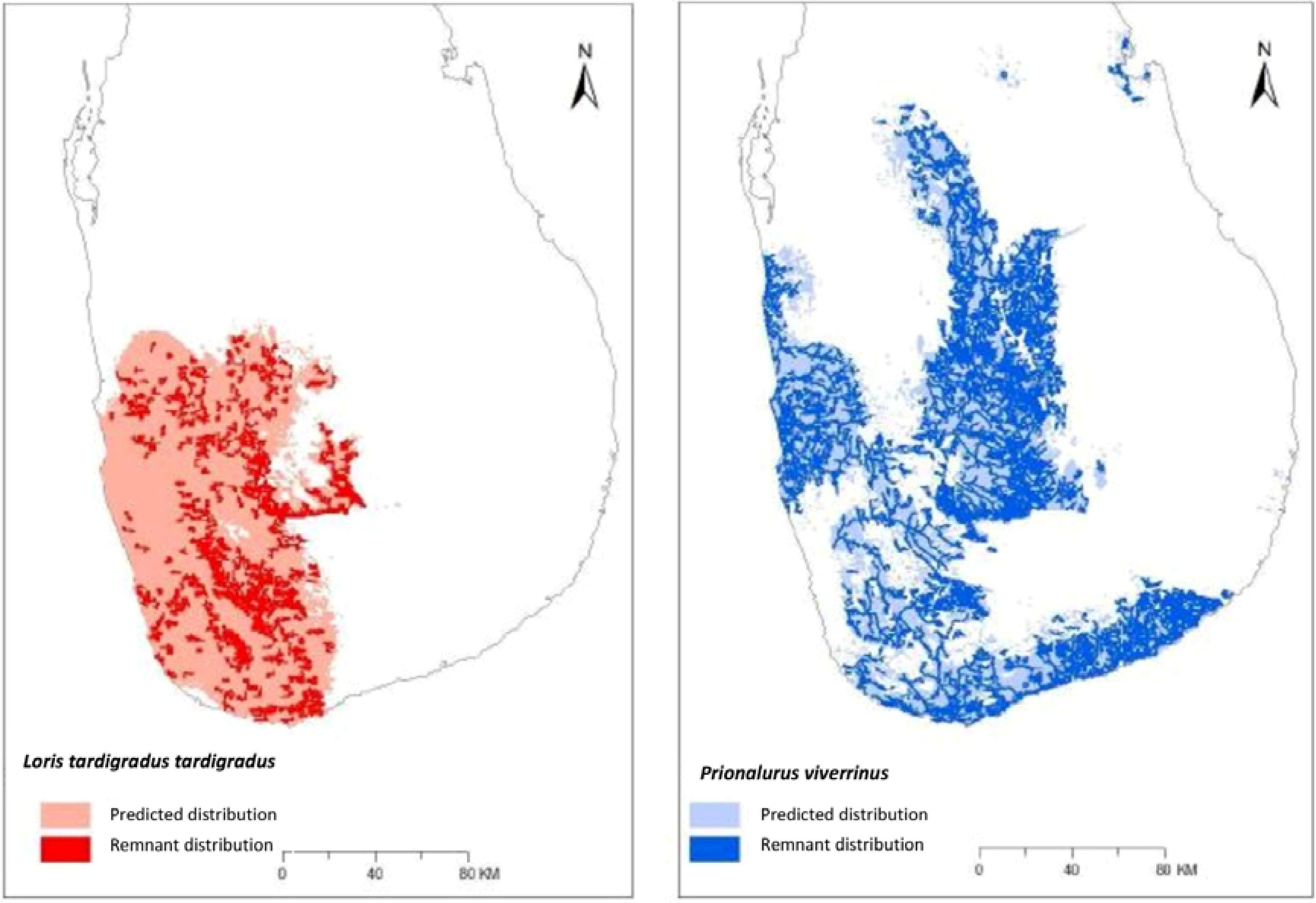
3.2. Ecological Niche Modelling
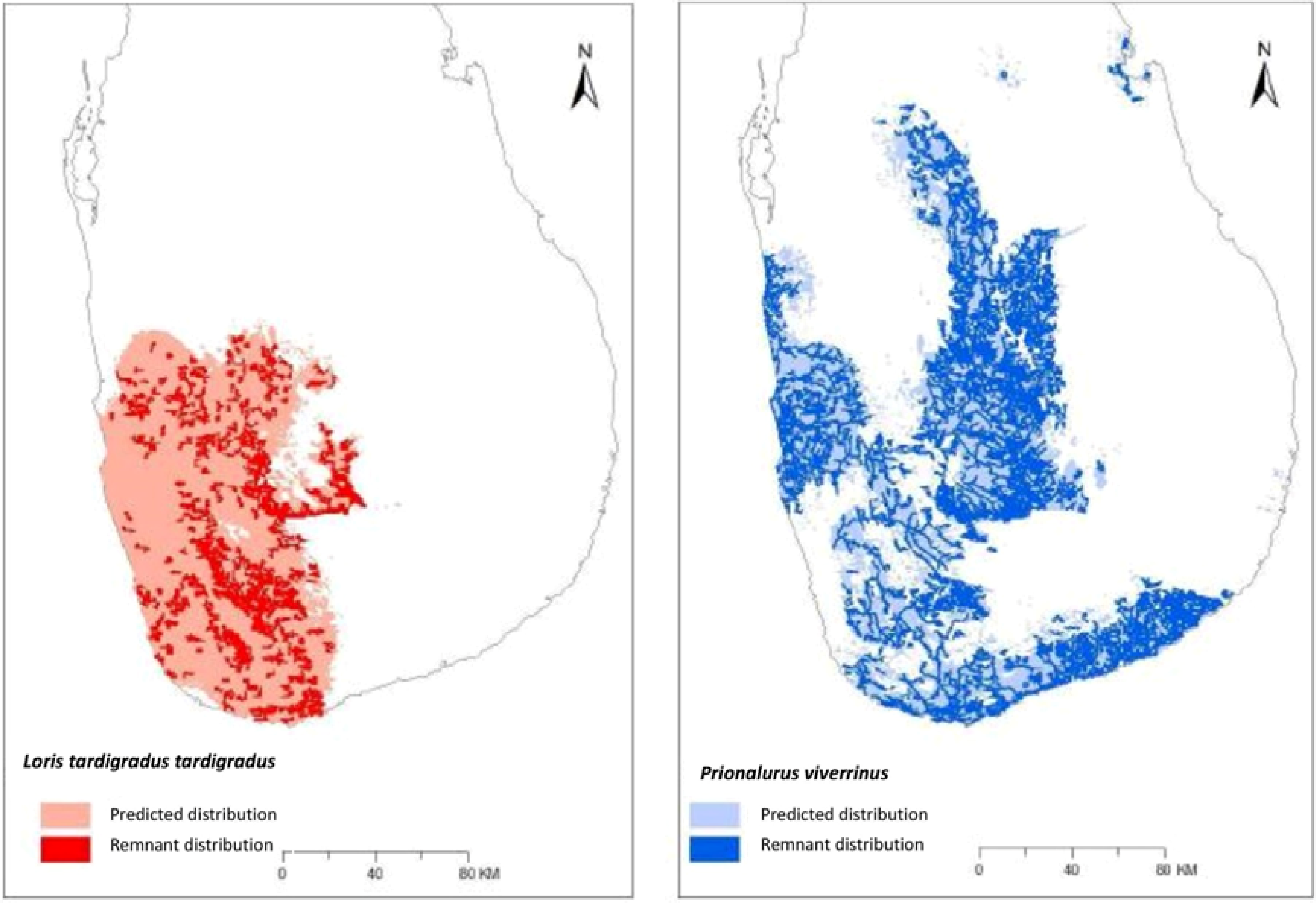
3.3. Overlap within Geographic Range
3.4. Number of Species Protected

{kind=link}
{kind=link}
{kind=link}
| Candidates | Protection Afforded to Beneficiary Species | ||
|---|---|---|---|
| No. of Species Afforded >50% Cover | No. of Species Afforded >75% Cover | No. of Species Afforded >95% Cover | |
| Prionailurus rubiginosus phillipsi | 8 | 7 | 2 |
| Moschiola kathygre | 9 | 9 | 3 |
| Ratufa macroura melanochra | 4 | 1 | 1 |
| Prionalurus viverrinus | 6 | 0 | 0 |
| Paradoxurus aureus | 7 | 3 | 0 |
| Funambulus layardi | 3 | 0 | 0 |
| Funambulus sublineatus | 6 | 2 | 0 |
| Trachypithecus vetulus vetulus | 2 | 0 | 0 |
| Macaca sinica aurifrons | 7 | 4 | 0 |
| Loris tardigradus tardigradus | 7 | 5 | 1 |
3.5. Overlap Efficiency
3.6. Selecting Cinderella Species
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Caro, T.M.; O’Doherty, G. On the use of surrogate species in conservation biology. Conserv. Biol. 1999, 13, 805–814. [Google Scholar] [CrossRef]
- Seddon, P.J.; Leech, T. Conservation shortcut, or long winding road? A critique of umbrella species criteria. Oryx 2008, 42, 240–245. [Google Scholar] [CrossRef]
- Simberloff, D. Flagships, umbrellas, and keystones: Is single-species management passe in the landscape era? Biol. Conserv. 1998, 83, 247–257. [Google Scholar] [CrossRef]
- Leader-Williams, N.; Dublin, H.T. Charismatic megafauna as flagship species. In A Review of Priorities for the Conservation of Mammalian Diversity: Has the Panda Had Its Day? Entwistle, A., Dunstone, N., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 53–81. [Google Scholar]
- Dietz, J.M.; Dietz, L.A.; Nagagata, E.Y. The effective use of flagship species for conservation of biodiversity: The example of lion tamarins in Brazil. In Conservation: Interactive Management of Wild and Captive Populations; Mace, G.M., Feistner, A.T.C., Olney, P.J., Eds.; Chapman & Hall: London, UK, 1994; pp. 32–49. [Google Scholar]
- Bandara, R.; Tisdell, C. Changing abundance of elephants and willingness to pay for their conservation. J. Environ. Manag. 2005, 76, 47–59. [Google Scholar] [CrossRef]
- Smith, R.J.; Veríssimo, D.; Isaac, N.J.B.; Jones, K.E. Identifying Cinderella species: Uncovering mammals with conservation flagship appeal. Conserv. Lett. 2012, 5, 205–212. [Google Scholar] [CrossRef]
- Clucas, B.; McHugh, K.; Caro, T. Flagship species on covers of US conservation and nature magazines. Biodivers. Conserv. 2008, 17, 1517–1528. [Google Scholar] [CrossRef]
- Mulder, M.B.; Schacht, R.; Caro, T.; Schacht, J.; Caro, B. Knowledge and attitudes of children of the Rupununi: Implications for conservation in Guyana. Biol. Conserv. 2009, 142, 879–887. [Google Scholar] [CrossRef]
- Andelman, S.J.; Fagan, W.F. Umbrellas and flagships: Efficient conservation surrogates or expensive mistakes? Proc. Nat. Acad. Sci. USA 2000, 97, 5954–5959. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.H.; Burgess, N.D.; Rahbek, C. Flagship species, ecological complementarity and conserving the diversity of mammals and birds in sub-Saharan Africa. Anim. Conserv. 2000, 3, 249–260. [Google Scholar] [CrossRef]
- Caro, T.; Engilis, A., Jr.; Fitzherbert, E.; Gardner, T. Preliminary assessment of the flagship species concept at a small scale. Anim. Conserv. 2004, 7, 63–70. [Google Scholar] [CrossRef]
- Laurance, W.F. Ecological correlates of extinction proneness in Australian tropical rain-forest mammals. Conserv. Biol. 1991, 5, 79–89. [Google Scholar] [CrossRef]
- Davies, K.F.; Margules, C.R.; Lawrence, J.F. Which traits of species predict population declines in experimental forest fragments? Ecology 2000, 81, 1450–1461. [Google Scholar] [CrossRef]
- Canale, G.R.; Peres, C.A.; Guidorizzi, C.E.; Gatto, C.A.F.; Kierulff, M.C.M. Pervasive defaunation of forest remnants in a tropical biodiversity hotspot. PLOS ONE 2012, 7, e41671. [Google Scholar] [CrossRef] [PubMed]
- Turner, I.M. Species loss in fragments of tropical rain forest: A review of the evidence. J. Appl. Ecol. 1996, 33, 200–209. [Google Scholar] [CrossRef]
- Turner, I.M.; Corlett, R.T. The conservation value of small, isolated fragments of lowland tropical rain forest. TREE 1996, 11, 330–333. [Google Scholar] [PubMed]
- Melbourne, B.A.; Davies, K.F.; Margules, C.R.; Lindenmayer, D.B.; Saunders, D.A.; Wissel, C.; Henle, K. Species survival in fragmented landscapes: Where to from here? Biodivers. Conserv. 2004, 13, 275–284. [Google Scholar] [CrossRef]
- Brancalion, P.H.S.; Melo, F.P.L.; Tabarelli, M.; Rodrigues, R.R. Biodiversity persistence in highly human-modified tropical landscapes depends on ecological restoration. Conserv. Lett. 2013, 6, 705–710. [Google Scholar]
- Marsh, L.K. Because conservation counts: Primates and fragmentation. In Primates in Fragments: Complexity and Resilience; Marsh, L.K., Chapman, C., Eds.; Springer: New York, NY, USA, 2013; pp. 3–12. [Google Scholar]
- Entwistle, A. Flagships for the future? Oryx 2000, 34, 239–240. [Google Scholar]
- Dasken, C.D. The human impact on rain forests in Sri Lanka: The case of “Sinharaja” rainforest reserve. Acta Geogr. (Trondheim) 2005, 10, 49–63. [Google Scholar]
- Gunawardene, N.R.; Daniels, A.E.D.; Gunatilleke, I.; Gunatilleke, C.V.S.; Karunakaran, P.V.; Nayak, K.G.; Prasad, S.; Puyravaud, P.; Ramesh, B.R.; Subramanian, K.A.; et al. A brief overview of the Western Ghats Sri Lanka biodiversity hotspot. Curr. Sci. 2007, 93, 1567–1572. [Google Scholar]
- Daily News—Top seven wild Sri Lanka as Flagship Species. Available online: http://www.dailynews.lk/?q=local/top-seven-wild-sri-lanka-flagship-species/ (accessed on 11 November 2014).
- The IUCN Red List of Threatened Species. Version 2014.2. Available online: http://www.iucnredlist.org/ (accessed on 7 November 2014).
- Nahallage, C.A.D.; Huffman, M.A.; Kuruppu, N.; Weerasingha, T. Diurnal primates in Sri Lanka and people’s perception of them. Prim. Conserv. 2008, 23, 81–87. [Google Scholar] [CrossRef]
- Bhandari, B.B.; Abe, O. Environmental education in the Asia-Pacific region: Some problems and prospects. Int. Rev. Environ. Strateg. 2000, 1, 57–77. [Google Scholar]
- Erdelen, W. Forest ecosystems and nature conservation in Sri Lanka. Biol. Conserv. 1988, 43, 115–135. [Google Scholar] [CrossRef]
- Rudran, R. A survey of Sri Lanka’s Endangered and endemic Western purple-faced langur (Trachypithecus vetulus nestor). Prim. Conserv. 2007, 22, 139–144. [Google Scholar] [CrossRef]
- Department of Census and Statistics—Sri Lanka. Available online: http://www.statistics.gov.lk/ (accessed on 14 January 2014).
- Couper, M.; Kapteyn, A.; Schonlau, M.; Winter, J. Noncoverage and nonresponse in an Internet survey. Soc. Sci. Res. 2007, 36, 131–148. [Google Scholar] [CrossRef]
- Phillips, S.J.; Pearson, R.G.; Schapire, R.E. Maximum entropy modelling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudik, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distr. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Anderson, R.P.; Martinez-Meyer, E. Modelling species’ geographic distributions for preliminary conservation assessments: An implementation with the spiny pocket mice (Heteromys) of Ecuador. Biol. Conserv. 2004, 116, 167–179. [Google Scholar] [CrossRef]
- Thorn, J.S.; Nijman, V.; Smith, D.; Nekaris, K.A.I. Ecological niche modelling as a technique for assessing threats setting conservation priorities for Asian slow lorises (Primates: Nycticebus). Divers. Distrib. 2009, 15, 289–298. [Google Scholar] [CrossRef]
- International Steering Committee for Global Mapping. Available online: http://www.iscgm.org/ (accessed on 7 June 2012).
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Peterson, T.; Papeşa, M.; Soberóna, J. Rethinking receiver operating characteristic analysis applications in ecological niche modelling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Worldclim: Global Climate Data. Available online: http://www.worldclim.org/ (accessed on 11 June 2012).
- Gunatilleke, I.A.U.N.; Gunatilleke, C.V.S. Distribution of foristic richness and its conservation in Sri Lanka. Conserv. Biol. 1990, 4, 21–31. [Google Scholar] [CrossRef]
- Nekaris, K.A.I.; Jayewardene, J. Survey of the slender loris (Primates, Lorisidae Gray, 1821: Loris tardigradus Linnaeus, 1758 and Loris lydekkerianus Cabrera, 1908) in Sri Lanka. J. Zool. 2004, 262, 1–12. [Google Scholar] [CrossRef]
- Rowland, M.M.; Wisdom, M.J.; Meinke, C.W.; Suring, L.H. Utility of greater sage-grouse as an umbrella species. In Habitat Threats in the Sagebrush Ecosystem—Methods of Regional Assessment Applications in the Great Basin; Wisdom, M.J., Rowland, M.M., Suring, L.H., Eds.; Allen Press: Lawrence, KS, USA, 2005; pp. 232–249. [Google Scholar]
- Fleishman, E.; Murphy, D.D.; Brussard, P.F. A new method for selection of umbrella species for conservation planning. Ecol. Appl. 2000, 10, 569–579. [Google Scholar] [CrossRef]
- Roberge, J.M.; Angelstam, P. Usefulness of the umbrella species concept as a conservation tool. Conserv. Biol. 2004, 18, 76–85. [Google Scholar] [CrossRef]
- Morris, D.W.; Kotler, B.P.; Brown, J.S.; Sundararaj, V.; Ale, S.B. Behavioral Indicators for Conserving Mammal Diversity. Year Ecol. Conserv. Biol. 2009, 1162, 334–356. [Google Scholar]
- Nekaris, K.A.I.; Faculty of Humanities and Social Sciences, Oxford Brookes University, Oxford, UK. Personal Observation, 2014.
- Nijman, V.; Nekaris, K.A.I. Testing a model for predicting primate crop-raiding using crop-and farm-specific risk values. Appl. Anim. Behav. Sci. 2010, 127, 125–129. [Google Scholar] [CrossRef]
- Kinan, I.; Dalzell, P. Sea turtles as a flagship species: Different perspectives create conflicts in the Pacific Islands. Mast 2005, 3/4, 195–212. [Google Scholar]
- Nekaris, K.A.I.; Shepherd, C.R.; Starr, C.R.; Nijman, V. Exploring cultural drivers for wildlife trade via an ethnoprimatological approach: A case study of slender and slow lorises (Loris and Nycticebus) in South and Southeast Asia. Am. J. Prim. 2010, 72, 877–886. [Google Scholar] [CrossRef]
- Nekaris, K.A.I.; Boulton, A.; Nijman, V. An ethnoprimatological approach to assessing levels of tolerance between human and commensal non-human primates in Sri Lanka. J. Anthropol. Sci. 2013, 91, 1–14. [Google Scholar]
- Somander, S.V.O. Ceylon’s slender loris: Queer habits of a small nocturnal forest monkey. Loris 1969, 11, 298–300. [Google Scholar]
- Entwistle, A.; Stephenson, P. Small mammals and the conservation agenda. In A Review of Priorities for the Conservation of Mammalian Diversity: Has the Panda Had Its Day? Entwistle, A., Dunstone, N., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 120–142. [Google Scholar]
- Nijman, V.; Nekaris, K.A.I. Effects of deforestation on attitudes and levels of tolerance towards commensal primates (Cercopithecidae) in Sri Lanka. Int. J. Pest Manag. 2010, 56, 153–158. [Google Scholar] [CrossRef]
- Bandara, R.; Tisdell, C. Comparison of rural and urban attitudes to the conservation of Asian elephants in Sri Lanka: Empirical evidence. Biol. Conserv. 2003, 110, 327–342. [Google Scholar] [CrossRef]
- Knight, A.T.; Cowling, R.M. Embracing opportunism in the selection of priority conservation areas. Conserv. Biol. 2007, 21, 1124–1126. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, M.; Arponen, A.; van Teeffelen, A. Top predators: Hot or not? A call for systematic assessment of biodiversity surrogates. J. Appl. Ecol. 2008, 45, 976–980. [Google Scholar] [CrossRef]
- Suter, W.; Graf, R.F.; Hess, R. Capercaillie (Tetrao urogallus) avian biodiversity: Testing the umbrella-species concept. Conserv. Biol. 2002, 16, 778–788. [Google Scholar] [CrossRef]
- Ozaki, K.; Isono, M.; Kawahara, T.; Iida, S.; Kudo, T.; Fukuyama, K. A mechanistic approach to evaluation of umbrella species as conservation surrogates. Conserv. Biol. 2006, 20, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Frazier, J.G. Marine turtles: The role of flagship species in interactions between people and the sea. Marit. Stud. 2005, 3, 5–38. [Google Scholar]
- Bowen-Jones, E.; Entwistle, A. Identifying appropriate flagship species: The importance of culture and local contexts. Oryx 2002, 36, 189–195. [Google Scholar] [CrossRef]
- Molina, R.; Marcot, B.G. Definitions and attributes of little-known species. In Conservation of Rare or Little Known Species; Raphael, M.G., Molina, R., Eds.; Island Press: London, UK; pp. 67–92.
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nekaris, K.A.-I.; Arnell, A.P.; Svensson, M.S. Selecting a Conservation Surrogate Species for Small Fragmented Habitats Using Ecological Niche Modelling. Animals 2015, 5, 27-40. https://doi.org/10.3390/ani5010027
Nekaris KA-I, Arnell AP, Svensson MS. Selecting a Conservation Surrogate Species for Small Fragmented Habitats Using Ecological Niche Modelling. Animals. 2015; 5(1):27-40. https://doi.org/10.3390/ani5010027
Chicago/Turabian StyleNekaris, K. Anne-Isola, Andrew P. Arnell, and Magdalena S. Svensson. 2015. "Selecting a Conservation Surrogate Species for Small Fragmented Habitats Using Ecological Niche Modelling" Animals 5, no. 1: 27-40. https://doi.org/10.3390/ani5010027
APA StyleNekaris, K. A.-I., Arnell, A. P., & Svensson, M. S. (2015). Selecting a Conservation Surrogate Species for Small Fragmented Habitats Using Ecological Niche Modelling. Animals, 5(1), 27-40. https://doi.org/10.3390/ani5010027