
Sexually Dimorphic Effect of Environmental Enrichment and Weaning on Stress in Piglets

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Management
2.2. Environmental Enrichment
- (1)
- Massages: one person picked up each piglet individually and provided them with tactile stimulation on their back and sides through one multi-purpose plastic glove and one domestic use plastic brush, both heavy-duty items.
- (2)
- Balls: ten 10 cm-diameter soft plastic balls with no filling were provided and placed on the floor of the pen.
- (3)
- Plastic hose: three 15 cm-pieces of ½ inch heavy-duty plastic hose tied together with a 40 cm cotton rope fixed to the pen walls.
- (4)
- Rope: three 20 cm-pieces of 9 mm polypropylene/polyester rope tied together through a 40 cm cotton rope and fixed to the pen walls.
- (5)
- Wood: 20 × 5 cm pine wood pieces, put together in a star shape with a 40 cm cotton rope.
2.3. Cortisol Fecal Metabolites
2.4. Behavior Analysis
2.5. Statistical Analysis
3. Results
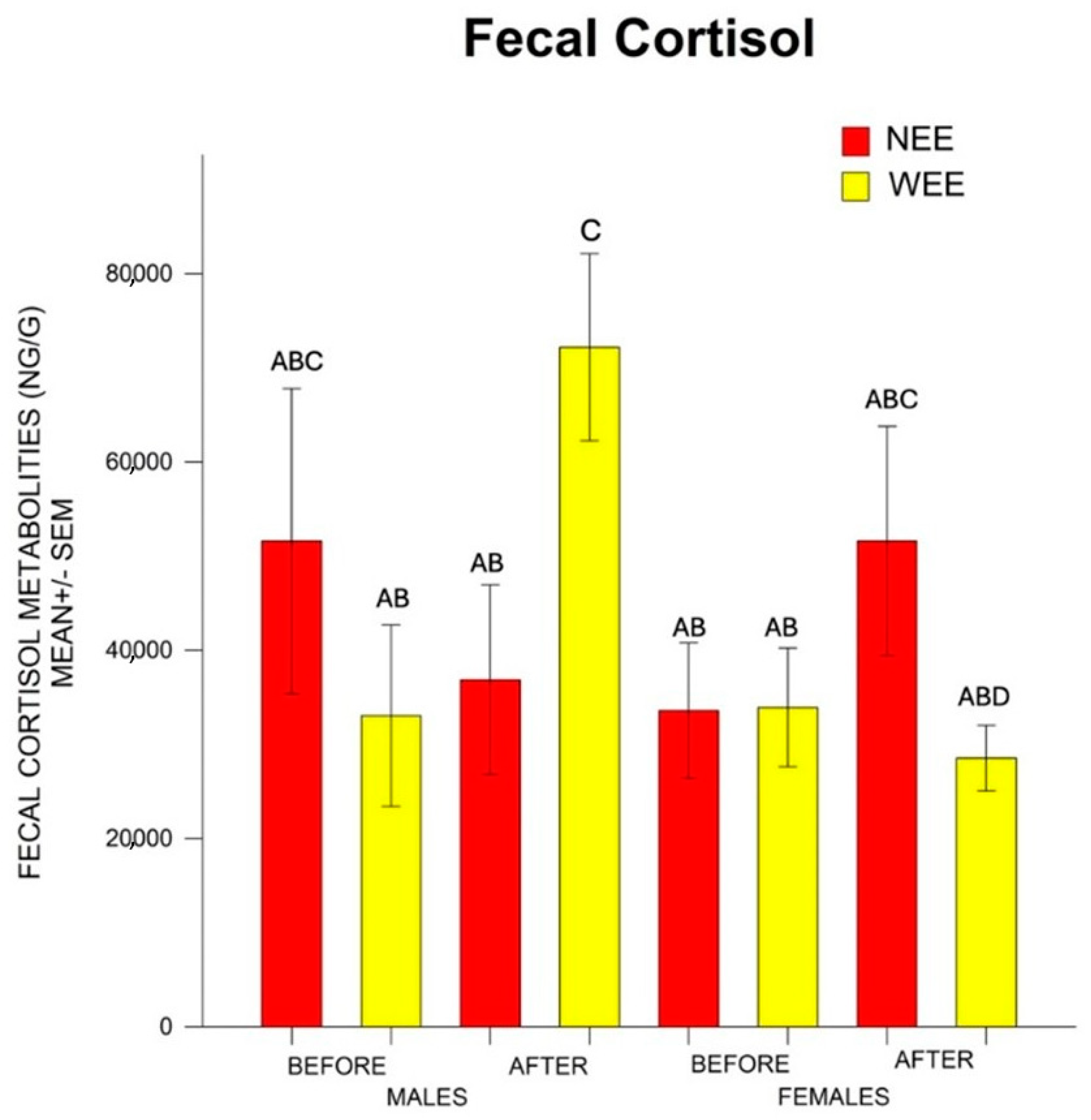
3.1. Fecal Cortisol Metabolites
3.2. Behavior
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bolhuis, J.E.; Schouten, W.G.P.; Schrama, J.W.; Wiegant, V.M. Behavioural development of pigs with different coping characteristics in barren and substrate-enriched housing conditions. Appl. Anim. Behav. Sci. 2005, 93, 213–228. [Google Scholar] [CrossRef]
- Marchant-Forde, J.N. Introduction to the welfare of pigs. In The Welfare of Pigs; Marchant-Forde, J.N., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 1–12. [Google Scholar]
- Pedersen, L.J. Chapter 1-Overview of commercial pig production systems and their main welfare challenges. In Advances in Pig Welfare, 2nd ed.; Camerlink, I., Baxter, E.M., Eds.; Woodhead Publishing: Cambridge, UK, 2018; pp. 3–25. [Google Scholar]
- Beattie, V.E.; O’Connell, N.E.; Kilpatrick, D.J.; Moss, B.W. Influence of environmental enrichment on welfare-related behavioural and physiological parameters in growing pigs. Anim. Sci. 2000, 70, 443–450. [Google Scholar] [CrossRef]
- Schouten, W.; Rushen, J.; De Passillé, A.M.B. Stereotypic behavior and heart rate in pigs. Physiol. Behav. 1991, 50, 617–624. [Google Scholar] [CrossRef]
- Hörning, B. Space allowance. In On Farm Monitoring of Pig Welfare; Verlade, A., Geers, R., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2007; pp. 121–126. [Google Scholar]
- Read, E.; Baxter, E.M.; Farish, M.; D’Eath, R.B. Trough half empty: Pregnant sows are fed under half of their ad libitum intake. Anim. Welf. 2020, 29, 151–162. [Google Scholar] [CrossRef]
- Grimberg-Henrici, C.G.; Vermaak, P.; Bolhuis, J.E.; Nordquist, R.E.; van der Staay, F.J. Effects of environmental enrichment on cognitive performance of pigs in a spatial holeboard discrimination task. Anim. Cogn. 2016, 19, 271–283. [Google Scholar] [CrossRef]
- Douglas, C.; Bateson, M.; Walsh, C.; Bédué, A.; Edwards, S.A. Environmental enrichment induces optimistic cognitive biases in pigs. Appl. Anim. Behav. Sci. 2012, 139, 65–73. [Google Scholar] [CrossRef]
- Barnett, J.L. Effects of confinement and research needs to underpin welfare standards. J. Vet. Behav. 2007, 2, 213–218. [Google Scholar] [CrossRef]
- Luo, L.; Jansen, C.A.; Bolhuis, J.E.; Arts, J.A.J.; Kemp, B.; Parmentier, H.K. Early and later life environmental enrichment affect specific antibody responses and blood leukocyte subpopulations in pigs. Physiol. Behav. 2020, 217, 112799. [Google Scholar] [CrossRef]
- Curtis, A.L.; Bethea, T.; Valentino, R.J.J.N. Sexually dimorphic responses of the brain norepinephrine system to stress and corticotropin-releasing factor. Neuropsychopharmacology 2006, 31, 544–554. [Google Scholar] [CrossRef]
- Luine, V.N.; Beck, K.; Bowman, R.E.; Frankfurt, M.; Maclusky, N.J. Chronic stress and neural function: Accounting for sex and age. J. Neuroendocrinol. 2007, 19, 743–751. [Google Scholar] [CrossRef]
- Van Keulen, B.J.; Dolan, C.V.; Van Der Voorn, B.; Andrew, R.; Walker, B.R.; Pol, H.H.; Boomsma, D.; Rotteveel, J.; Finken, M.J.J. Sexual dimorphism in cortisol metabolism throughout pubertal development: A longitudinal study. Endocr. Connect. 2020, 9, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Biason-Lauber, A. Control of sex development. Best Pract. Res. Clin. Endocrinol. Metab. 2010, 24, 163–186. [Google Scholar] [CrossRef]
- Dunshea, F.R. Sexual dimorphism in growth of sucking and growing pigs. Asian-Australas. J. Anim. Sci. 2001, 14, 1610–1615. [Google Scholar] [CrossRef]
- Schmid, E. Atlas of Animal Bones. For Prehistorians, Archaeologists and Quaternary Geologists; Elsevier Publishing Company: New York, NY, USA, 1972. [Google Scholar]
- Mayer, J.J.; Brisbin, I.L. Sex identification of Sus scrofa based on canine morphology. J. Mammal. 1988, 69, 408–412. [Google Scholar] [CrossRef]
- Gray, J.A. Sex differences in emotional behaviour in mammals including man: Endocrine bases. Acta Psychol. 1971, 35, 29–46. [Google Scholar] [CrossRef] [PubMed]
- Power, G.; Pluske, J.; Campbell, R.; Cranwell, P.; Kerton, D.; King, R.; Dunshea, F. Effect of sex, weight and age on post-weaning growth of pigs. In Proceedings of the Nutrition Society of Australia, V20, Twentieth Annual Scientific Meeting, Sydney, Australia, 28 September–1 October 1996. [Google Scholar]
- Dunshea, F.R.; Boyce, J.M.; King, R.H. Effect of supplemental nutrients on the growth performance of sucking pigs. Aust. J. Agric. Res. 1998, 49, 883–888. [Google Scholar] [CrossRef]
- Conrad, M.S.; Dilger, R.N.; Johnson, R.W. Brain growth of the domestic pig (Sus scrofa) from 2 to 24 weeks of age: A longitudinal MRI study. Dev. Neurosci. 2012, 34, 291–298. [Google Scholar] [CrossRef]
- Moisan, M.-P. Sexual dimorphism in glucocorticoid stress response. Int. J. Mol. Sci. 2021, 22, 3139. [Google Scholar] [CrossRef] [PubMed]
- Newberry, R.C. Environmental enrichment: Increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 1995, 44, 229–243. [Google Scholar] [CrossRef]
- Yang, C.-H.; Ko, H.-L.; Salazar, L.C.; Llonch, L.; Manteca, X.; Camerlink, I.; Llonch, P. Pre-weaning environmental enrichment increases piglets’ object play behaviour on a large scale commercial pig farm. Appl. Anim. Behav. Sci. 2018, 202, 7–12. [Google Scholar] [CrossRef]
- Martin, J.E.; Ison, S.H.; Baxter, E.M. The influence of neonatal environment on piglet play behaviour and post-weaning social and cognitive development. Appl. Anim. Behav. Sci. 2015, 163, 69–79. [Google Scholar] [CrossRef]
- Espejo, B.G.; Paredes-Ramos, P.; Ahuja-Aguirre, C.; Carrasco-García, A.; Naranjo-Chacón, F. Effect of environmental enrichment on performance of piglets during anxiety tests. Rev. Bio Cienc. 2020, 7, e789. [Google Scholar]
- Espejo, B.G.; Ahuja-Aguirre, C.; Carrasco-García, A.A.; Hernandez-Cruz, B.; Paredes-Ramos, P. Environmental enrichment for primiparous and multiparous pregnant sows and its effect on cortisol and behavior at farrowing and production parameters at weaning. Livest. Sci. 2022, 265, 105103. [Google Scholar] [CrossRef]
- Graham, L.; Schwarzenberger, F.; Möstl, E.; Galama, W.; Savage, A. A versatile enzyme immunoassay for the determination of progestogens in feces and serum. Zoo Biol. 2001, 20, 227–236. [Google Scholar] [CrossRef]
- Luo, L.; Reimert, I.; Middelkoop, A.; Kemp, B.; Bolhuis, J.E. Effects of early and current environmental enrichment on behavior and growth in pigs. Front. Vet. Sci. 2020, 7, 268. [Google Scholar] [CrossRef] [PubMed]
- Moeser, A.J.; Roney, A.; Fardisi, M.; Thelen, K. Biological sex: An understudied factor driving disease susceptibility in pigs. J. Anim. Sci. 2022, 100, skac146. [Google Scholar] [CrossRef]
- McCarthy, M.M. A new view of sexual differentiation of mammalian brain. J. Comp. Physiol. A 2020, 206, 369–378. [Google Scholar] [CrossRef]
- McPherson, F.J.; Chenoweth, P.J. Mammalian sexual dimorphism. Anim. Reprod. Sci. 2012, 131, 109–122. [Google Scholar] [CrossRef]
- Jang, Y.D.; Ma, Y.L.; Lindemann, M.D. Intrauterine position affects fetal weight and crown-rump length throughout gestation. J. Anim. Sci. 2014, 92, 4400–4406. [Google Scholar] [CrossRef]
- Ford, J.J. Serum estrogen concentrations during postnatal development in male pigs. Exp. Biol. Med. 1983, 174, 160–164. [Google Scholar] [CrossRef]
- Baxter, E.M.; Jarvis, S.; Palarea-Albaladejo, J.; Edwards, S.A. The weaker sex? The propensity for male-biased piglet mortality. PLoS ONE 2012, 7, e30318. [Google Scholar] [CrossRef] [PubMed]
- Stenhouse, C.; Bazer, F.W.; Ashworth, C.J. Sexual dimorphism in placental development and function: Comparative physiology with an emphasis on the pig. Mol. Reprod. Dev. 2023, 90, 684–696. [Google Scholar] [CrossRef] [PubMed]
- Tallet, C.; Brilloüet, A.; Meunier-Salaün, M.C.; Paulmier, V.; Guérin, C.; Prunier, A. Effects of neonatal castration on social behaviour, human–animal relationship and feeding activity in finishing pigs reared in a conventional or an enriched housing. Appl. Anim. Behav. Sci. 2013, 145, 70–83. [Google Scholar] [CrossRef]
- Tang, A.C.; Akers, K.G.; Reeb, B.C.; Romeo, R.D.; McEwen, B.S. Programming social, cognitive, and neuroendocrine development by early exposure to novelty. Proc. Natl. Acad. Sci. USA 2006, 103, 15716–15721. [Google Scholar] [CrossRef]
- Marchant, J.; Rudd, A.; Mendl, M.T.; Broom, D.M.; Meredith, M.; Corning, S.; Simmins, P. Timing and causes of piglet mortality in alternative and conventional farrowing systems. Vet. Rec. 2000, 147, 209–214. [Google Scholar] [CrossRef]
- Otten, W.; Kanitz, E.; Tuchscherer, M. The impact of pre-natal stress on offspring development in pigs. J. Agric. Sci. 2015, 153, 907–919. [Google Scholar] [CrossRef]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef]
- Bolhuis, J.E.; Oostindjer, M.; van den Brand, H.; Gerrits, W.J.J.; Kemp, B. Voluntary feed intake in piglets: Potential impact of early experience with flavours derived from the maternal diet. In Voluntary Feed Intake in Pigs; Torrallardona, D., Roura, E., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2009; pp. 37–60. [Google Scholar]
- Morrone, B.; Bernardino, T.; Tatemoto, P.; Rodrigues, F.A.M.L.; de Moraes, J.E.; da Cruz, T.D.A.; Zanella, A.J. Indication that the presence of older conspecifics reduces agonistic behaviour in piglets at weaning. Appl. Anim. Behav. Sci. 2021, 234, 105201. [Google Scholar] [CrossRef]
- Muns, R.; Rault, J.-L.; Hemsworth, P. Positive human contact on the first day of life alters the piglet’s behavioural response to humans and husbandry practices. Physiol. Behav. 2015, 151, 162–167. [Google Scholar] [CrossRef]
- Brunton, P.J. Effects of maternal exposure to social stress during pregnancy: Consequences for mother and offspring. Reproduction 2013, 146, R175–R189. [Google Scholar] [CrossRef]
- D’Eath, R.B.; Turner, S.P. The natural behaviour of the pig. In The Welfare of Pigs; Marchant-Forde, J.N., Ed.; Springer: Dordrecht, Netherlands, 2009; pp. 13–45. [Google Scholar]
- Colson, V.; Orgeur, P.; Foury, A.; Mormède, P. Consequences of weaning piglets at 21 and 28 days on growth, behaviour and hormonal responses. Appl. Anim. Behav. Sci. 2006, 98, 70–88. [Google Scholar] [CrossRef]
- Meese, G.B.; Ewbank, R. Exploratory behaviour and leadership in the domesticated pig. Br. Vet. J. 1973, 129, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Mei, H.; Yang, B.; Luo, J.; Gan, L. The effect of mixing levels on aggression at weaning in piglets. Appl. Anim. Behav. Sci. 2016, 179, 32–38. [Google Scholar] [CrossRef]
- Bocian, M.; Jankowiak, H.; Cebulska, A.; Wiśniewska, J.; Frątczak, K.; Włodarski, W.; Kapelański, W. Differences in piglets sex proportion in litter and in body weight at birth and weaning and fattening results. J. Cent. Eur. Agric. 2012, 13, 475–482. [Google Scholar] [CrossRef]
- Cenni, C.; Fawcett, T.W. The coevolution of juvenile play–fighting and adult competition. Ethology 2018, 124, 290–301. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Behavior | Description |
|---|---|
| Laying inactive | Laying on their side or on their belly, with open or closed eyes, performing no other behavior. |
| Social exploration | Any physical and/or exploratory contact such as sniffing any part of the body of another piglet. |
| Pen exploration | Exploration and sniffing directed towards the pen walls, floor, feeder or drinker. |
| Social manipulation | Nibbling, sucking, chewing or biting at the ears, tail or another part of the body of another piglet. |
| Aggression | Thumping or pushing with the head or snout towards the body of another piglet. |
| Comfort behavior | Rubbing their body against objects or pen mates, scratching their own body with the leg or stretching the body quivering. |
| Ingestion | Eating and/or drinking water. |
| Locomotion | Continuous walk for at least 2 m. |
| Behaviour | Male Before Weaning NEE | Male Before Weaning WEE | Male After Weaning NEE | Male After Weaning WEE | Female Before Weaning NEE | Female Before Weaning WEE | Female After Weaning NEE | Female After Weaning WEE | p | |
|---|---|---|---|---|---|---|---|---|---|---|
| Laying inactive | L | 5.70 ±4.7 | 30.70 ± 5.8 | 310 ± 117.7 | 161.33 ± 62.9 | 42.50 ± 40.8 | 119.66 ± 49.1 | 313.90 ± 140.5 | 149.46 ± 53.2 | 0.73 |
| F | 4.70 a± 0.73 | 2.80 b ± 0.52 | 3.20 b ± 0.4 | 4.06 a ± 0.32 | 4.70 a ± 0.6 | 4.16 a ± 0.4 | 4.80 a ± 0.4 | 4.20 a ± 0.3 | 0.046 | |
| D | 1151.40 ± 140.1 | 1260.60 ± 238.2 | 722.20 ± 109.9 | 875.66 ± 73.8 | 1233.70 ± 122.5 | 995.76 ± 70.2 | 723.90 ± 150.5 | 954.40 ± 77.2 | 0.161 | |
| Social exploration | L | 1180.20 ± 213.5 | 1105.33 ± 208.8 | 1697.80 ± 69.1 | 645.63 ± 118.3 | 1546.70 ± 168.8 | 1036.86 ± 147.6 | 1263.20 ± 231.9 | 574.43 ± 129.6 | 0.11 |
| F | 0.90 ± 0.4 | 2.10 ± 0.39 | 0.20 ± 0.1 | 4.70 ± 1.11 | 0.20 ± 0.1 | 2.60 ± 0.5 | 0.80 ± 0.3 | 4.40 ± 0.6 | 0.42 | |
| D | 29.80 ± 12.06 | 131.86 ± 24.9 | 14.60 ± 12.3 | 303.70 ± 42.3 | 7.20 ± 6.07 | 147.90 ± 34.6 | 51.70 ± 28.5 | 251.00 ± 39.8 | 0.32 | |
| Pen exploration | L | 1466.70 ± 120.7 | 1162.80 ± 219.7 | 300.90 ±100.4 | 965.43 ± 146.2 | 1257.00 ± 217.9 | 818.53 ± 147.5 | 851.20 ± 266.1 | 662.33 ± 149.4 | 0.18 |
| F | 0.80 ± 0.32 | 0.80 ± 0.15 | 5.30 ± 0.6 | 1.90 ± 0.4 | 0.70 ± 0.3 | 2.03 ± 0.39 | 3.30 ± 0.9 | 2.76 ± 0.5 | 0.32 | |
| D | 60.50 ± 25.2 | 33.10 ± 6-2 | 432.60 ± 71.0 | 48.46 ± 12.2 | 54.70 ± 25.7 | 107.50 ± 30.4 | 251.10 ± 69.3 | 91.90 ± 18.1 | 0.11 | |
| Social manipulation | L | 921.00 ± 227.0 | 1468.50 ± 277.5 | 797.80 ± 221.1 | 1596.90 ± 94.7 | 485.40 ± 157.6 | 1210.70 ± 142.5 | 1146.40 ± 227.7 | 1655.50 ± 80.03 | 0.30 |
| F | 2.00 ± 0.6 | 0.40 ± 0.07 | 2.70 ± 0.5 | 0.36 ± 0.1 | 3.10 ± 0.7 | 1.26 ± 0.37 | 2.20 ± 0.7 | 0.26 ± 0.1 | 0.57 | |
| D | 169.50 ± 78.5 | 28.23 ± 5.3 | 261.20 ± 76.8 | 14.00 ± 8.06 | 165.50 ± 69.05 | 80.66 ± 30.4 | 154.80 ± 59.09 | 11.83 ± 7.1 | 0.70 | |
| Aggression | L | 1800.00 ± 0 | 1579.70 ± 298.5 | 1297.30 ± 209.6 | 1355.50 ± 120.7 | 1293.30 ± 209.05 | 1087.00 ± 146.9 | 1108.80 ± 198.6 | 1103.60 ± 141.3 | 0.87 |
| F | 0.55 ± 0 | 0.20 ± 0.03 | 0.80 ± 0.4 | 0.93 ± 0.2 | 0.50 ± 0.2 | 1.00 ± 0.2 | 1.20 ± 0.3 | 0.86 ± 0.2 | 0.36 | |
| D | 1.10 b ± 0 | 0.70 b ± 0.13 | 2.20 a ± 1.2 | 2.50 a ± 1.3 | 0.80 b ± 0.3 | 3.80 a ± 1.2 | 7.30 c ± 2.8 | 2.50 a ± 0.6 | 0.05 | |
| Comfort behavior | L | 1702.00 ± 98 | 1800.00 ± 340.1 | 1800.00 ± 0 | 1733.30 ± 48.9 | 1800.00 ± 0 | 1697.20 ± 65.4 | 1800.00 ± 0 | 1617.40 ± 91.5 | 0.69 |
| F | 0.10 ± 0.1 | 0.06 ± 0 | 0.01 ± 0 | 0.10 ± 0.07 | 0.06 ± 0 | 0.10 ± 0.05 | 0.06 ± 0 | 0.13 ± 0.06 | 0.41 | |
| D | 2.40 ± 2.4 | 0.01 ± 0 | 0.007 ± 0 | 5.80 ± 5.0 | 3.33 ± 0 | 1.50 ± 0.8 | 0.07 ± 0 | 7.70 ± 4.6 | 0.86 | |
| Ingestion | L | 1075.20 ± 0.5 | 1066.20 ± 201.4 | 809.80 ± 224.6 | 617.80 ± 122.6 | 938.50 ± 207.9 | 778.03 ± 131.4 | 782.50 ± 213.01 | 454.96 ± 107.8 | 0.97 |
| F | 2.10 a ± 80.7 | 1.90 b ± 0.3 | 1.80 ab ± 0.5 | 3.40 a ± 0.4 | 1.60 b ± 0.4 | 2.90 a ± 0.4 | 3.10 a ± 0.6 | 3.46 a ± 0.4 | 0.05 | |
| D | 315.90 ab ± 0 | 221.16 a ± 41.7 | 180.50 d ± 56.7 | 395.40 b ± 47.9 | 273.10 a ± 85.4 | 289.83 a ± 44.6 | 415.00 c ± 104.7 | 371.46 b ± 42.1 | 0.043 | |
| Locomotion | L | 853.20 ± 220.4 | 858.76 ± 162.2 | 735.50 ± 86.5 | 416.06 ± 81.8 | 815.40 ± 219.2 | 672.30 ± 116.5 | 447.10 ± 183.2 | 475.86 ± 115.7 | 0.75 |
| F | 2.70 ± 0.7 | 2.43 ± 0.45 | 4.70 ± 0.6 | 5.06 ± 0.4 | 2.50 ± 0.6 | 3.26 ± 0.37 | 6.00 ± 0.8 | 4.50 ± 0.5 | 0.10 | |
| D | 77.50 ± 32.0 | 124.00 ± 23.4 | 168.70 ± 28.7 | 159.23 ± 26.0 | 72.00 ± 32.6 | 166.80 ± 33.4 | 139.40 ± 32.4 | 107.40 ± 16.5 | 0.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espejo-Beristain, G.; Paredes-Ramos, P.; Ahuja-Aguirre, C.; Carrasco-García, A. Sexually Dimorphic Effect of Environmental Enrichment and Weaning on Stress in Piglets. Animals 2025, 15, 1099. https://doi.org/10.3390/ani15081099
Espejo-Beristain G, Paredes-Ramos P, Ahuja-Aguirre C, Carrasco-García A. Sexually Dimorphic Effect of Environmental Enrichment and Weaning on Stress in Piglets. Animals. 2025; 15(8):1099. https://doi.org/10.3390/ani15081099
Chicago/Turabian StyleEspejo-Beristain, Guadalupe, Pedro Paredes-Ramos, Concepción Ahuja-Aguirre, and Apolo Carrasco-García. 2025. "Sexually Dimorphic Effect of Environmental Enrichment and Weaning on Stress in Piglets" Animals 15, no. 8: 1099. https://doi.org/10.3390/ani15081099
APA StyleEspejo-Beristain, G., Paredes-Ramos, P., Ahuja-Aguirre, C., & Carrasco-García, A. (2025). Sexually Dimorphic Effect of Environmental Enrichment and Weaning on Stress in Piglets. Animals, 15(8), 1099. https://doi.org/10.3390/ani15081099