Exploration of the Hypothalamic-Pituitary-Adrenal Axis to Improve Animal Welfare by Means of Genetic Selection: Lessons from the South African Merino


Simple Summary
Abstract
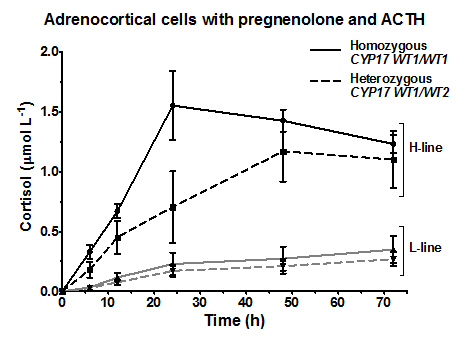
{kind=link}
{kind=link}
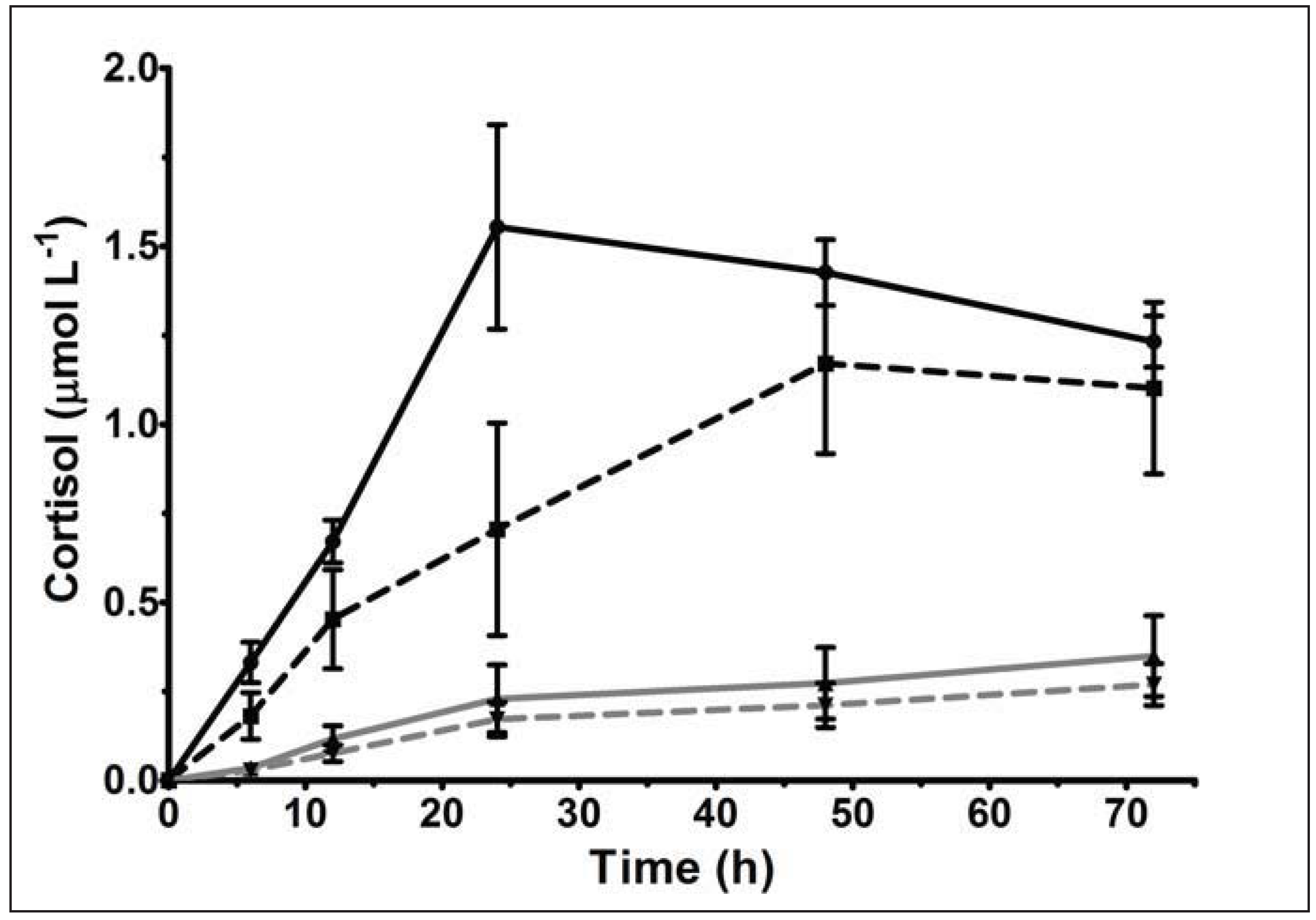{kind=link}
1. Introduction
2. Lessons from the South African Sheep Industry
2.1. Current Breeding Strategies in South Africa
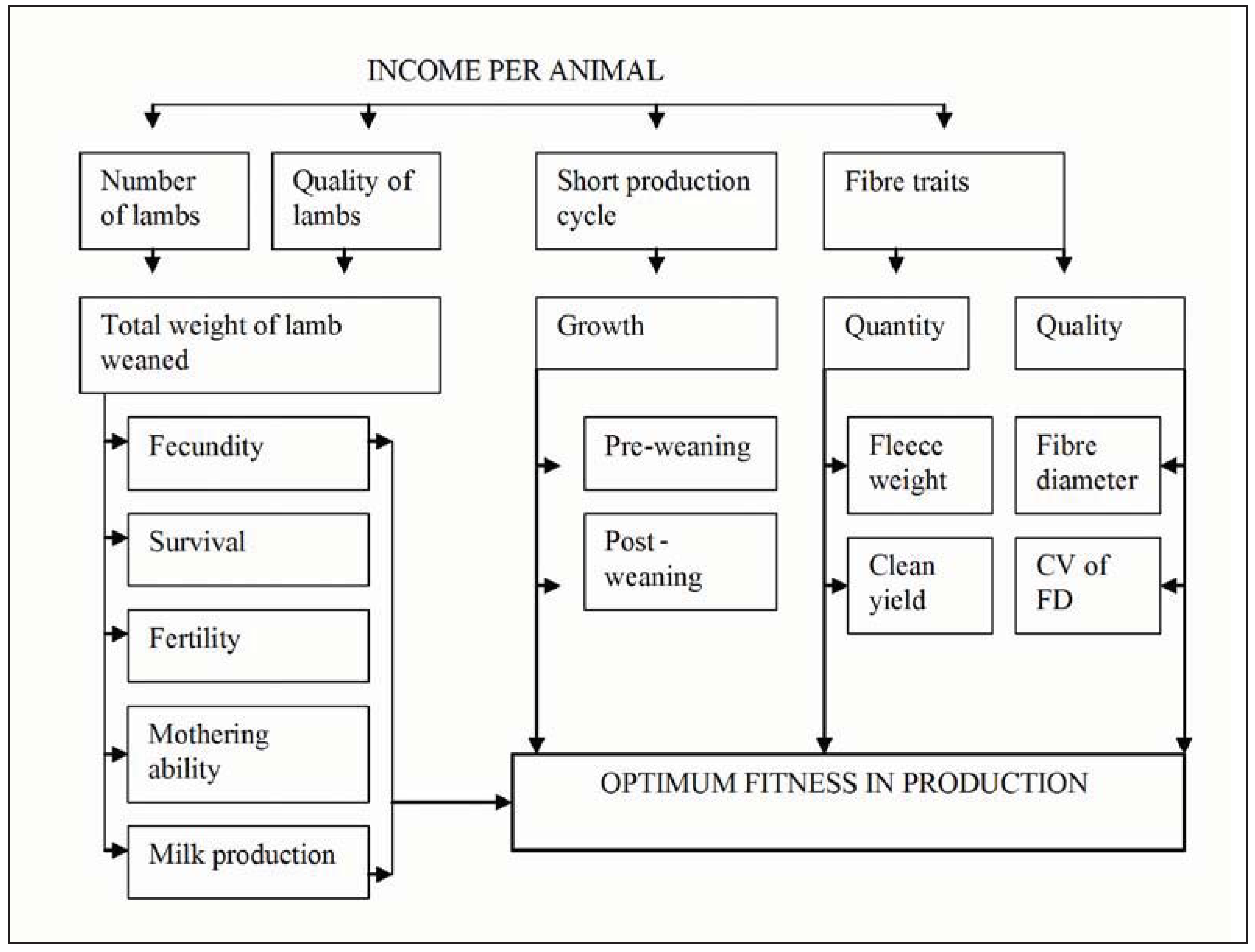
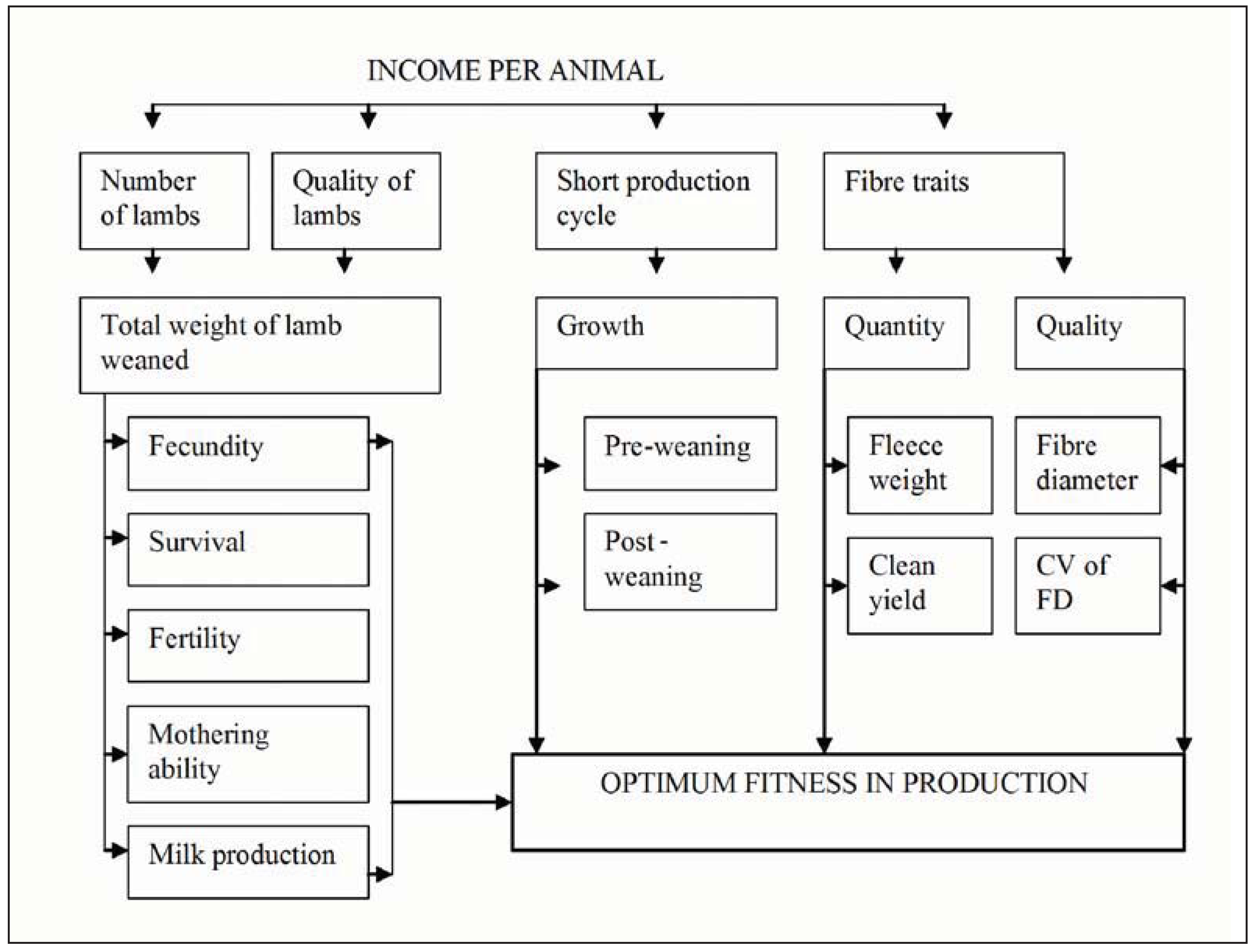
2.2. Selection Based on Maternal Ranking Values to Rear Multiple Offspring
3. Robustness as Breeding Goal: Utilization of HPAA Function
3.1. Introduction to Stress
3.2. Stress in Sheep
3.3. The Hypothalamic-Pituitary-Adrenal Axis
3.3.1. The Hypothalamus
3.3.2. The Pituitary
3.3.3. The Adrenal Gland
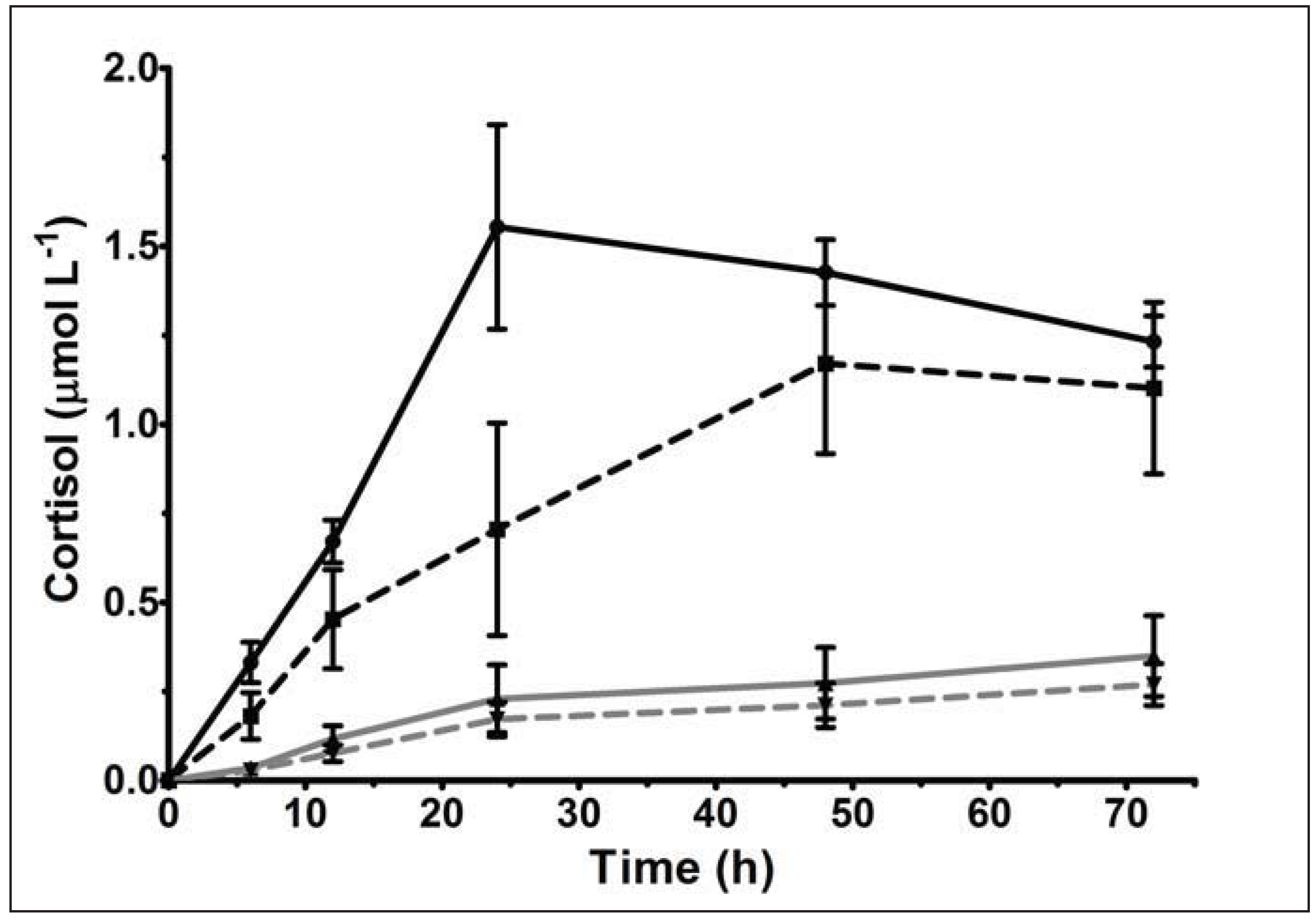
3.3.4. Extended Glucocorticoid Metabolism
3.4. Mechanism of Action of Glucocorticoids
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Mormède, P.; Foury, A.; Terenina, E.; Knap, P.W. Breeding for robustness: The role of cortisol. Animal 2011, 5, 651–657. [Google Scholar] [CrossRef]
- Knap, P.W.; Rauw, W.M. Selection for high production in pigs. In Resource Allocation Theory Applied to Farm Animal Production; Rauw, W.M., Ed.; CABI Publishing: Wallingford, UK, 2009; pp. 210–229. [Google Scholar]
- Knap, P.W. Breeding robust pigs. Austr. J. Exper. Agr. 2005, 45, 763–773. [Google Scholar] [CrossRef]
- Beilharz, R.G. Environmental limit to genetic change. An alternative theorem of natural selection. J. Anim. Breed. Genet. 1998, 115, 433–437. [Google Scholar] [CrossRef]
- Cloete, S.W.P.; Olivier, J.J. South African Sheep and Wool Industry. In The International Sheep and Wool Handbook; Cottle, D.J., Ed.; Nottingham University Press: Nottingham, UK, 2010; pp. 95–112. [Google Scholar]
- Rauw, W.M.; Kanis, E.; Noordhuizen-Stassen, E.N.; Grommers, F.J. Undesirable side effects of selection for high production efficiency in farm animals: A review. Livest. Prod. Sci. 1998, 56, 15–33. [Google Scholar] [CrossRef]
- Star, L.; Ellen, E.D.; Uitdehaag, K.; Brom, F.W.A. A plea to implement robustness into a breeding goal: Poultry as an example. J. Agr. Environ. Ethics 2008, 21, 109–125. [Google Scholar] [CrossRef]
- Siegel, P.B.; Honaker, C.F.; Rauw, W.M. Selection for high production in poultry. In Resource Allocation Theory Applied to Farm Animal Production; Rauw, W.M., Ed.; CABI Publishing: Wallingford, UK, 2009; pp. 230–242. [Google Scholar]
- Veerkamp, R.F.; Windig, J.J.; Calus, M.P.L.; Ouweltjes, W.; De Haas, Y.; Beerda, B. Selection for high production in dairy cattle. In Resource Allocation Theory Applied to Farm Animal Production; Rauw, W.M., Ed.; CABI Publishing: Wallingford, UK, 2009; pp. 243–260. [Google Scholar]
- Van Rensburg, S.J. Reproductive physiology and endocrinology of normal and habitually aborting Angora goats. Onderstepoort J. Vet. Res. 1971, 38, 1–62. [Google Scholar]
- Engelbrecht, Y.; Herselman, T.; Louw, A.; Swart, P. Adrenal function in Angora goats: A comparative study of adrenal steroidogenesis in Angora goats, Boer goats, and Merino sheep. J. Anim. Sci. 2000, 78, 371–379. [Google Scholar]
- Storbeck, K.; Swart, A.C.; Snyman, M.A.; Swart, P. Two CYP17 genes in the South African Angora goat (Capra hircus)—The identification of three genotypes that differ in copy number and steroidogenic output. FEBS J. 2008, 275, 3934–3943. [Google Scholar] [CrossRef]
- Department of Agriculture, Forestry and Fisheries. Abstract of Agricultural Statistics. 2012. Available online: http://www.nda.agric.za/docs/statsinfo/Ab2012.pdf (accessed on 9 May 2013).
- Cape Wools SA. Annual Report 2008/2009. Available online: http://www.capewools.co.za/index.php?option=com_docman&task=doc_download&gid=5353 (accessed on 20 February 2013).
- Lynch, J.J.; Hinch, G.N.; Adams, D.B. The Behavior of Sheep; CSIRO Publications: Melbourne, Australia, 1992; pp. 4–5, 51, 81–86,182–188. [Google Scholar]
- Schoeman, S.J.; Cloete, S.W.P.; Olivier, J.J. Returns on investment in sheep and goat breeding in South Africa. Livest. Sci. 2010, 130, 70–82. [Google Scholar] [CrossRef]
- Mason, I.L. A World Dictionary of Livestock Breeds, Types and Varieties, 2nd ed.; Farnham Royal: Buckinghamshire, UK, 1969. [Google Scholar]
- Olivier, J.J. The South African Merino performance testing. Proc. Assoc. Adv. Anim. Breed. Genet. 1999, 13, 119–124. [Google Scholar]
- Department of Agriculture, Forestry and Fisheries. Livestock Development Strategy for South Africa. Available online: http://www.nda.agric.za/docs/GenPub/DevStrat2007.pdf (accessed on 20 February 2013).
- Agricultural Research Council. Small Stock Improvement Scheme. Available online: http://www.arc.agric.za/home.asp?pid=2753 (accessed on 10 November 2011).
- Cloete, S.W.P.; Gilmour, A.R.; Olivier, J.J.; Van Wyk, J.B. Genetic and phenotypic trends and parameters in reproduction, greasy fleece weight and live weight in Merino lines divergently selected for multiple rearing ability. Austr. J. Exper. Agr. 2004, 44, 745–754. [Google Scholar] [CrossRef]
- Cloete, S.W.P.; Scholtz, A.J. Lamb survival in relation to lambing and neonatal behavior in medium wool Merino lines divergently selected for multiple rearing ability. Austr. J. Exper. Agr. 1998, 38, 801–811. [Google Scholar] [CrossRef]
- Cloete, S.W.P.; Scholtz, A.J.; Ten Hoope, J.M. A note on separation from one or more lambs in Merino lines divergently selected for ewe multiple rearing ability. Appl. Anim. Behav. Sci. 1998, 58, 189–195. [Google Scholar] [CrossRef]
- Cloete, S.W.P.; Scholtz, A.J.; Taljaard, R. Lambing behavior of Merino ewes from lines subjected to divergent selection for multiple rearing ability from the same base population. S. Afr. J. Anim. Sci. 2002, 32, 57–65. [Google Scholar]
- Cloete, S.W.P.; Scholtz, A.J.; Cloete, J.J.E.; Van Wyk, J.B. The ability of Merino ewes and lambs to reunite after separation, as affected by divergent selection for ewe multiple rearing capacity. Austr. J. Exper. Agr. 2005, 45, 1131–1137. [Google Scholar] [CrossRef]
- Cloete, S.W.P.; Olivier, J.J.; Van Wyk, J.B.; Erasmus, G.J.; Schoeman, S.J. Genetic parameters and trends for birth weight, birth coat score and weaning weight in Merino lines divergently selected for ewe multiple rearing ability. S. Afr. J. Anim. Sci. 2003, 33, 248–256. [Google Scholar]
- Cloete, S.W.P.; Misztal, I.; Olivier, J.J. Genetic parameters and trends for lamb survival and birth weight in a Merino flock divergently selected for multiple rearing ability. J. Anim. Sci. 2009, 87, 2196–2208. [Google Scholar] [CrossRef]
- Cloete, S.W.P.; Gilmour, A.R.; Olivier, J.J.; Van Wyk, J.B. Age trends in economically important traits of Merino ewes subjected to 10 years of divergent selection for multiple rearing ability. S. Afr. J. Anim. Sci 2003, 33, 43–51. [Google Scholar]
- Cloete, S.W.P.; Olivier, J.J. Direct and correlated responses to selection for multiple rearing ability in South African Merinos. Proc. Ann. Congr. S. Afr. Soc. Anim. Sci. 1998, 36, 65–68. [Google Scholar]
- Cloete, S.W.P.; Olivier, J.J.; Van Wyk, J.B.; Schoeman, S.J.; Erasmus, G.J. Genetic parameters and trends for hogget traits in Merino lines divergently selected for multiple rearing ability. Proc. Assoc. Adv. Anim. Breed. Genet. 2005, 16, 24–27. [Google Scholar]
- Scholtz, A.J.; Cloete, S.W.P.; Van Wyk, J.B.; Kruger, A.C.M.; Van der Linde, T.C.deK. Influence of divergent selection for reproduction on the occurrence of breech strike in mature Merino ewes. Anim. Prod. Sci. 2010, 50, 203–209. [Google Scholar] [CrossRef]
- Seddon, H.R. Conditions which predispose sheep to blowfly attack. Agr. Gazette New South Wales 1931, 42, 581–594. [Google Scholar]
- Seddon, H.R.; Belschner, H.G.; Mulhearn, C.R. Studies on Cutaneous Myiasis of Sheep (Sheep Blowfly Attack); Science Bulletin No. 37; New South Wales Department of Agriculture: Beecroft, Australia, 1931; pp. 3–30. [Google Scholar]
- Tillyard, R.J.; Seddon, H.R.; Council for Scientific and Industrial Research; Joint Blowfly Committee (Australia). The Sheep Blowfly Problem in Australia; Report No. 1; Council for Scientific and Industrial Research: Melbourne, Australia, 1933. [Google Scholar]
- Dun, R.B. Skin folds and Merino breeding. 1. The net reproductive rates of flocks selected for and against skin fold. Austr. J. Exper. Agr. Anim. Husbandry 1964, 4, 376–385. [Google Scholar] [CrossRef]
- McGuirk, B.J. Skin folds and Merino breeding. 8. Fertility of individual rams in flocks selected for high and low skin fold. Austr. J. Exper. Agr. Anim. Husbandry 1969, 9, 147–150. [Google Scholar] [CrossRef]
- Atkins, K.D. Selection for skin folds and fertility. Skin folds as a breeding objective. Proc. Austr. Soc. Anim. Prod. 1980, 13, 174–176. [Google Scholar]
- Cloete, J.J.E.; Cloete, S.W.P.; Hoffman, L.C. Behavior of Merinos divergently selected for multiple rearing ability in response to external stimuli. Small Ruminant Res. 2005, 60, 227–236. [Google Scholar] [CrossRef]
- Hoffman, L.C.; Schmidt, D.; Muller, M.M.; Cloete, J.J.E.; Cloete, S.W.P. Sensory and objective mutton quality characteristics of SA Merino sheep selected for and against reproductive fitness. S. Afr. J. Anim. Sci. 2003, 33, 52–64. [Google Scholar]
- Van der Walt, D.; Cloete, S.W.P.; Storbeck, K.; Swart, P. The role of cytochrome P450 17α-hydroxylase/17,20-lyase (CYP17) in the stress coping ability in a divergently selected Merino sheep population. Proc. Assoc. Adv. Anim. Breed. Genet. 2009, 18, 100–103. [Google Scholar]
- Hough, D.; Storbeck, K.; Cloete, S.W.P.; Swart, P. Improving stress coping ability: Comparison between the CYP17 genotype of Ovis aries and Capra hircus. In Proceedings of the 9th World Congress on Genetics Applied to Livestock Production, Leipzig, Germany, 1–6 August 2010.
- Hough, D. Comparison of two CYP17 isoforms: Implications for cortisol production in the South African Merino. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2012. [Google Scholar]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics, 4th ed.; Longman: New York, NY, USA, 1996. [Google Scholar]
- Mackay, T.F.C. The nature of quantitative genetic variation revisited: Lessons from Drosophila bristles. BioEssays 1996, 18, 113–121. [Google Scholar] [CrossRef]
- Mackay, T.F.C. Quantitative trait loci in Drosophila. Nat. Rev. Genet. 2001, 2, 11–20. [Google Scholar] [CrossRef]
- Camp, N.J.; Cox, A. Methods in Molecular Biology. In Quantitative Trait Loci: Methods and Protocols; Humana Press, Inc.: Totowa, NJ, USA, 2002; Volume 195. [Google Scholar]
- Wu, R.; Ma, C.; Casella, G. Statistical Genetics of Quantitative Traits; Springer Science and Business Media: New York, NY, USA, 2007. [Google Scholar]
- DeRijk, R.H. Single nucleotide polymorphisms related to HPA axis activity. Neuroimmunomodulation 2009, 16, 340–352. [Google Scholar]
- Bartels, M.; Berg, M.V.D.; Sluyter, F.; Boomsma, D.I.; Geus, E.J.D. Heritability of cortisol levels: Review and simultaneous analysis of twin studies. Psychoneuroendocrinology 2003, 28, 121–137. [Google Scholar] [CrossRef]
- Federenko, I.S.; Nagamine, M.; Hellhammer, D.H.; Wadhwa, P.D.; Wust, S. The heritability of hypothalamus pituitary adrenal axis responses to psychosocial stress is context dependent. J. Clin. Endocrinol. Metabol. 2004, 89, 6244–6250. [Google Scholar] [CrossRef]
- Guimont, F.S.; Wynne-Edwards, K.E. Individual variation in cortisol responses to acute “on-back” restraint in an outbred hamster. Hormone. Behav. 2006, 50, 252–260. [Google Scholar] [CrossRef]
- You, Q.; Karrow, N.A.; Cao, H.; Rodriguez, A.; Mallard, B.A.; Boermans, H.J. Variation in the ovine cortisol response to systemic bacterial endotoxin challenge is predominantly determined by signaling within the hypothalamic-pituitary-adrenal axis. Toxicol. Appl. Pharmacol. 2008, 230, 1–8. [Google Scholar] [CrossRef]
- DeRijk, R.; Kloet, E.D. Corticosteroid receptor genetic polymorphisms and stress responsivity. Endocrine 2005, 28, 263–270. [Google Scholar] [CrossRef]
- Wüst, S.; Rossum, E.F.C.V.; Federenko, I.S.; Koper, J.W.; Kumsta, R.; Hellhammer, D.H. Common polymorphisms in the glucocorticoid receptor gene are associated with adrenocortical responses to psychosocial stress. J. Clin. Endocrinol. Metabol. 2004, 89, 565–573. [Google Scholar] [CrossRef]
- Xu, D.; Buehner, A.; Xu, J.; Lambert, T.; Nekl, C.; Nielsen, M.K.; Zhou, Y. A polymorphic glucocorticoid receptor in a mouse population may explain inherited altered stress response and increased anxiety-type behaviors. FASEB J. 2006, 20, 2414–2416. [Google Scholar] [CrossRef]
- Hough, D.; Cloete, S.W.P.; Storbeck, K.; Swart, A.C.; Swart, P. Cortisol production in sheep is influenced by the functional expression of two cytochrome P450 17α-hydroxylase/17,20-lyase (CYP17) isoforms. J. Anim. Sci. 2013, 91, 1193–1206. [Google Scholar] [CrossRef]
- Meaney, M.J.; Szyf, M. Environmental programming of stress responses through DNA methylation: Life at the interface between a dynamic environment and a fixed genome. Dial. Clin. Neurosci. 2005, 7, 103–123. [Google Scholar]
- Weaver, I.C.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behavior. Nat. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef]
- SanCristobal-Gaudy, M.; Bodin, L.; Elsen, J.M.; Chevalet, C. Genetic components of litter size variability in sheep. Genet. Sel. Evol. 2001, 33, 249–271. [Google Scholar] [CrossRef]
- Foury, A.; Geverink, N.A.; Gil, M.; Gispert, M.; Hortos, M.; Font i Furnols, M.; Carrion, D.; Blott, S.C.; Plastow, G.S.; Mormède, P. Stress neuroendocrine profiles in five pig breeding lines and the relationship with carcass composition. Animal 2007, 1, 973–982. [Google Scholar] [CrossRef]
- Mormède, P.; Andanson, S.; Aupérin, B.; Beerda, B.; Guémené, D.; Malmkivist, J.; Manteca, X.; Manteuffel, G.; Prunet, P.; Van Reenen, C.G.; Richard, S.; Veissier, I. Exploration of the hypothalamic-pituitary-adrenal function as a tool to evaluate animal welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef]
- Selye, H. The general adaptation syndrome and the diseases of adaptation. J. Clin. Endocrinol. 1946, 6, 117–230. [Google Scholar] [CrossRef]
- Ewbank, R. The behavioral needs of farm and laboratory animals. In Animal Experimentation: Improvements and Alternatives; Marsh, N., Haywood, S., Eds.; FRAME: Nottingham, UK, 1985; pp. 31–35. [Google Scholar]
- Dickens, M.J.; Delehanty, D.J.; Romero, L.M. Stress: An inevitable component of animal translocation. Biol. Conserv. 2010, 143, 1329–1341. [Google Scholar] [CrossRef]
- Wingfield, J.C.; Maney, D.L.; Breuner, C.W.; Jacobs, J.D.; Lynn, S.; Ramenofsky, M.; Richardson, R.D. Ecological bases of hormone-behavior interactions: The emergency life history stage. Am. Zool. 1998, 38, 191–206. [Google Scholar]
- Nazifi, S.; Saeb, M.; Rowghani, E.; Kaveh, K. The influences of thermal stress on serum biochemical parameters of Iranian fat-tailed sheep and their correlation with triiodothyronine (T3), thyroxine (T4) and cortisol concentrations. Comp. Clin. Pathol. 2003, 12, 135–139. [Google Scholar] [CrossRef]
- Michel, V.; Peinnequin, A.; Alonso, A.; Buguet, A.; Cespuglio, R.; Canini, F. Decreased heat tolerance is associated with hypothalamic-pituitary-adrenocortical axis impairment. Neuroscience 2007, 147, 522–531. [Google Scholar] [CrossRef]
- Knap, P.W. Robustness. In Resource Allocation Theory Applied to Farm Animal Production; Rauw, W.M., Ed.; CABI Publishing: Wallingford, UK, 2009; pp. 288–301. [Google Scholar]
- Wingfield, J.C.; Romero, L.M. Adrenocortical responses to stress and their modulation in free-living vertebrates. In Handbook of Physiology, Section 7: The Endocrine System; McEwen, B.S., Goodman, H.M., Eds.; Oxford University Press: New York, NY, USA, 2001; Volume IV, pp. 211–234. [Google Scholar]
- Spraker, T.R.; Hibler, C.P.; Schoonveld, G.G.; Adney, W.S. Pathologic changes and microorganisms found in Bighorn sheep during a stress-related die-off. J. Wildl. Dis. 1984, 20, 319–327. [Google Scholar]
- Kilgour, R. Sheep behavior: Its importance in farming systems, handling, transport and pre-slaughter treatment. In Proceedings of Sheep Assembly and Transport Workshop: A Workshop to Study the Supply, Holding and Transport of Sheep for Abattoir Slaughter or Live Transport, Perth, Australia, October 1976; pp. 64–84.
- Degabrielle, R.; Fell, L.R. Changes in behavior, cortisol and lymphocyte types during isolation and group confinement of sheep. Immunol. Cell Biol. 2001, 79, 583–589. [Google Scholar] [CrossRef]
- Da Costa, A.P.; Leigh, A.E.; Man, M.; Kendrick, K.M. Face pictures reduce behavioral, autonomic, endocrine and neural indices of stress and fear in sheep. Proc. Roy. Soc. Lond. B 2004, 271, 2077–2084. [Google Scholar] [CrossRef]
- Grandin, T. Assessment of stress during handling and transport. J. Anim. Sci. 1997, 75, 249–257. [Google Scholar]
- Hargreaves, A.L.; Hutson, G.D. The stress response in sheep during routine handling procedures. Appl. Anim. Behav. Sci. 1990, 26, 83–90. [Google Scholar]
- Alexander, G. Constraints to lambs’ survival. In Reproduction in Sheep; Lindsay, D.R., Pearce, D.T., Eds.; Australian Academy of Science and the Australian Wool Corporation: Canberra, Australia, 1984; pp. 199–209. [Google Scholar]
- Murphy, P.M.; Purvis, I.W.; Lindsay, D.R.; LeNeindre, P.; Orgeur, P.; Poindron, P. Measures of temperament are highly repeatable in Merino sheep and some are related to maternal behavior. Proc. Austr. Soc. Anim. Prod. 1994, 20, 247–250. [Google Scholar]
- Murphy, P.M. Maternal behavior and rearing ability of Merino ewes can be improved by strategic feed supplementation during late pregnancy and selection for calm temperament. Ph.D. Thesis, The University of Western Australia, Perth, Australia, 1999. [Google Scholar]
- Pryce, C.R.; Abbott, A.H.; Hodges, J.K.; Martin, R.D. Maternal behavior is related to prepartum urinary estradiol levels in the red-bellied tamarin monkeys. Physiol. Behav. 1988, 44, 717–726. [Google Scholar] [CrossRef]
- Dwyer, C.M.; Gilbert, C.L.; Lawrence, A.B. Prepartum plasma estradiol and postpartum cortisol, but not oxytocin, are associated with interindividual and breed differences in the expression of maternal behavior in sheep. Hormone. Behav. 2004, 46, 529–543. [Google Scholar] [CrossRef]
- Bickell, S.; Nowak, R.; Poindron, P.; Chadwick, A.; Ferguson, D.; Blache, D. Challenge by a novel object does not impair the capacity of ewes and lambs selected for a nervous temperament to display early preference for each other. Anim. Prod. Sci. 2011, 51, 575–581. [Google Scholar] [CrossRef]
- Kilgour, R.J.; Szantar-Coddington, M.R. Arena behavior of ewes selected for superior mothering ability differs from that of unselected ewes. Anim. Reprod. Sci. 1995, 37, 133–141. [Google Scholar] [CrossRef]
- Rivier, C.; Rivest, S. Effect of stress on the activity of the hypothalamic-pituitary-gonadal axis: Peripheral and central mechanisms. Biol. Reprod. 1991, 45, 523–532. [Google Scholar] [CrossRef]
- Manteuffel, G. Central nervous regulation of the hypothalamic-pituitary-adrenal axis and its impact on fertility, immunity, metabolism and animal welfare, a review. Archiv fur Tierzucht 2002, 45, 575–595. [Google Scholar]
- Cook, N.J. Minimally invasive sampling media and the measurement of corticosteroids as biomarkers of stress in animals. Can. J. Anim. Sci. 2011, 92, 227–259. [Google Scholar] [CrossRef]
- Mormede, P.; Foury, A.; Barat, P.; Corcuff, J.B.; Terenina, E.; Marissal-Arvy, N.; Moisan, M.P. Molecular genetics of hypothalamic-pituitary-adrenal axis activity and function. Ann. New York Acad. Sci. 2011, 1220, 127–136. [Google Scholar] [CrossRef]
- Weitzman, E.D.; Fukushima, D.; Nogeire, C.; Roffwarg, H.; Gallagher, T.F.; Hellman, L. Twenty-four hour pattern of the episodic secretion of cortisol in normal subjects. J. Clin. Endocrinol. Metabol. 1971, 33, 14–22. [Google Scholar] [CrossRef]
- Whitnall, M.H. Regulation of the hypothalamic corticotropin-releasing hormone neurosecretory system. Progr. Neurobiol. 1993, 40, 573–629. [Google Scholar] [CrossRef]
- Aguilera, G. Corticotropin releasing hormone, receptor regulation and the stress response. Trends Endocrinol. Metabol. 1998, 9, 329–336. [Google Scholar] [CrossRef]
- Lightman, S.L. How does the hypothalamus respond to stress? Neurosciences 1994, 6, 215–219. [Google Scholar]
- Bonaz, B.; Rivest, S. Effect of chronic stress on CRF neuronal activity and expression of its type 1 receptor in the rat brain. Am. J. Physiol. 1998, 44, 1438–1449. [Google Scholar]
- Blache, D.; Bickell, S.L. Temperament and reproductive biology: Emotional reactivity and reproduction in sheep. Rev. Bras. Zootecn. 2010, 39, 401–408. [Google Scholar] [CrossRef]
- Keller, M.; Meurisse, M.; Levy, F. Mapping the neural substrates involved in maternal responsiveness and lamb olfactory memory in parturient ewes using Fos imaging. Behav. Neurosci. 2004, 118, 1271–1284. [Google Scholar]
- Novak, R.; Keller, M.; Lévy, F. Mother-young relationships in sheep: A model for a multidisciplinary approach of the study of attachment in mammals. J. Neuroendocrinol. 2011, 23, 1042–1053. [Google Scholar] [CrossRef]
- Vander, A.; Sherman, J.; Luciano, D. Human Physiology, the Mechanisms of Body Function, 9th ed.; McGraw-Hill Companies Inc.: New York, NY, USA, 2004. [Google Scholar]
- Von Borell, E.; Dobson, H.; Prunier, A. Stress, behavior and reproductive performance in female cattle and pigs. Hormone. Behav. 2007, 52, 130–138. [Google Scholar] [CrossRef]
- Dunn, A.F.; Berridge, C.W. Physiological and behavioral responses to corticotropin-releasing factor administration: Is CRF a mediator of anxiety or stress responses? Brain Res. Rev. 1990, 15, 71–100. [Google Scholar] [CrossRef]
- Chen, F.M.; Bilezikjian, L.M.; Perrin, M.H.; Rivier, J.; Vale, W. Corticotropin releasing factor receptor-mediated stimulation of adenylate cyclase activity in the rat brain. Brain Res. 1986, 381, 49–57. [Google Scholar] [CrossRef]
- Familari, M.; Smith, A.I.; Smith, R.; Funder, J.W. Arginine vasopressin is a much more potent stimulus to ACTH release from ovine anterior pituitary cells than ovine corticotrophin-releasing factor: In vitro studies. Neuroendocrinology 1989, 50, 152–157. [Google Scholar] [CrossRef]
- Owens, M.J.; Nemeroff, C.B. Physiology and pharmacology of corticotropin-releasing factor. Pharmacol. Rev. 1992, 43, 425–436. [Google Scholar]
- Liu, J.; Robinson, P.J.; Funder, J.W.; Engler, D. A comparative study of the role of adenylate cyclase in the release of adrenocorticotropin from the ovine and rat anterior pituitary. Mol. Cell. Endocrinol. 1994, 101, 173–181. [Google Scholar] [CrossRef]
- Mastorakos, G.; Weber, J.S.; Magiakou, M.A.; Gunn, H.; Chrousos, G.P. Hypothalamic-pituitary-adrenal axis evaluation and stimulation of systemic vasopressin secretion by recombinant interleukin 6 in humans: Potential implications for the syndrome of inappropriate vasopressin secretion. J. Clin. Endocrinol. Metabol. 1994, 79, 934–939. [Google Scholar] [CrossRef]
- Charmandari, E.; Tsigos, C.; Chrousos, G. Endocrinology of the stress response. Ann. Rev. Physiol. 2005, 67, 259–284. [Google Scholar] [CrossRef]
- Stevens, A.; Begum, G.; Cook, A.; Connor, K.; Rumball, C.; Oliver, M.; Challis, J.; Bloomfield, F.; White, A. Epigenetic changes in the hypothalamic proopiomelanocortin and glucocorticoid receptor genes in the ovine fetus after periconceptional undernutrition. Endocrinology 2010, 151, 3652–3664. [Google Scholar] [CrossRef]
- McMillen, I.C.; Phillips, I.D.; Ross, J.T.; Robinson, J.S.; Owens, J.A. Chronic stress: The key to parturition? Reprod. Fert. Develop. 1995, 7, 499–507. [Google Scholar] [CrossRef]
- Schwartz, J.; Ash, P.; Ford, V.; Raff, H.; Crosby, S.; White, A. Secretion of adrenocorticotrophin (ACTH) and ACTH precursors in ovine anterior pituitary cells: Actions of corticotrophinreleasing hormone, arginine vasopressin and glucocorticoids. J. Endocrinol. 1994, 140, 189–195. [Google Scholar] [CrossRef]
- Challis, G., Jr.; Brooks, A.N. Maturation and activation of hypothalamic-pituitary-adrenal function in foetal sheep. Endocrine Rev. 1989, 10, 182–204. [Google Scholar] [CrossRef]
- Miller, W.L.; Auchus, R.J. The molecular biology, biochemistry, and physiology of human steroidogenesis and its disorders. Endocrine Res. 2011, 32, 81–151. [Google Scholar] [CrossRef]
- Gwynne, J.T.; Strauss, J.F, 3rd. The role of lipoproteins in steroidogenesis and cholesterol metabolism in steroidogenic glands. Endocrine Rev. 1982, 3, 299–329. [Google Scholar] [CrossRef]
- Jefcoate, C. High-flux mitochondrial cholesterol trafficking, a specialized function of the adrenal cortex. J. Clin. Invest. 2002, 110, 881–890. [Google Scholar] [Green Version]
- Brown, M.S.; Kovanen, P.T.; Goldstein, J.L. Receptor mediated uptake of lipoprotein-cholesterol and its utilization for steroid synthesis in the adrenal cortex. Recent Prog. Hormone Res. 1979, 35, 215–257. [Google Scholar]
- Liu, J.; Heikkila, P.; Meng, Q.H.; Kahri, A.I.; Tikkanen, M.J.; Voutilainen, R. Expression of low and high density lipoprotein receptor genes in human adrenals. Eur. J. Endocrinol. 2000, 142, 677–682. [Google Scholar] [CrossRef]
- Mason, J.I.; Rainey, W.E. Steroidogenesis in the human foetal adrenal: A role for cholesterol synthesized de novo. J. Clin. Endocrinol. Metabol. 1987, 64, 140–147. [Google Scholar] [CrossRef]
- Kraemer, F.B. Adrenal cholesterol utilization. Mol. Cell. Endocrinol. 2007, 265, 42–45. [Google Scholar] [CrossRef]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Invest. 2002, 109, 1125–1131. [Google Scholar]
- Juengel, J.L.; Meberg, B.M.; Turzillo, A.M.; Nett, T.M.; Niswender, G.D. Hormonal regulation of messenger ribonucleic acid encoding steroidogenic acute regulatory protein in ovine corpora lutea. Endocrinology 1995, 136, 5423–5429. [Google Scholar] [CrossRef]
- Hogg, K.; McNeilly, A.S.; Duncan, W.C. Prenatal androgen exposure leads to alterations in gene and protein expression in the ovine foetal ovary. Endocrinology 2011, 152, 2048–2059. [Google Scholar] [CrossRef]
- Storbeck, K.; Kolar, N.W.; Stander, M; Swart, A.C.; Prevoo, D.; Swart, P. The development of an ultra performance liquid chromatography-coupled atmospheric pressure chemical ionization mass spectrometry assay for seven adrenal steroids. Anal. Biochem. 2008, 37, 11–20. [Google Scholar]
- Boon, W.C.; Roche, P.J.; Butkus, A.; McDougall, J.G.; Jeyaseelan, K.; Coghlan, P. Functional and expression analysis of ovine steroid 11β-hydroxylase (cytochrome P45011β). Endocrine Res. 1997, 23, 325–347. [Google Scholar] [CrossRef]
- Imai, T.; Yamazaki, T.; Kominami, S. Kinetic studies on bovine cytochrome P45011B catalyzing successive reactions from 11-deoxycorticosterone to aldosterone. Biochemistry 1998, 37, 8097–8104. [Google Scholar] [CrossRef]
- Lisurek, M.; Bernhardt, R. Modulation of aldosterone and cortisol synthesis on the molecular level. Mol. Cell. Endocrinol. 2004, 215, 149–159. [Google Scholar] [CrossRef]
- Ehrhart-Bornstein, M.; Hinson, J.P.; Bornstein, S.R.; Scherbaum, W.A.; Vinson, G.P. Intraadrenal interactions in the regulation of adrenocortical steroidogenesis. Endocrine Rev. 1998, 19, 101–143. [Google Scholar] [CrossRef]
- Hinson, J.P.; Kapas, S. The role of endothelial cell products in the regulation of adrenocortical function: Actions of endothelin, nitric oxide, adrenomedullin and PAMP. Hormone Metab. Res. 1998, 30, 334–340. [Google Scholar] [CrossRef]
- Young, B.; Lowe, J.S.; Stevens, A.; Heath, J.W. Organ systems. In Wheather's Functional Histology: A Text and Colour Atlas, 5th ed.; Ozols, I., Whitehouse, A., Eds.; Churchill Livingstone Elsevier Ltd.: London, UK, 2006; pp. 331–341. [Google Scholar]
- Kempna, P.; Hirch, A.; Hofer, G.; Mullis, P.E.; Flück, C.E. Impact of differential P450c17 phosphorylation by cAMP stimulation and by starvation conditions on enzyme activities and androgen production in NCI-H295R Cells. Endocrinology 2010, 151, 3686–3696. [Google Scholar] [CrossRef]
- Rainey, W.E.; Shay, J.W.; Mason, J.I. The effect of cytochalasin D on steroid production and stress fiber organization in cultured bovine adrenocortical cells. Mol. Cell. Endocrinol. 1984, 35, 189–197. [Google Scholar] [CrossRef]
- Voorhees, H.; Aschenbrenner, J.; Carnes, J.; Mrotek, J. Rounding and steroidogenesis of enzyme- and ACTH-treated Y-1 mouse adrenal tumor cells. Cell Biol. Int. Rep. 1984, 8, 483–497. [Google Scholar] [CrossRef]
- Rainey, W.E.; Kramer, R.E.; Mason, J.I.; Shay, J.W. The effects of taxol, a microtubule-stabilizing drug, on steroidogenic cells. J. Cell. Physiol. 1985, 123, 17–24. [Google Scholar] [CrossRef]
- Sackett, D.L.; Wolff, J. Cyclic AMP-independent stimulation of steroidogenesis in Y-1 adrenal tumor cells by antimitotic agents. Biochim. Biophys. Acta 1986, 888, 163–170. [Google Scholar] [CrossRef]
- Denkova, R.; Ivanov, I.; Dimitrova, M. Microtubules and regulation of granulosa cell steroidogenesis by porcine granulosa cell conditioned medium. Endocrine Regulat. 1992, 26, 195–199. [Google Scholar]
- Shiver, T.M.; Sackett, D.L.; Knipling, L.; Wolff, J. Intermediate filaments and steroidogenesis in adrenal Y-1 cells: Acrylamide stimulation of steroid production. Endocrinology 1992, 131, 201–207. [Google Scholar] [CrossRef]
- Han, J.D.; Rubin, C.S. Regulation of cytoskeleton organization and paxillin dephosphorylation by cAMP. J. Biol. Chem. 1996, 271, 29211–29215. [Google Scholar]
- Hall, P.F.; Almahbobi, G. Roles of microfilaments and intermediate filaments in adrenal steroidogenesis. Microsc. Res. Tech. 1997, 36, 463–479. [Google Scholar] [CrossRef]
- Lee, L.J.; Chen, J.S.; Ko, T.L.; Wang, S.M. Mechanism of colchicineinduced steroidogenesis in rat adrenocortical cells. J. Cell. Biochem. 2001, 81, 162–171. [Google Scholar]
- Whitehouse, B.J.; Gyles, S.L.; Squires, P.E.; Sayed, S.B.; Burnes, C.J.; Persaud, S.J.; Jones, P.M. Interdependence of steroidogenesis and shape changes in Y1 adrenocortical cells: Studies with inhibitors of phosphoprotein phosphatases. J. Endocrinol. 2002, 172, 583–593. [Google Scholar] [CrossRef]
- Nan, X.; Potma, E.O.; Xie, X.S. Nonperturbative chemical imaging of organelle transport in living cells with coherent anti-strokes Raman scattering microscopy. Biophys. J. 2006, 91, 728–735. [Google Scholar] [CrossRef]
- Sewer, M.B.; Li, D. Regulation of steroid hormone biosynthesis by the cytoskeleton. Lipids 2008, 43, 1109–1115. [Google Scholar] [CrossRef]
- Li, D.; Sewer, M.B. RhoA and DIAPH1 mediate adrenocorticotropin-stimulated cortisol biosynthesis by regulating mitochondrial trafficking. Endocrinology 2010, 151, 4313–4323. [Google Scholar] [CrossRef]
- Payne, A.H.; Hales, D.B. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocrine Rev. 2004, 25, 947–970. [Google Scholar] [CrossRef]
- Sirianni, R.; Rehman, K.S.; Carr, B.R.; Parker, C.R., Jr.; Rainey, W.E. Corticotropin-releasing hormone directly stimulates cortisol and the cortisol biosynthetic pathway in human foetal adrenal cells. J. Clin. Endocrinol. Metab. 2005, 90, 279–285. [Google Scholar]
- Xing, Y.; Edwards, M.A.; Ahlem, C.; Kennedy, M.; Cohen, A.; Gomez-Sanchez, C.E.; Rainey, W.E. The effects of ACTH on steroid metabolomic profiles in human adrenal cells. J. Endocrinol. 2011, 209, 327–335. [Google Scholar] [CrossRef]
- Phillips, I.D.; Ross, J.T.; Young, I.R.; McMillen, I.C. Adrenal steroidogenic enzyme expression in the hypothalamopituitary disconnected fetal sheep. Proc. Austr. Soc. Med. Res. 1994, 33, 150. [Google Scholar]
- Colomer, C.; Desarménien, M.G.; Guérineau, N.C. Revisiting the stimulus-secretion coupling in the adrenal medulla: Role of gap junction-mediated intercellular communication. Mol. Neurobiol. 2009, 40, 87–100. [Google Scholar] [CrossRef]
- Turnbull, A.V.; Prehar, S.; Kennedy, A.R.; Little, R.A.; Hopkins, S.J. Interleukin-6 is an afferent signal to the hypothalamo-pituitary-adrenal axis during local inflammation in mice. Endocrinology 2003, 144, 1894–1906. [Google Scholar] [CrossRef]
- Karrow, N.A. Activation of the hypothalamic-pituitary-adrenal axis and autonomic nervous system during inflammation and altered programming of the neuroendocrine-immune axis during fetal and neonatal development: Lessons learned from the model inflammagen, lipopolysaccharide. Brain Behav. Immun. 2006, 20, 144–158. [Google Scholar] [CrossRef]
- Muller-Steinhardt, M.; Ebel, B.; Hartel, C. The impact of interleukin-6 promoter-597/-572/-174genotype on interleukin-6 production after lipopolysaccharide stimulation. Clin. Exper. Immunol. 2007, 147, 339–345. [Google Scholar]
- Chrousos, G.P. The hypothalamic-pituitary-adrenal axis and immune-mediated inflammation. N. Engl. J. Med. 1995, 332, 1351–1362. [Google Scholar] [CrossRef]
- Campbell, L.E.; Yang, M.Y.K. Ovine 11β-hydroxysteroid dehydrogenase type 2 gene predicts a protein distinct from that deduced by the cloned kidney cDNA at the C-terminus. Mol. Cell. Endocrinol. 1996, 119, 113–118. [Google Scholar] [CrossRef]
- Simmons, R.M.; Satterfield, M.C.; Welsh, T.H., Jr.; Brazer, F.W.; Spencer, T.E. HSD11B1, HSD11B2, PTGS2, and NR3C1 expression in the peri-implantation ovine uterus: Effects of pregnancy, progesterone and interferon tau. Biol. Reprod. 2010, 82, 35–43. [Google Scholar] [CrossRef]
- Quinkler, M.; Stewart, P.M. Hypertension and the cortisol-cortisone shuttle. J. Clin. Endocrinol. Metab. 2003, 88, 2384–2392. [Google Scholar] [CrossRef]
- Smith, C.L.; Hammond, G.L. Hormonal regulation of corticosteroid-binding globulin biosynthesis in the male rat. Endocrinology 1992, 130, 2245–2251. [Google Scholar] [CrossRef]
- Mihrshahi, R.; Lewis, J.G.; Ali, S.O. Hormonal effects on the secretion and glycoform profile of corticosteroid-binding globulin. J. Steroid Biochem. Mol. Biol. 2006, 101, 275–285. [Google Scholar] [CrossRef]
- Duma, D.; Jewell, C.M.; Cidlowski, J.A. Multiple glucocorticoid receptor isoforms and mechanisms of post-translational modification. J. Steroid Biochem. Mol. Biol. 2006, 102, 11–21. [Google Scholar] [CrossRef]
- Hantzis, V.; Albiston, A.; Matsacos, D.; Wintour, E.M.; Peers, A.; Koukoulas, I.; Myles, K.; Moritz, K.; Dodic, M. Effect of early glucocorticoid treatment on MR and GR in late gestation ovine kidney. Kidney Int. 2002, 61, 405–413. [Google Scholar] [CrossRef]
- Oakley, R.H.; Webster, J.C.; Sar, M.; Parker, C.R., Jr.; Cidlowski, J.A. Expression and subcellular distribution of the beta-isoform of the human glucocorticoid receptor. Endocrinology 1997, 138, 5028–5038. [Google Scholar] [CrossRef]
- Beato, M.; Truss, M.; Chavez, S. Control of transcription by steroid hormones. Ann. N. Y. Acad. Sci. 1996, 784, 93–123. [Google Scholar] [CrossRef]
- Olefsky, J.M. Nuclear receptor minireview series. J. Biol. Chem. 2001, 276, 36836–36864. [Google Scholar] [CrossRef]
- Newton, R. Molecular mechanisms of glucocorticoid action: What is important? Thorax 2000, 55, 603–613. [Google Scholar] [CrossRef]
- Barouki, R.; Chobert, M.N.; Billon, M.C.; Finidori, J.; Tsapis, R.; Hanoune, J. Glucocorticoid hormones increase the activity of gamma-glutamyltransferase in a highly differentiated hepatoma cell line. Biochim. Biophys. Acta 1982, 721, 11–21. [Google Scholar] [CrossRef]
- Schmid, E.; Schmid, W.; Jantzen, M.; Mayer, D.; Jastorff, B.; Schutz, G. Transcription activation of the tyrosine aminotransferase gene by glucocorticoids and cAMP in primary hepatocytes. Eur. J. Biochem. 1987, 165, 499–506. [Google Scholar] [CrossRef]
- Orth, D.N.; Kovacs, W.J.; DeBold, C.R. The adrenal cortex. In Textbook of Endocrinology; Wilson, J.D., Foster, D.W., Eds.; WB Saunders Company: Philadelphia, PA, USA, 1992; pp. 489–619. [Google Scholar]
- DuBois, D.C.; Xu, Z.X.; McKay, L.; Almon, R.R.; Pyszcznski, N.; Jusko, W.J. Differential dynamics of receptor down-regulation and tyrosine aminotransferase induction following glucocorticoid treatment. J. Steroid Biochem. Mol. Biol. 1995, 54, 237–243. [Google Scholar] [CrossRef]
- Stalmans, W.; Laloux, M. Glucocorticoids and hepatic glycogen metabolism. In Glucocorticoid hormone Action; Baxter, J.D., Rousseau, G.G., Eds.; Springer-Verlag: New York, NY, USA, 1979; pp. 518–533. [Google Scholar]
- Kuchel, O. Stress and catecholamines. In Stress Revisited. I. Neuroendocrinology of Stress; Jasmin, G., Cantin, M., Eds.; Karger: New York, NY, USA, 1991; pp. 80–103. [Google Scholar]
- Ferguson, D.M.; Warner, R.D. Have we underestimated the impact of pre-slaughter stress on meat quality in ruminants? Meat Sci. 2008, 80, 12–19. [Google Scholar]
- Rooyackers, O.E.; Nair, K.S. Hormonal regulation of human muscle protein metabolism. Ann. Rev. Nutr. 1997, 17, 457–485. [Google Scholar] [CrossRef]
- Jacob, R.H.; Pethick, D.W.; Chapman, H.M. Muscle glycogen concentrations in commercial consignments of Australian lamb measured on farm and post-slaughter after three different lairage periods. Austr. J. Exper. Agr. 2005, 45, 543–552. [Google Scholar] [CrossRef]
- Pemberton, P.A.; Stein, P.E.; Pepys, M.B.; Potter, J.M.; Carrell, R.W. Hormone binding globulins undergo serpin conformational change in inflammation. Nature 1988, 336, 257–258. [Google Scholar] [CrossRef]
- Rescher, U.; Goebeler, V.; Wilbers, A.; Gerke, V. Proteolytic cleavage of annexin 1 by human leukocyte elastase. Biochim. Biophys. Acta 2006, 1763, 1320–1324. [Google Scholar] [CrossRef]
- Kaliner, M. Mechanisms of glucocorticosteroid action in bronchial asthma. J. Allerg. Clin. Immunol. 1985, 76, 321–329. [Google Scholar] [CrossRef]
- Reid, S.D.; Perry, S.F. The effects and physiological consequences of raised levels of cortisol on rainbow trout (Oncorhynchus mykiss) erythrocyte beta-adrenoreceptors. J. Exper. Biol. 1991, 158, 217–240. [Google Scholar]
- Kita, H.; Jorgensen, R.K.; Reed, C.E.; Dunnette, S.L.; Swanson, M.C.; Bartemes, K.R.; Squillace, D.; Blomgren, J.; Bachman, K.; Gleich, G.L. Mechanism of topical glucocorticoid treatment of hay fever: IL-5 and eosinophil activation during natural allergen exposure are suppressed, but IL-4, IL-6, and IgE antibody production are unaffected. J. Allerg. Clin. Immunol. 2000, 106, 521–529. [Google Scholar] [CrossRef]
- Angeli, A.; Masera, R.G.; Sartori, M.L.; Fortunati, N.; Racca, S.; Dovio, A.; Staurenghi, A.; Frairia, R. Modulation by cytokines of glucocorticoid action. Ann. N. Y. Acad. Sci. 1999, 876, 210–220. [Google Scholar] [CrossRef]
- Uhlar, C.M.; Whitehead, A.S. Serum amyloid A, the major vertebrate acute-phase reactant. Eur. J. Biochem. 1999, 265, 501–523. [Google Scholar] [CrossRef]
- Moisan, M. Genotype-phenotype associations in understanding the role of corticosteroid-binding globulin in health and disease animal models. Mol. Cell. Endocrinol. 2010, 316, 35–41. [Google Scholar] [CrossRef]
- MacFarlane, M.S.; Breen, K.M.; Sakurai, H.; Adams, B.M.; Adams, T.E. Effect of duration of infusion of stress-like concentrations of cortisol on follicular development and the preovulatory surge of LH in sheep. Anim. Reprod. Sci. 2000, 63, 167–175. [Google Scholar] [CrossRef]
- Breen, K.M.; Billings, H.J.; Wagenmaker, E.R.; Wessinger, E.W.; Karsch, F.J. Endocrine basis for disruptive effects of cortisol on preovulatory events. Endocrinology 2005, 146, 2107–2115. [Google Scholar] [CrossRef]
- Whittle, W.L.; Patel, F.A.; Alfaidy, N.; Holloway, A.C.; Fraser, M.; Gyomorey, S.; Lye, S.J.; Gibb, W.; Challis, J.R.G. Glucocorticoid regulation of human and ovine parturition: The relationship between foetal hypothalamic-pituitary-adrenal axis activation and intrauterine prostaglandin production. Biol. Reprod. 2001, 64, 1019–1034. [Google Scholar]
- Hough, D.; Swart, P.; Cloete, S.W.P. Comparison of peripartum steroid profiles and the relation to lamb survival in South African Merinos divergently selected for multiple rearing ability. Unpublished data. 2013. [Google Scholar]
- Matthews, S.G. Dynamic changes in glucocorticoid and mineralocorticoid receptor mRNA in the developing guinea pig brain. Develop. Brain Res. 1998, 107, 123–132. [Google Scholar] [CrossRef]
- Meyer, U.; Kruhoffer, M.; Flugge, G.; Fuchs, E. Cloning of glucocorticoid receptor and mineralocorticoid receptor cDNA and gene expression in the central nervous system of the tree shrew (Tupaia belangeri). Mol. Brain Res. 1998, 55, 243–253. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hough, D.; Swart, P.; Cloete, S. Exploration of the Hypothalamic-Pituitary-Adrenal Axis to Improve Animal Welfare by Means of Genetic Selection: Lessons from the South African Merino. Animals 2013, 3, 442-474. https://doi.org/10.3390/ani3020442
Hough D, Swart P, Cloete S. Exploration of the Hypothalamic-Pituitary-Adrenal Axis to Improve Animal Welfare by Means of Genetic Selection: Lessons from the South African Merino. Animals. 2013; 3(2):442-474. https://doi.org/10.3390/ani3020442
Chicago/Turabian StyleHough, Denise, Pieter Swart, and Schalk Cloete. 2013. "Exploration of the Hypothalamic-Pituitary-Adrenal Axis to Improve Animal Welfare by Means of Genetic Selection: Lessons from the South African Merino" Animals 3, no. 2: 442-474. https://doi.org/10.3390/ani3020442
APA StyleHough, D., Swart, P., & Cloete, S. (2013). Exploration of the Hypothalamic-Pituitary-Adrenal Axis to Improve Animal Welfare by Means of Genetic Selection: Lessons from the South African Merino. Animals, 3(2), 442-474. https://doi.org/10.3390/ani3020442