
Analysis of the Microbial Community Structure of Ixodes persulcatus at Each Developmental Stage

 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA and RNA Extraction
2.3. Library Construction and High-Throughput Sequencing
2.4. Data Analysis
3. Result
3.1. Sequencing Results of Illumina PE250
3.1.1. General Statistics
3.1.2. Alpha Diversity Analysis
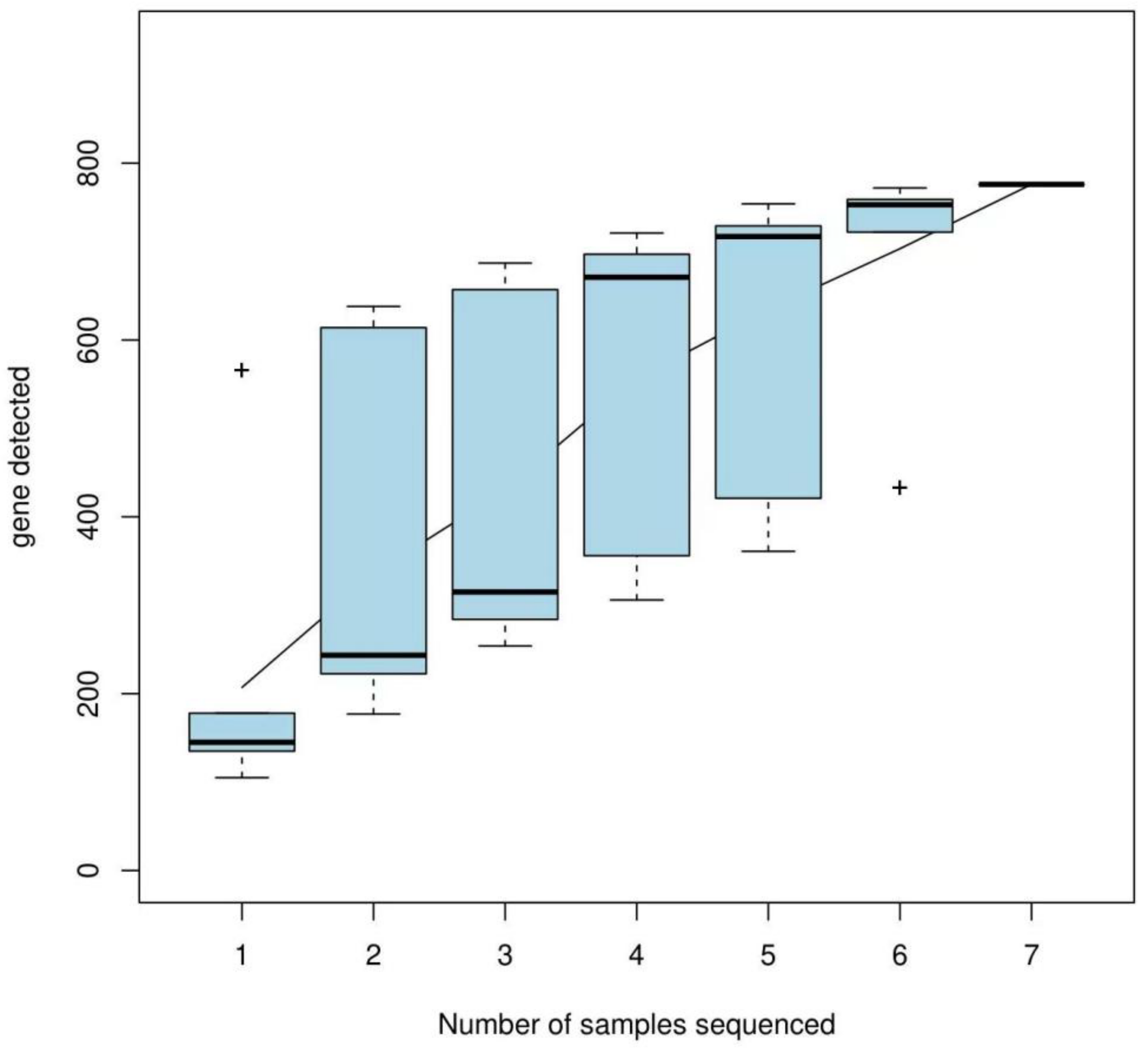
3.1.3. OTU Cluster Analysis
3.1.4. Microbial Population
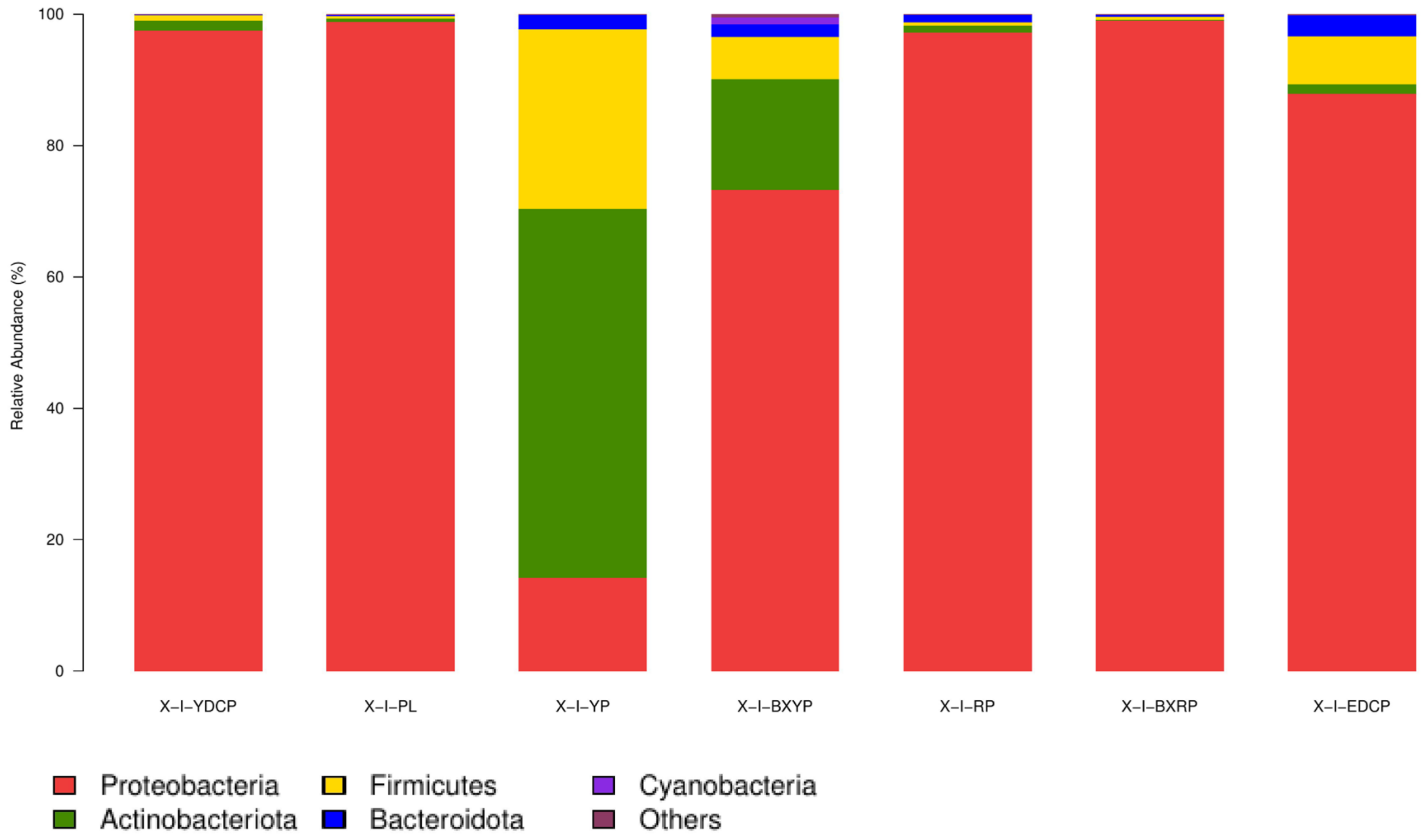
Microbial Community Composition at the Phylum Level
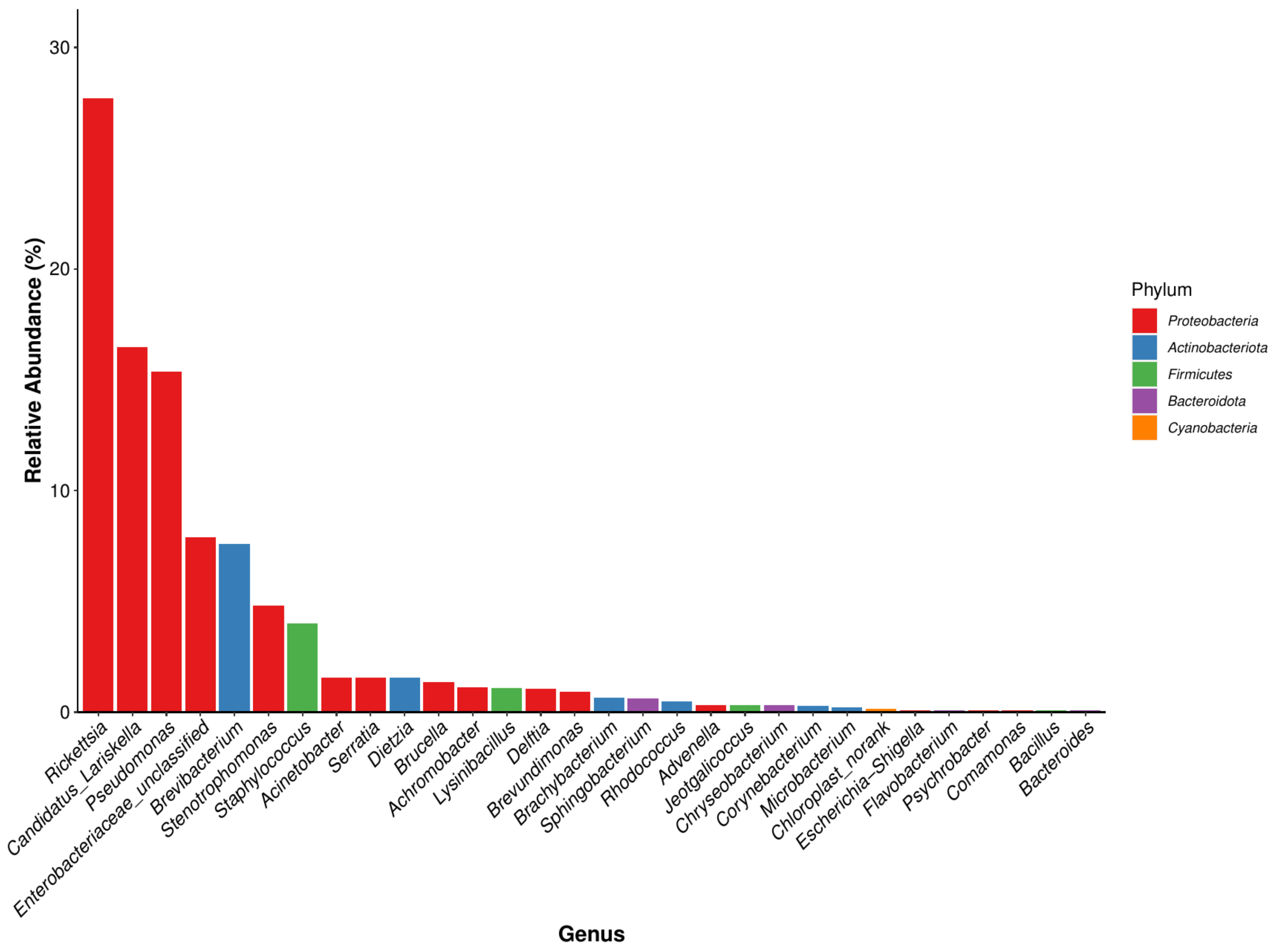
Microbial Community Composition at the Genus Level
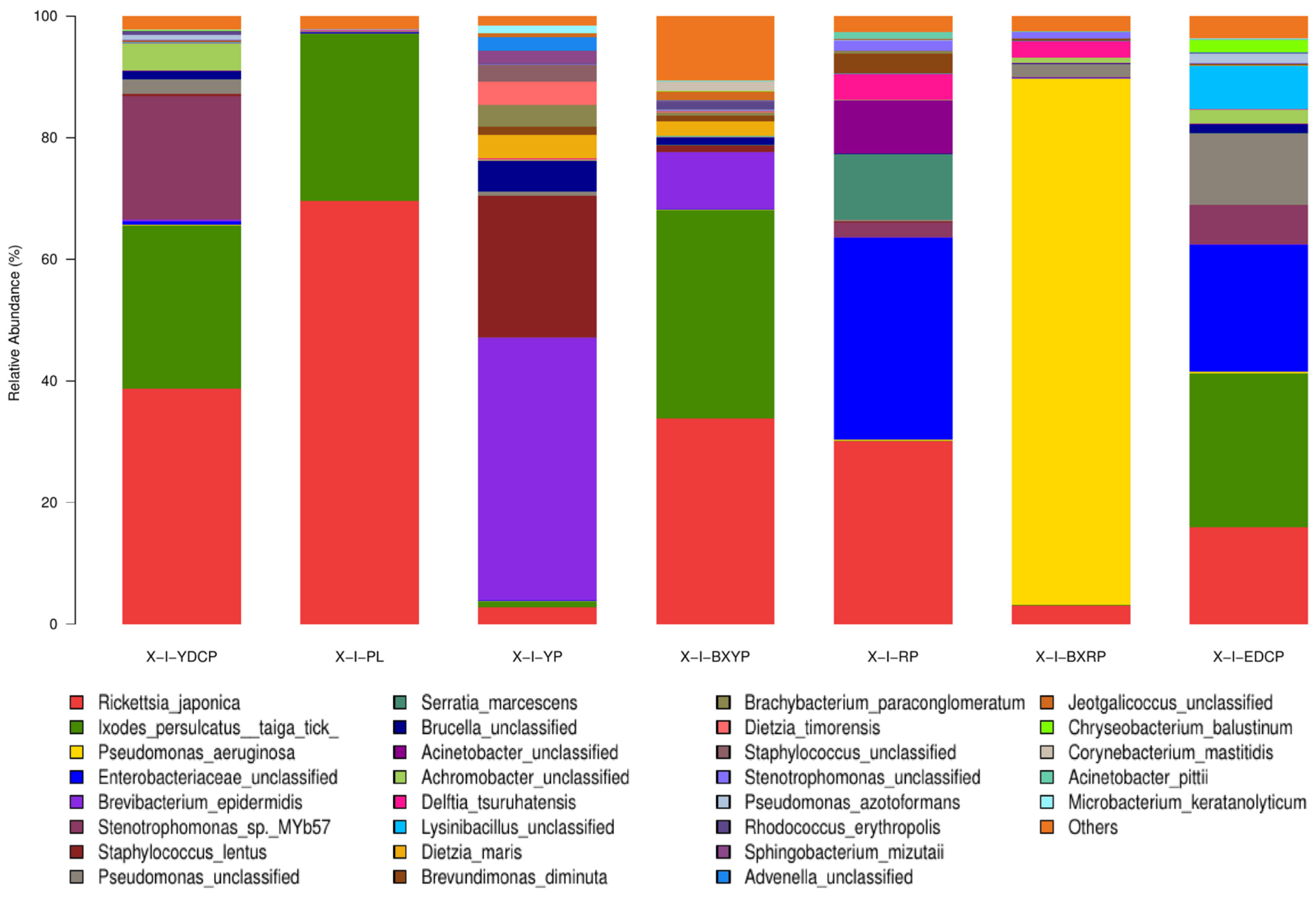
Microbial Community Composition at the Species Level
3.2. Sequencing Results of Illumina PE150
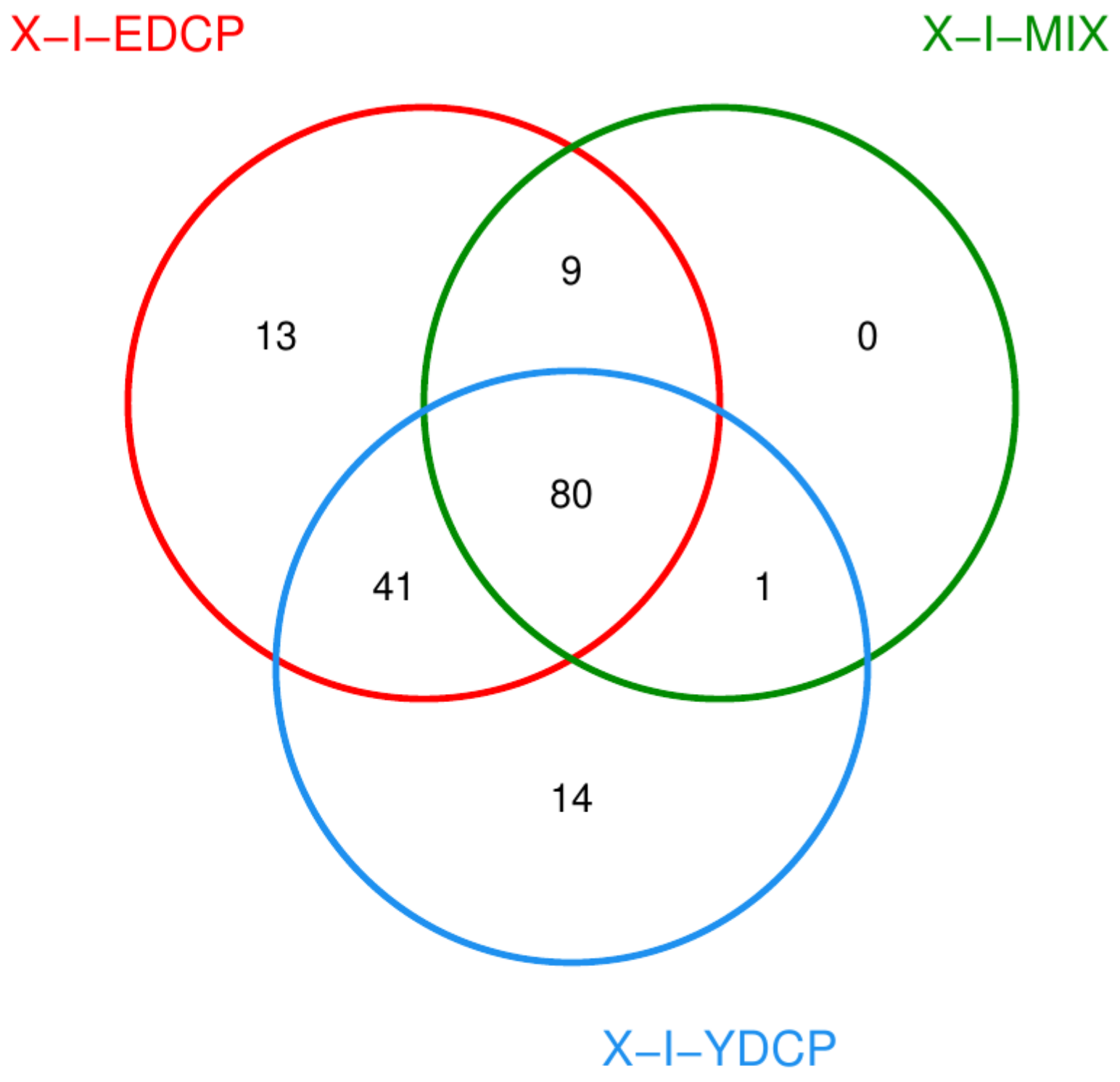
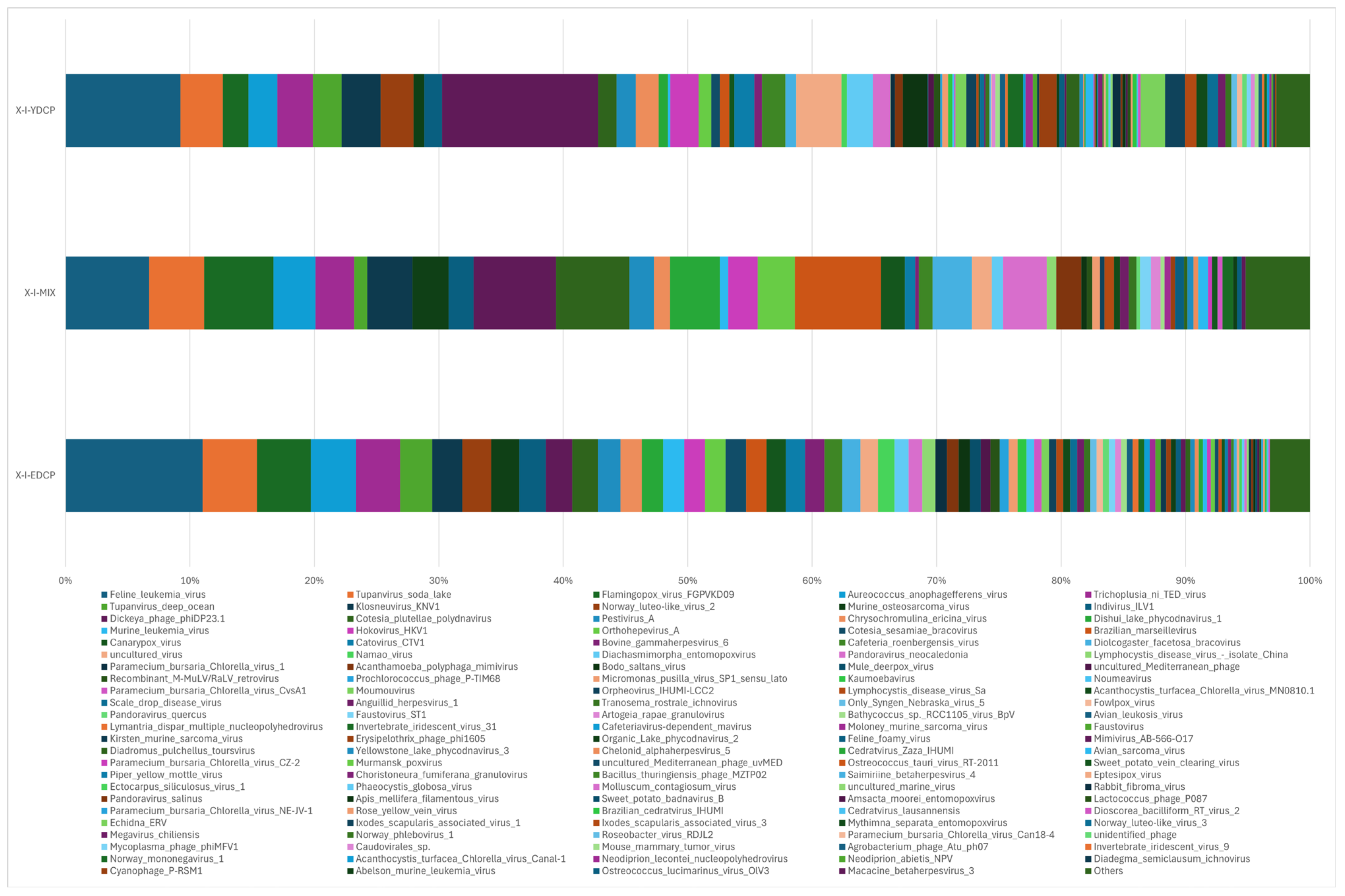
3.2.1. Species Classification Annotation Data
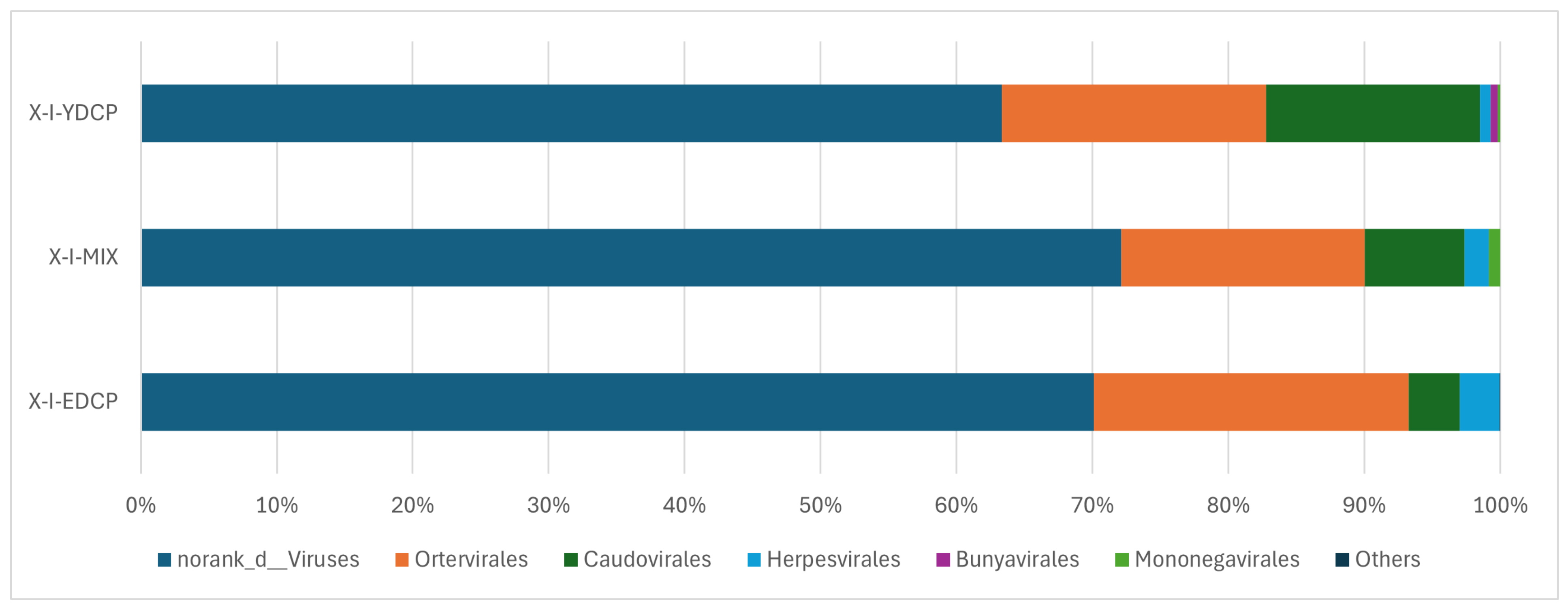
3.2.2. Species Composition at the Order Level
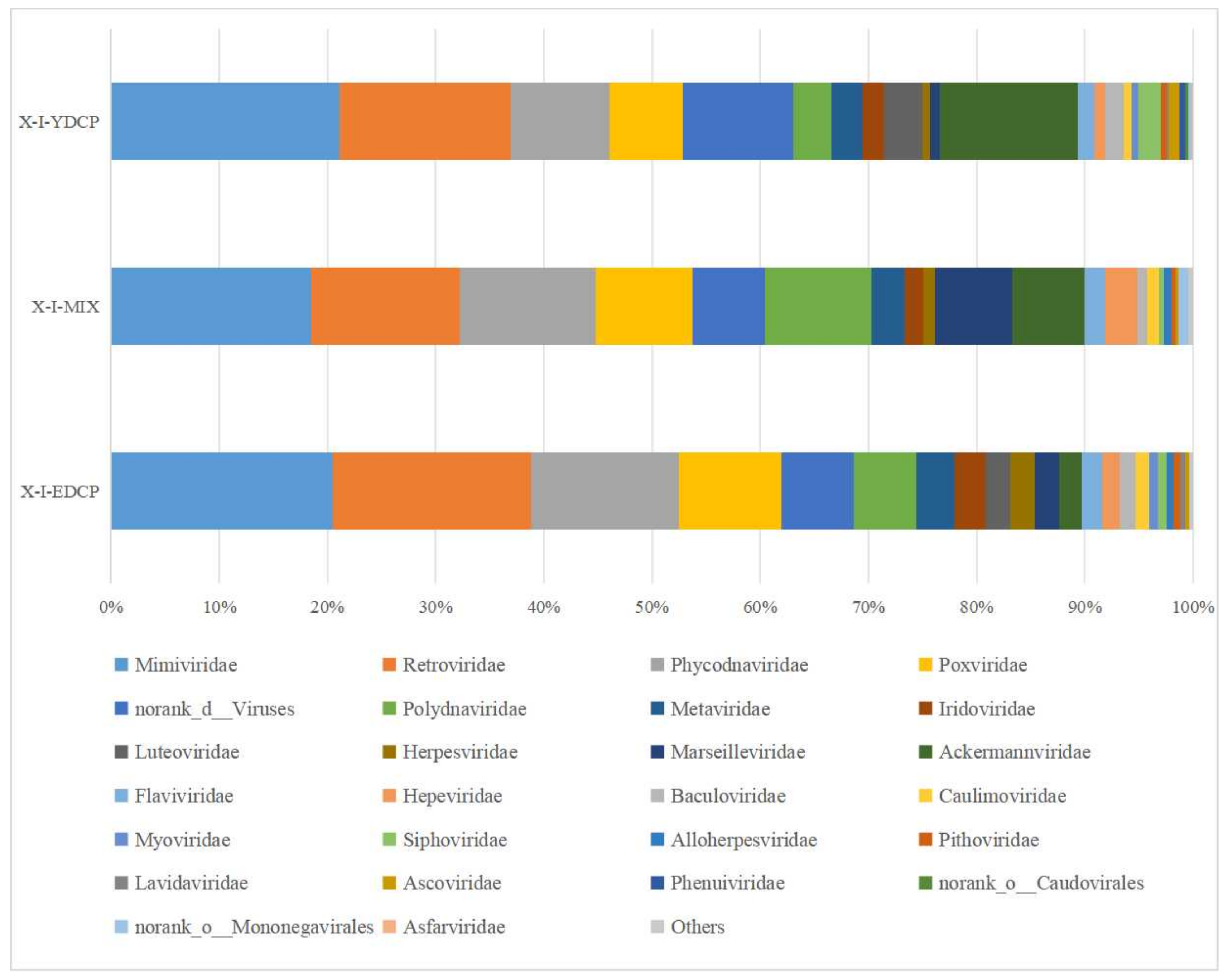
3.2.3. Species Composition at the Family Level
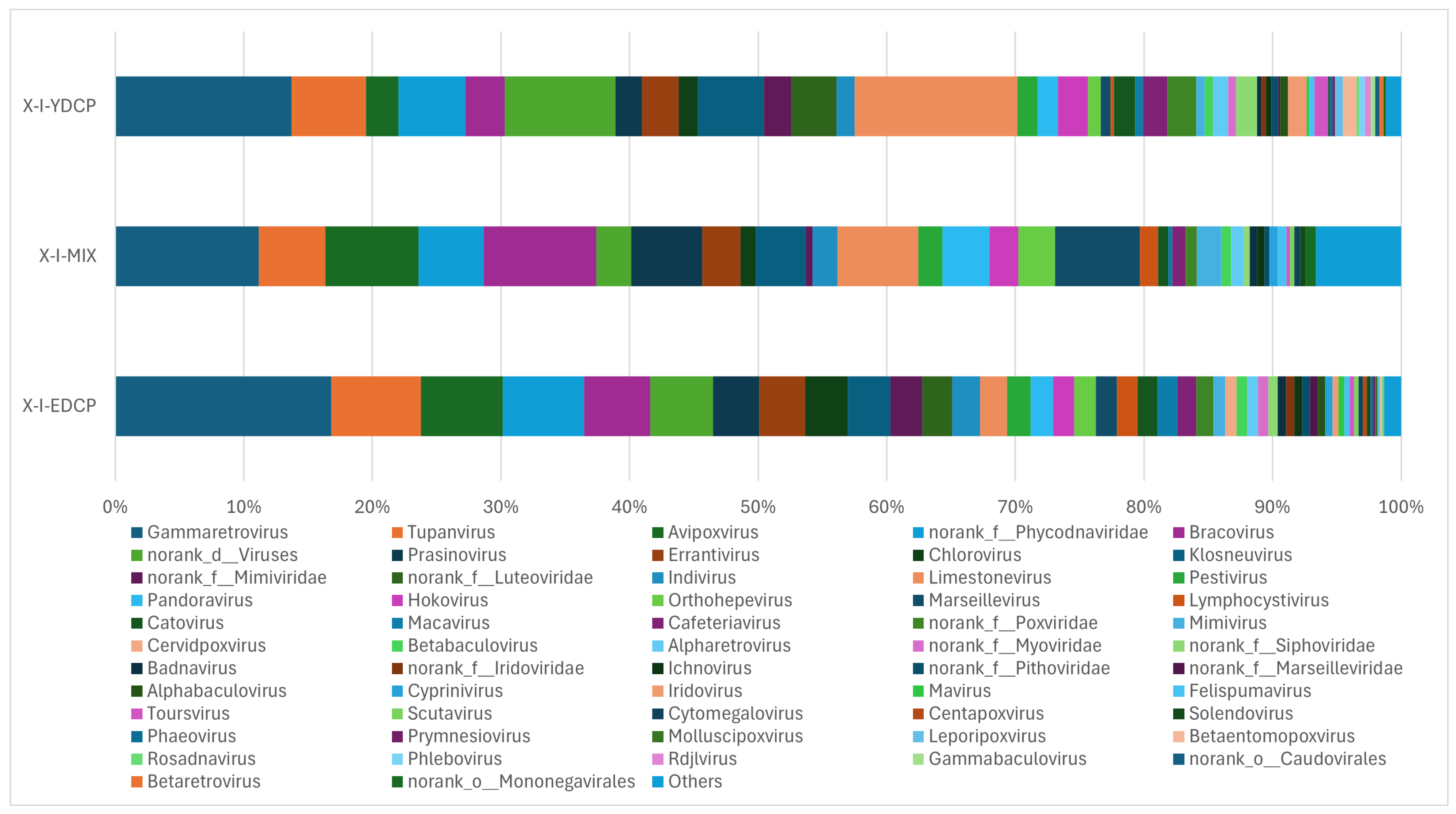
3.2.4. Species Composition at the Genus Level
3.2.5. Species Composition at the Species Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jia, N.; Wang, J.; Shi, W.; Du, L.; Sun, Y.; Zhan, W.; Jiang, J.F.; Wang, Q.; Zhang, B.; Ji, P.; et al. Large-scale comparative analyses of tick genomes elucidate their genetic diversity and vector capacities. Cell 2020, 182, 1328–1340. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.S.; Ogden, N.H. Ticks, human babesiosis and climate change. Pathogens 2021, 10, 1430. [Google Scholar] [CrossRef] [PubMed]
- Grigoryeva, L.A.; Stanyukovich, M.K. Life cycle of the taiga tick Ixodes persulcatus (Acari: Ixodidae) in the North-West of Russia. Exp. Appl. Acarol. 2016, 69, 347–357. [Google Scholar] [CrossRef]
- Wang, S.S.; Liu, J.Y.; Wang, B.Y.; Wang, W.J.; Cui, X.M.; Jiang, J.F.; Sun, Y.; Guo, W.B.; Pan, Y.-S.; Zhou, Y.H.; et al. Geographical distribution of Ixodes persulcatus and associated pathogens: Analysis of integrated data from a China field survey and global published data. One Health 2023, 16, 100508. [Google Scholar] [CrossRef]
- Sousa-Paula, L.C.; Schwan, T.G.; Tirloni, L. Ornithodoros hermsi. Trends Parasitol. 2024, 40, 765–766. [Google Scholar] [CrossRef]
- Fukunaga, M.; Takahashi, Y.; Tsuruta, Y.; Matsushita, O.; Ralph, D.; McClelland, M.; Nakao, M. Genetic and phenotypic analysis of Borrelia miyamotoi sp. nov., isolated from the ixodid tick Ixodes persulcatus, the vector for Lyme disease in Japan. Int. J. Syst. Bacteriol. 1995, 45, 804–810. [Google Scholar] [CrossRef]
- Kholodilov, I.S.; Belova, O.A.; Ivannikova, A.Y.; Gadzhikurbanov, M.N.; Makenov, M.T.; Yakovlev, A.S.; Polienko, A.E.; Dereventsova, A.V.; Litov, A.G.; Gmyl, L.V.; et al. Distribution and characterisation of tick-borne flavi-, flavi-like, and phenuiviruses in the Chelyabinsk region of Russia. Viruses 2022, 14, 2699. [Google Scholar] [CrossRef]
- Livanova, N.N.; Fomenko, N.V.; Akimov, I.A.; Ivanov, M.J.; Tikunova, N.V.; Armstrong, R.; Konyaev, S.V. Dog survey in Russian veterinary hospitals: Tick identification and molecular detection of tick-borne pathogens. Parasites Vectors 2018, 11, 591. [Google Scholar] [CrossRef]
- Černý, J.; Buyannemekh, B.; Needham, T.; Gankhuyag, G.; Oyuntsetseg, D. Hard ticks and tick-borne pathogens in Mongolia—A review. Ticks Tick-Borne Dis. 2019, 10, 101268. [Google Scholar] [CrossRef]
- Pakanen, V.M.; Sormunen, J.J.; Sippola, E.; Blomqvist, D.; Kallio, E.R. Questing abundance of adult taiga ticks Ixodes persulcatus and their Borrelia prevalence at the north-western part of their distribution. Parasites Vectors 2020, 13, 384. [Google Scholar] [CrossRef]
- Jaenson, T.G.T.; Wilhelmsson, P. First records of tick-borne pathogens in populations of the taiga tick Ixodes persulcatus in Sweden. Parasites Vectors 2019, 12, 559. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Y.; Li, L.; Xu, W.B.; Yuan, Y.X.; Liang, X.J.; Zhang, L.; Wei, Z.K.; Sui, L.Y.; Zhao, Y.H.; Cui, Y.Y.; et al. Extensive diversity of RNA viruses in ticks revealed by metagenomics in northeastern China. PloS Neglected Trop. Dis. 2022, 16, e0011017. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.K.; Zhang, L.; Hou, X.X.; Bao, Z.H.; Zeng, Y.; He, L.J.; Liu, Z.L.; Zhou, H.J.; Hao, Q.; Dong, A.Y. Surveillance of tick-borne bacteria infection in ticks and forestry populations in Inner Mongolia, China. Front. Public Health 2024, 12, 1302133. [Google Scholar] [CrossRef]
- Wang, Z.D.; Wang, B.; Wei, F.; Han, S.Z.; Zhang, L.; Yang, Z.T.; Yan, Y.; Lv, X.L.; Li, L.; Wang, S.C.; et al. A new segmented virus associated with human febrile illness in China. N. Engl. J. Med. 2019, 380, 2116–2125. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.L.; Liu, Z.Y.; Li, L.; Xu, W.B.; Yuan, Y.X.; Liang, X.J.; Zhang, L.; Wei, Z.K.; Sui, L.Y.; Zhao, Y.H.; et al. Yezo virus infection in tick-bitten patient and ticks, northeastern China. Emerg. Infect. Dis. 2023, 29, 797–800. [Google Scholar] [CrossRef]
- Li, X.H.; Ji, H.W.; Wang, D.; Che, L.H.; Zhang, L.; Li, L.; Yin, Q.; Liu, Q.; Wei, F.; Wang, Z.D. Molecular detection and phylogenetic analysis of tick-borne encephalitis virus in ticks in northeastern China. J. Med. Virol. 2022, 94, 507–513. [Google Scholar] [CrossRef]
- Dong, R.X.; Fang, R.Y.; Yang, X.; Sun, Y.F.; Zhang, Y.S.; Li, S. An updated review on the spatial distribution of Borrelia burgdorferi Sensu Lato Across ticks, animals and humans in northeastern China and adjacent Regions. Zoonoses Public Health 2024, 71, 763–778. [Google Scholar] [CrossRef]
- Hu, Y.C.; Yin, T.; Ma, W.; Qiu, J.W.; Zhang, J.X.; Wang, Q. A systematic review and meta-analysis of the prevalence of tick-borne SFGR in China from 2000 to 2022. PLoS Neglected Trop. Dis. 2024, 18, e0012550. [Google Scholar] [CrossRef]
- Zhao, G.P.; Wang, Y.X.; Fan, Z.W.; Ji, Y.; Liu, M.J.; Zhang, W.H.; Li, X.L.; Zhou, S.X.; Li, H.; Liang, S.; et al. Mapping ticks and tick-borne pathogens in China. Nat. Commun. 2021, 12, 1075. [Google Scholar] [CrossRef]
- Zhao, L.; Ma, Y.M.; Yang, B.; Han, W.X.; Zhao, W.H.; Chai, H.L.; Zhang, Z.S.; Zhan, Y.J.; Wang, L.F.; Xing, Y.; et al. Comparative analysis of microbial communities in different growth stages of Dermacentor nuttalli. Front. Vet. Sci. 2022, 9, 1021426. [Google Scholar] [CrossRef]
- Duan, D.Y.; Liu, G.H.; Cheng, T.Y.; Wang, Y.Q. Microbial population analysis of the midgut of Melophagus ovinus via high-throughput sequencing. Parasite Vectors 2017, 10, 382. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Lv, X.L.; Zhang, X.; Han, S.Z.; Wang, Z.D.; Li, L.; Sun, H.T.; Ma, L.X.; Cheng, Z.L.; Shao, J.W.; et al. Identification of a new orthonairovirus associated with human febrile illness in China. Nat. Med. 2021, 27, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Sirotkin, M.B.; Korenberg, E.I. Thermal constants of development of Ixodes persulcatus and Ixodes ricinus ticks, which determine the duration of their life cycle and their distribution. Entmol. Rev. 2022, 102, 257–263. [Google Scholar] [CrossRef]
- Duan, D.Y.; Cheng, T.Y. Determination of the microbial community features of Haemaphysalis flava in different developmental stages by high-throughput sequencing. J. Basic Microbiol. 2017, 57, 302–308. [Google Scholar] [CrossRef]
- Duan, D.Y.; Zhou, H.M.; Cheng, T.Y. Comparative analysis of microbial community in the whole body and midgut from fully engorged and unfed female adult Melophagus ovinus. Med. Vet. Entomol. 2020, 34, 215–224. [Google Scholar] [CrossRef]
- Tokarz, R.; Tagliafierro, T.; Sameroff, S.; Cucura, D.M.; Oleynik, A.; Che, X.; Jain, K.; Lipkin, W.I. Microbiome analysis of Ixodes saacapularis ticks from New York and Connecticut. Ticks Tick-Borne Dis. 2019, 10, 894–900. [Google Scholar] [CrossRef]
- Gurfield, N.; Grewal, S.; Cua, L.S.; Torres, P.J.; Kelley, S.T. Endosymbiont interference and microbial diversity of the Pacific coast tick, Dermacentor occidentalis, in San Diego County, California. PeerJ 2017, 5, e3202. [Google Scholar] [CrossRef]
- Kim, M.; Oh, S.; Yi, M.; Kim, M.; Yun, S.; Choi, J.H.; Yoon, M.; Yong, T.-S.; Lee, B.; Kyung, T.N.; et al. Comparative microbiome analysis of Haemaphysalis longicornis ticks at the Korea Combat Training Center in 2022. Entomol. Res. 2023, 53, 627–635. [Google Scholar] [CrossRef]
- Benyedem, H.; Lekired, A.; Mhadhbi, M.; Dhibi, M.; Romdhane, R.; Chaari, S.; Rekik, M.; Ouzari, H.I.; Hajji, T.; Darghouth, M.A. First insights into the microbiome of Tunisian Hyalomma ticks gained through next-generation sequencing with a special focus on H. scupense. PLoS ONE 2022, 17, e0268172. [Google Scholar] [CrossRef]
- Budachetri, K.; Browning, R.E.; Adamson, S.W.; Dowd, S.E.; Chao, C.C.; Ching, W.M.; Karim, S. An insight into the microbiome of the Amblyomma maculatum (Acari: Ixodidae). J. Med. Entomol. 2014, 51, 119–129. [Google Scholar] [CrossRef]
- Budachetri, K.; Gaillard, D.; Williams, J.; Mukherjee, N.; Karim, S. A snapshot of the microbiome of Amblyomma tuberculatum ticks infesting the gopher tortoise, an endangered species. Ticks Tick-Borne Dis. 2016, 7, 1225–1229. [Google Scholar] [CrossRef] [PubMed]
- Yadav, K.K.; Datta, S.; Naglot, A.; Bora, A.; Hmuaka, V.; Bhagyawant, S.; Gogoi, H.K.; Veer, V.; Raju, P.S. Diversity of cultivable midgut microbiota at different stages of the Asian tiger mosquito, Aedes albopictus from Tezpur, India. PLoS ONE 2016, 11, e0167409. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gilbreath, T.M.; Kukutla, P., 3rd; Yan, G.; Xu, J. Dynamic gut microbiome across life history of the malaria mosquito Anopheles Gambia in Kenya. PLoS ONE 2011, 6, e24767. [Google Scholar] [CrossRef]
- Oliveira, J.H.; Gonçalves, R.L.; Lara, F.A.; Dias, F.A.; Gandara, A.C.; Menna-Barreto, R.F.; Edwards, M.C.; Laurindo, F.R.; Silva-Neto, M.A.; Sorgine, M.H.; et al. Blood meal-derived heme decreases ROS levels in the midgut of Aedes aegypti and allows proliferation of intestinal microbiota. PLoS Pathog. 2011, 7, e1001320. [Google Scholar] [CrossRef]
- Kelly, P.H.; Bahr, S.M.; Serafim, T.D.; Ajami, N.J.; Petrosino, J.F.; Meneses, C.; Meneses, C.; Kirby, J.R.; Valenzuela, J.G.; Kamhawi, S.; et al. The gut microbiome of the vector Lutzomyia longipalpis is essential for survival of Leishmania infantum. mBio 2017, 8, e01121–e01216. [Google Scholar] [CrossRef]
- Telleria, E.L.; Martins-da-Silva, A.; Tempone, A.J.; Traub-Csekö, Y.M. Leishmania, microbiota and sand fly immunity. Parasitology 2018, 145, 1336–1353. [Google Scholar] [CrossRef]
- Beninati, T.; Lo, N.; Sacchi, L.; Genchi, C.; Noda, H.; Bandi, C. A novel alpha-Proteobacterium resides in the mitochondria of ovarian cells of the tick Ixodes ricinus. Appl. Environ. Microb. 2004, 70, 2596–2602. [Google Scholar] [CrossRef]
- Jensen, B.B.; Bruun, M.T.; Jensen, P.M.; Pedersen, A.K.; Fournier, P.E.; Skarphedinsson, S.; Cheng, M. Evaluation of factors influencing tick bites and tick-borne infections: A longitudinal study. Parasites Vectors 2021, 14, 289. [Google Scholar] [CrossRef]
- Lkhagvatseren, S.; Hogan, K.M.; Boldbaatar, B.; von Fricken, M.E.; Anderson, B.D.; Pulscher, L.A.; Caddell, L.; Nymadawa, P.; Gray, G.C. Discrepancies between self-reported tick bites and evidence of tick-borne disease exposure among nomadic Mongolian herders. Zoonoses Public Health 2019, 66, 480–486. [Google Scholar] [CrossRef]
- Noguchi, M.; Oshita, S.; Yamazoe, N.; Miyazaki, M.; Takemura, Y.C. Important clinical features of Japanese spotted fever. Am. J. Trop. Med. Hyg. 2018, 99, 466–469. [Google Scholar] [CrossRef]
- Gao, S.Y.; Li, L.F.; Zhou, X.L.; Dai, X.C.; Lu, L.; Chen, Y.D.; Han, J.; Huang, X.L.; Xiao, Q.; He, H.Q.; et al. Fatal Rickettsia japonica infection complicating disseminated intravascular coagulation in Yichang, China. Infect. Drug. Resist. 2022, 15, 6613–6623. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.N.; Wang, Q.; Shen, Y.J.; Shen, B.; Zhang, Y.; Wang, W.H.; Li, X.F. A case of critical japanese spotted fever in Zhejiang. Infect. Drug Resist. 2023, 16, 3425–3430. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Li, C.F.; Gao, A.; Jiang, N.; Feng, X.Y.; Li, J.; Hu, W. Evidence-practice gap analysis in the role of tick in brucellosis transmission: A scoping review. Infect. Dis. Poverty 2024, 13, 3. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhao, S.; Wureli, H.; Xie, S.; Chen, C.; Wei, Q.; Cui, B.; Tu, C.; Wang, Y. Brucella melitensis and B. abortus in eggs, larvae and engorged females of Dermacentor marginatus. Ticks Tick-Borne Dis. 2018, 9, 1045–1048. [Google Scholar] [CrossRef]
- Tovar, R.M. Infection and transmission of Brucella by ectoparasites. Am. J. Vet. Res. 1947, 8, 138–140. [Google Scholar]
- Pritulin, P.I. On the transmission of brucellosis by the pasture ticks Dermacentor nuttallia and Hyalomma marginatum. Veterinariia 1954, 7, 31–33. [Google Scholar]
- Zheng, R.J.; Xie, S.S.; Lu, X.B.; Sun, L.H.; Zhou, Y.; Zhang, Y.X.; Wang, K. A systematic review and meta-analysis of epidemiology and clinical manifestations of human brucellosis in China. Biomed Res. Int. 2018, 2018, 5712920. [Google Scholar] [CrossRef]
- Tian, Z.H.; Wan, L.Y.; Pei, J.; Li, T.T.; Wang, X.Z.; Peng, Y.; Guo, A.Z.; Chen, Y.Y. Brucellosis seroprevalence in cattle in China during 2014–2024: A systematic review and meta-analysis. Emerg. Microbes Infect. 2024, 13, 2417859. [Google Scholar] [CrossRef]
- Huang, T.P.; Zhang, J.B.; Sun, C.Y.; Liu, Z.C.; He, H.Y.; Wu, J.; Geriletu. A Novel Arthropod host of brucellosis in the arid steppe ecosystem. Front. Vet. Sci. 2020, 7, 566253. [Google Scholar] [CrossRef]
- Shi, J.M.; Hu, Z.H.; Deng, F.; Shen, S. Tick-borne viruses. Virol. Sin. 2018, 33, 21–43. [Google Scholar] [CrossRef]
- Ricci, S.; Bartolini, S.; Morandi, F.; Cuteri, V.; Preziuso, S. Genotyping of pestivirus A (Bovine Viral Diarrhea Virus 1) detected in faeces and in other specimens of domestic and wild ruminants at the wildlife-livestock interface. Vet. Microbiol. 2019, 235, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Peterhans, E.; Jungi, T.W.; Schweizer, M. BVDV and innate immunity. Biologicals 2003, 31, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.M.; Ward, M.P. Risk factors for the spread of African swine fever in China: A systematic review of Chinese-language literature. Transbound. Emerg. Dis. 2022, 69, e1289–e1298. [Google Scholar] [CrossRef]
- Wang, T.; Sun, Y.; Qiu, H.J. African swine fever: An unprecedented disaster and challenge to China. Infect. Dis. Poverty 2018, 7, 111. [Google Scholar] [CrossRef]
- Lu, G.; Pan, J.L.; Zhang, G.H. African swine fever virus in Asia: Its rapid spread and potential threat to unaffected countries. J. Infect. 2020, 80, 350–371. [Google Scholar] [CrossRef]
- Brookes, V.J.; Barrett, T.E.; Ward, M.P.; Roby, J.A.; Hernandez-Jover, M.; Cross, E.M.; Donnelly, C.M.; Barnes, T.S.; Wilson, C.S.; Khalfan, S. A scoping review of African swine fever virus spread between domestic and free-living pigs. Transbound. Emerg. Dis. 2021, 68, 2643–2656. [Google Scholar] [CrossRef]
- Plowright, W.; Perry, C.T.; Peirce, M.A.; Parker, J. Experimental infection of the argasid tick, Ornithodoros moubata porcinus, with African swine fever virus. Arch. Gesamte Virusforsch. 1970, 31, 33–50. [Google Scholar] [CrossRef]
- Plowright, W.; Perry, C.T.; Greig, A. Sexual transmission of African swine fever virus in the tick, Ornithodoros moubata porcinus, Walton. Res. Vet. Sci. 1974, 17, 106–113. [Google Scholar] [CrossRef]
- Plowright, W.; Perry, C.T.; Peirce, M.A. Transovarial infection with African swine fever virus in the argasid tick, Ornithodoros moubata porcinus, Walton. Res. Vet. Sci. 1970, 11, 582–584. [Google Scholar] [CrossRef]
- Golnar, A.J.; Martin, E.; Wormington, J.D.; Kading, R.C.; Teel, P.D.; Hamer, S.A.; Hamer, G.L. Reviewing the potential vectors and hosts of african swine fever virus transmission in the United States. Vector-Borne Zoonotic Dis. 2019, 19, 512–524. [Google Scholar] [CrossRef]
- Chen, Z.; Xu, X.F.; Wang, Y.F.; Bei, J.; Jin, X.F.; Dou, W.H.; Ji, H.; Duan, Y.; Yang, X.; Gao, S. DNA segments of African swine fever virus detected for the first time in hard ticks from sheep and bovines. Syst. Appl. Acarol. 2019, 24, 180–184. [Google Scholar] [CrossRef]
- de Carvalho Ferreira, H.C.; Tudela Zúquete, S.; Wijnveld, M.; Weesendorp, E.; Jongejan, F.; Stegeman, A.; Loeffen, W.L. No evidence of African swine fever virus replication in hard ticks. Ticks Tick-Borne Dis. 2014, 5, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, S.I.; Bouhsira, E.; De Regge, N.; Fite, J.; Etoré, F.; Garigliany, M.M.; Jori, F.; Lempereur, L.; Le Potier, M.-F.; Quillery, E.; et al. Putative role of arthropod vectors in African swine fever virus transmission in relation to their bio-ecological properties. Viruses 2020, 12, 778. [Google Scholar] [CrossRef]
- Xu, X.L.; Cheng, T.Y.; Yang, H.; Yan, F. Identification of intestinal bacterial flora in Rhipicephalus microplus ticks by conventional methods and PCR-DGGE analysis. Exp. Appl. Acarol. 2015, 66, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Williams-Newkirk, A.J.; Rowe, L.A.; Mixson-Hayden, T.R.; Dasch, G.A. Characterization of the bacterial communities of life stages of free living lone star ticks (Amblyomma americanum). PLoS ONE 2014, 9, e102130. [Google Scholar] [CrossRef]
- Clay, K.; Klyachko, O.; Grindle, N.; Civitello, D.; Oleske, D.; Fuqua, C. Microbial communities and interactions in the lone star tick, Amblyomma americanum. Mol. Ecol. 2008, 17, 4371–4381. [Google Scholar] [CrossRef]
- Swei, A.; Kwan, J.Y. Tick microbiome and pathogen acquisition altered by host blood meal. ISME J. 2017, 11, 813–816. [Google Scholar] [CrossRef]
- Heise, S.R.; Elshahed, M.S.; Little, S.E. Bacterial diversity in Amblyomma americanum (Acari: Ixodidae) with a focus on members of the genus Rickettsia. J. Med. Entomol. 2010, 47, 258–268. [Google Scholar] [CrossRef]
- Zhang, X.C.; Yang, Z.N.; Lu, B.; Ma, X.F.; Zhang, C.X.; Xu, H.J. The composition and transmission of microbiome in hard tick, Ixodes persulcatus, during blood meal. Ticks Tick-Borne Dis. 2014, 5, 864–870. [Google Scholar] [CrossRef]
- Carpi, G.; Cagnacci, F.; Wittekindt, N.E.; Zhao, F.; Qi, J.; Tomsho, L.P.; Drautz, D.I.; Rizzoli, A.; Schuster, S.C. Metagenomic profile of the bacterial communities associated with Ixodes ricinus ticks. PLoS ONE 2011, 6, e25604. [Google Scholar] [CrossRef]
- Greay, T.L.; Gofton, A.W.; Paparini, A.; Ryan, U.M.; Oskam, C.L.; Irwin, P.J. Recent insights into the tick microbiome gained through next-generation sequencing. Parasite Vectors 2018, 11, 12. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Sequences | Bases (bp) | Average Legnth (bp) |
|---|---|---|---|
| First-generation female adult ticks | 193,013 | 79,599,688 | 412.41 |
| Eggs | 77,459 | 31,356,590 | 404.82 |
| Larval ticks | 691,824 | 287,258,716 | 415.22 |
| Engorged larval ticks | 141,628 | 57,750,469 | 407.76 |
| Nymphal ticks | 78,104 | 32,874,931 | 420.91 |
| Engorged nymphal ticks | 54,276 | 23,220,668 | 427.83 |
| Second-generation adult ticks | 93,433 | 39,060,954 | 418.06 |
| Total | 1,329,737 | 551,122,016 | 414.46 |
| Sample | 97% | ||||
|---|---|---|---|---|---|
| Good’s Coverage (%) | Shannon | Simpson | Chao | ACE | |
| First-generation female adult ticks | 99.9754 | 1.70 | 0.2649 | 203 | 219 |
| Eggs | 99.9841 | 0.81 | 0.5598 | 146 | 149 |
| Larval ticks | 99.9963 | 1.98 | 0.2491 | 190 | 193 |
| Engorged larval ticks | 99.9888 | 2.25 | 0.2436 | 569 | 571 |
| Nymphal ticks | 99.9679 | 1.98 | 0.2136 | 128 | 123 |
| Engorged nymphal ticks | 99.9554 | 0.76 | 0.7510 | 144 | 147 |
| Second-generation adult ticks | 99.9706 | 2.20 | 0.1577 | 160 | 163 |
| Sample | Proteobacteria (%) | Actinobacteria (%) | Firmicutes (%) | Bacteroidetes (%) | Cyanobacteria (%) | Other (%) |
|---|---|---|---|---|---|---|
| First-generation female adult ticks | 97.45 | 1.59 | 0.75 | 0.17 | - | 0.04 |
| Eggs | 98.87 | 0.43 | 0.42 | 0.15 | 0.07 | 0.06 |
| Larval ticks | 14.19 | 56.23 | 27.30 | 2.26 | - | 0.02 |
| Engorged larval ticks | 73.26 | 16.78 | 6.53 | 1.87 | 1.13 | 0.43 |
| Nymphal ticks | 97.22 | 1.01 | 0.51 | 1.23 | - | 0.03 |
| Engorged nymphal ticks | 99.01 | 0.08 | 0.51 | 0.27 | - | 0.13 |
| Second-generation adult ticks | 87.91 | 1.44 | 7.24 | 3.22 | - | 0.19 |
| Total proportion | 81.13 | 11.08 | 6.18 | 1.31 | 0.17 | 0.13 |
| Genus | Relative Abundance (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| First-Generation Female Adult Ticks | Eggs | Larval Ticks | Engorged Larval Ticks | Nymph Ticks | Engorged Nymph Ticks | Second-Generation Adult Ticks | Total Proportion | |
| Rickettsia | 38.708 | 69.596 | 2.714 | 3.827 | 30.166 | 3.143 | 15.885 | 27.720 |
| Candidatus Lariskella | 26.870 | 27.503 | 1.036 | 34.363 | 0.028 | 0.034 | 25.380 | 16.459 |
| Pseudomonas | 3.245 | 0.393 | 0.767 | 0.259 | 0.459 | 88.883 | 13.513 | 15.360 |
| Enterobacteriaceae unclassified | 0.546 | 0.263 | 0.006 | 0.042 | 33.252 | 0.170 | 20.950 | 7.890 |
| Brevibacterium | 0.304 | - | 43.261 | 9.384 | 0.038 | 0.011 | 0.030 | 7.575 |
| Stenotrophomonas | 21.345 | 0.200 | 0.087 | 0.136 | 4.198 | 1.138 | 6.662 | 4.827 |
| Staphylococcus | 0.367 | 0.028 | 26.229 | 1.123 | 0.200 | 0.085 | 0.004 | 4.005 |
| Acinetobacter | 0.049 | - | 0.002 | 0.189 | 10.03 | 0.062 | 0.677 | 1.573 |
| Serratia | 0.017 | - | - | 0.017 | 10.835 | 0.049 | 0.028 | 1.564 |
| Dietzia | 0.223 | - | 7.700 | 2.935 | - | - | 0.049 | 1.558 |
| Brucella | 1.487 | 0.006 | 5.139 | 1.111 | 0.085 | 0.066 | 1.491 | 1.341 |
| Achromobacter | 4.432 | - | 0.113 | 0.0347 | 0.023 | 0.837 | 2.364 | 1.115 |
| Lysinibacillus | 0.104 | - | - | 0.282 | 0.011 | 0.038 | 7.161 | 1.085 |
| Delftia | 0.057 | 0.017 | 0.202 | 0.074 | 4.279 | 2.801 | 0.057 | 1.070 |
| Brevundimonas | 0.093 | 0.013 | 1.368 | 0.890 | 3.321 | 0.357 | 0.342 | 0.912 |
| Brachybacterium | 0.062 | - | 3.544 | 0.52 | 0.454 | 0.004 | 0.015 | 0.657 |
| Sphingobacterium | 0.087 | - | 2.183 | 0.242 | 0.750 | 0.042 | 0.973 | 0.611 |
| Rhodococcus | 0.741 | 0.021 | 0.083 | 1.213 | 0.189 | 0.036 | 1.138 | 0.489 |
| Advenella | 0.040 | - | 2.164 | 0.106 | 0.011 | - | 0.002 | 0.332 |
| Jeotgalicoccus | 0.015 | - | 0.66 | 1.567 | 0.055 | - | - | 0.328 |
| Chryseobacterium | 0.002 | 0.002 | - | 0.028 | 0.021 | 0.019 | 2.034 | 0.301 |
| Corynebacterium | - | - | 0.032 | 1.867 | 0.181 | 0.006 | - | 0.298 |
| Microbacterium | 0.036 | - | 1.255 | 0.070 | - | - | 0.047 | 0.201 |
| Chloroplast_norank | - | 0.021 | - | 1.075 | - | - | - | 0.157 |
| EscherichiaShigella | - | 0.567 | 0.004 | 0.053 | 0.002 | 0.023 | - | 0.093 |
| Flavobacterium | - | 0.002 | - | 0.0539 | - | 0.002 | 0.104 | 0.093 |
| Psychrobacter | 0.079 | - | - | 0.539 | - | - | 0.017 | 0.091 |
| Comamonas | 0.002 | - | - | - | 0.089 | 0.459 | 0.066 | 0.088 |
| Bacillus | 0.104 | 0.117 | 0.053 | 0.183 | 0.006 | 0.127 | 0.011 | 0.086 |
| Bacteroides | 0.004 | 0.053 | 0.002 | 0.406 | 0.064 | 0.021 | 0.015 | 0.081 |
| Bacterial Species | Relative Abundance (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| First-Generation Female Adult Ticks | Eggs | Larval Ticks | Engorged Larval Ticks | Nymph Ticks | Engorged Nymph Ticks | Second-Generation Adult Ticks | Total Abundance | |
| Rickettsia japonica | 38.708 | 69.596 | 2.714 | 33.827 | 30.166 | 3.143 | 15.885 | 27.720 |
| Ixodes persulcatus taiga tick | 26.870 | 27.503 | 1.036 | 34.363 | 0.028 | 0.034 | 25.380 | 16.459 |
| Pseudomonas aeruginosa | 0.074 | 0 | 0.134 | 0.017 | 0.151 | 86.517 | 0.212 | 12.444 |
| Enterobacteriaceae unclassified | 0.546 | 0.263 | 0.006 | 0.042 | 33.252 | 0.170 | 20.950 | 7.890 |
| Brevibacterium epidermidis | 0.304 | 0 | 43.248 | 9.382 | 0.038 | 0.011 | 0.030 | 7.573 |
| Stenotrophomonas sp. MYb57 | 20.338 | 0.025 | 0.068 | 0.036 | 2.440 | 0.087 | 6.464 | 4.208 |
| Staphylococcus lentus | 0.329 | 0.013 | 23.309 | 1.068 | 0.151 | 0.040 | 0 | 3.559 |
| Pseudomonas unclassified | 2.395 | 0.316 | 0.626 | 0.106 | 0.282 | 2.130 | 11.803 | 2.523 |
| Serratia marcescens | 0 | 0 | 0 | 0.017 | 10.826 | 0.049 | 0.015 | 1.558 |
| Brucella unclassified | 1.487 | 0.006 | 5.139 | 1.111 | 0.085 | 0.066 | 1.491 | 1.341 |
| Acinetobacter unclassified | 0.002 | 0 | 0.002 | 0.181 | 8.807 | 0.051 | 0.093 | 1.305 |
| Achromobacter unclassified | 4.432 | 0 | 0.113 | 0.034 | 0.023 | 0.837 | 2.364 | 1.115 |
| Delftia tsuruhatensis | 0.057 | 0.017 | 0.202 | 0.074 | 4.279 | 2.801 | 0.057 | 1.070 |
| Lysinibacillus_unclassified | 0.104 | 0 | 0 | 0.002 | 0.011 | 0.038 | 7.161 | 1.045 |
| Sample | Kingdom | Phylum | Class | Order | Family | Genus | Species |
|---|---|---|---|---|---|---|---|
| First-generation adult ticks | norank | norank | norank | 6 | 27 | 64 | 136 |
| Mixed sample | norank | norank | norank | 5 | 23 | 51 | 90 |
| Second-generation adult ticks | norank | norank | norank | 5 | 27 | 71 | 143 |
| Total | - | - | - | 6 | 28 | 72 | 158 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Dong, X.; Sun, L.; Cui, H.; Kang, J.; Bu, N.; Zhang, Y.; Qi, Z.; Li, Z.; Zhang, Z.; et al. Analysis of the Microbial Community Structure of Ixodes persulcatus at Each Developmental Stage. Animals 2025, 15, 830. https://doi.org/10.3390/ani15060830
Liu Y, Dong X, Sun L, Cui H, Kang J, Bu N, Zhang Y, Qi Z, Li Z, Zhang Z, et al. Analysis of the Microbial Community Structure of Ixodes persulcatus at Each Developmental Stage. Animals. 2025; 15(6):830. https://doi.org/10.3390/ani15060830
Chicago/Turabian StyleLiu, Yonghong, Xiaonan Dong, Lianyang Sun, Hao Cui, Jiamei Kang, Nan Bu, Yishuai Zhang, Zehao Qi, Zixuan Li, Zilong Zhang, and et al. 2025. "Analysis of the Microbial Community Structure of Ixodes persulcatus at Each Developmental Stage" Animals 15, no. 6: 830. https://doi.org/10.3390/ani15060830
APA StyleLiu, Y., Dong, X., Sun, L., Cui, H., Kang, J., Bu, N., Zhang, Y., Qi, Z., Li, Z., Zhang, Z., & Zhao, L. (2025). Analysis of the Microbial Community Structure of Ixodes persulcatus at Each Developmental Stage. Animals, 15(6), 830. https://doi.org/10.3390/ani15060830