




Changes in Core Temperature of Cyan-Shank Partridge Chickens Exposed to Continuously Increased Ambient Temperature at Different Relative Humidity Levels

 ,
,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Birds and Treatments
2.2. Data Measurements
2.3. Statistical Analysis
3. Results
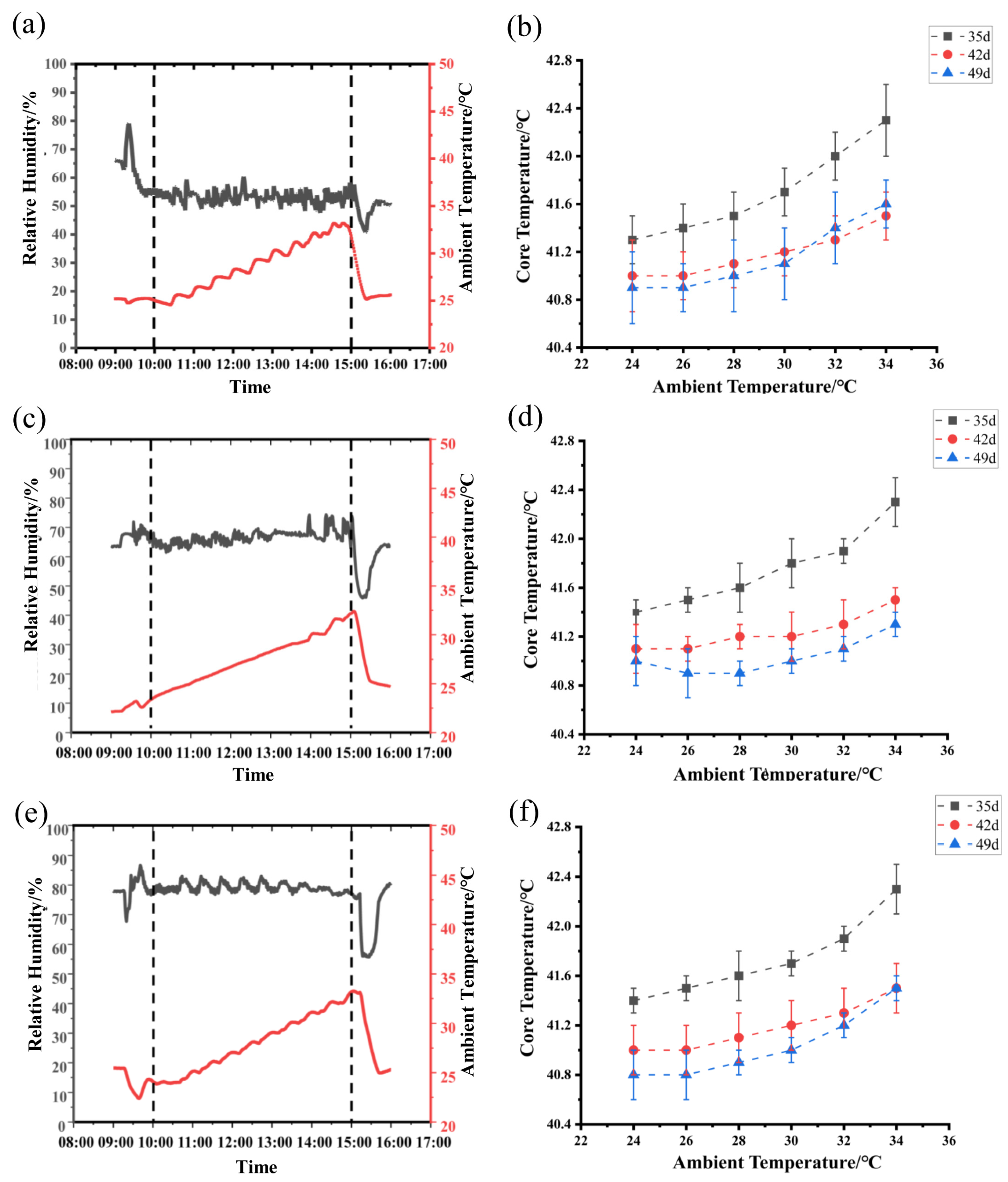
3.1. Changes in Chamber AT and Bird Core Temperature as Exposed to Different RH Levels
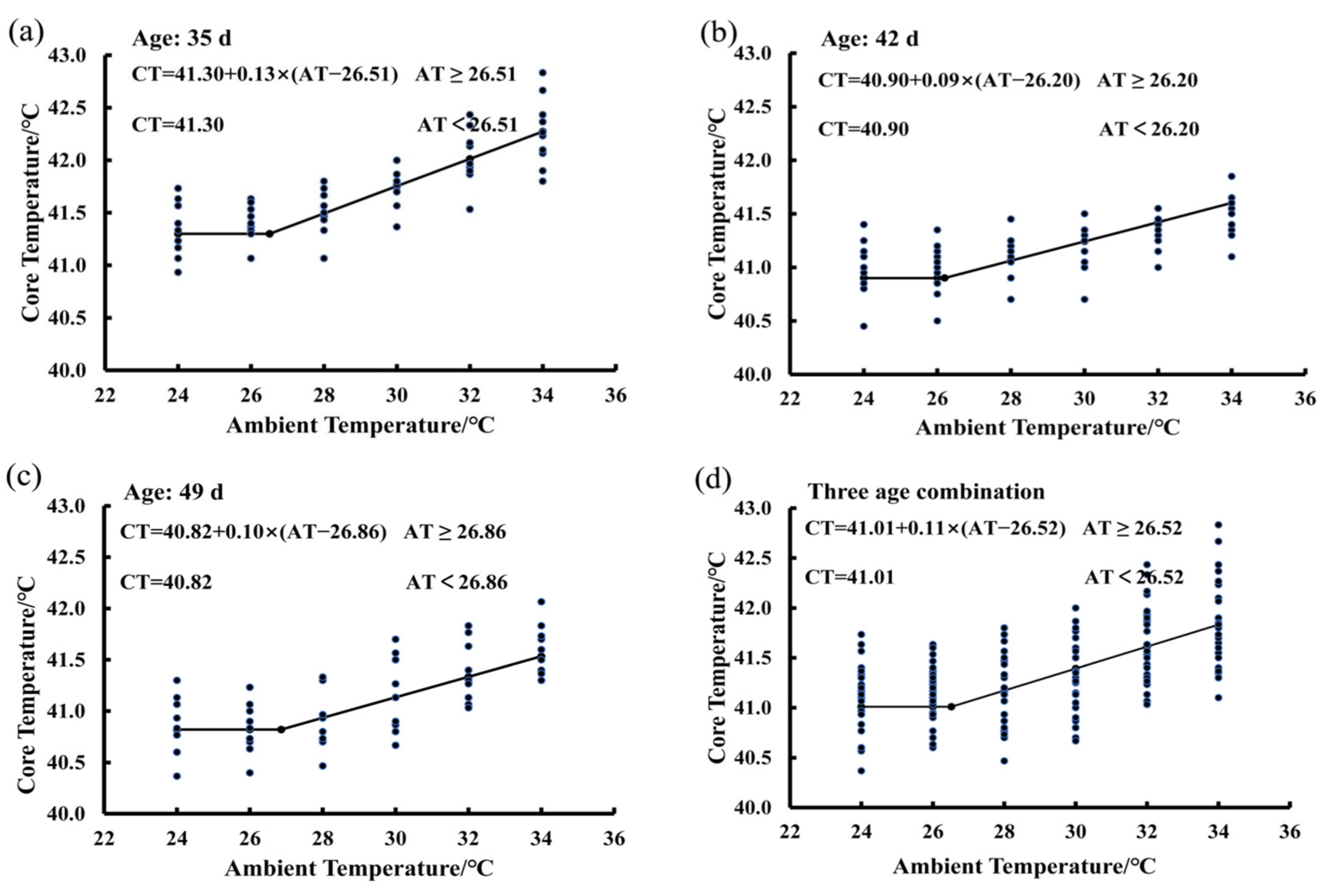
3.2. Estimating IPT Based on the Core Temperature Variations of Birds Exposed to the 50% RH Level
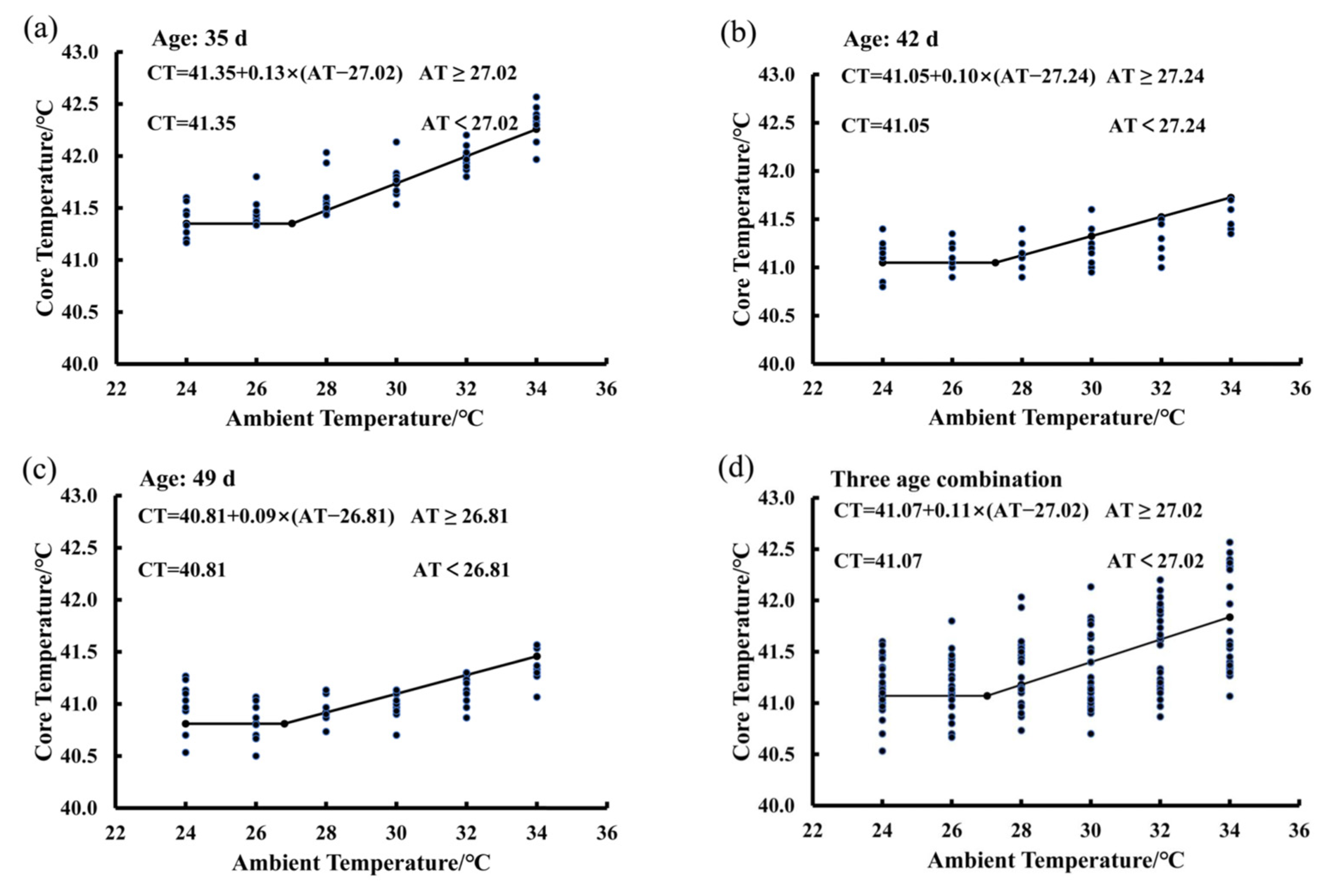
3.3. Estimating IPT Based on the Core Temperature Variations of Birds Exposed to the 65% RH Level
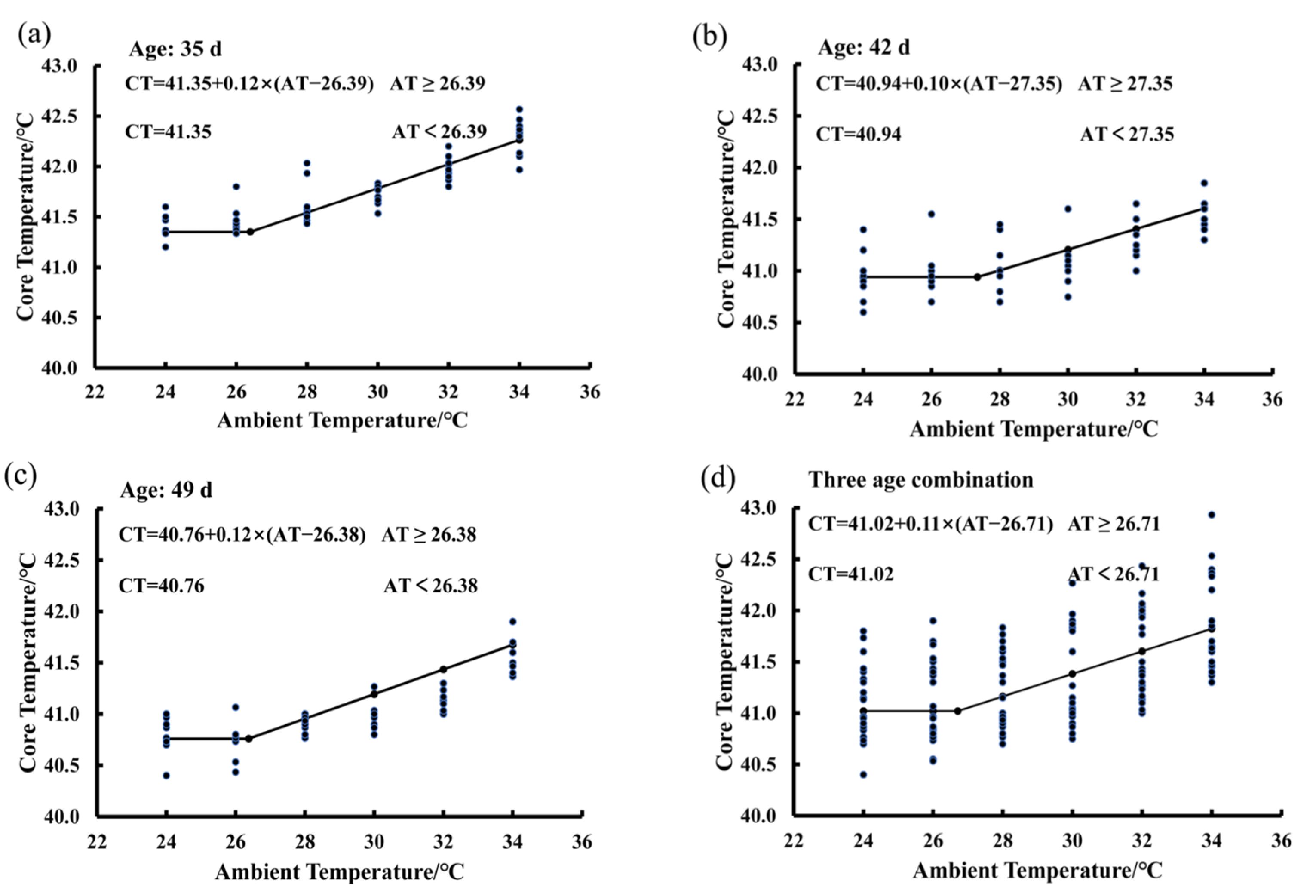
3.4. Estimating IPT Based on the Core Temperature Variations of Birds Exposed to the 80% RH Level
3.5. Effect of RH and Age on the IPT of Birds Exposed to the Increasing Ambient Temperature
4. Discussion
4.1. Changes in Core Temperature and IPT as Exposed to the Increasing Ambient Temperature
4.2. Changes in Slope in BLM with the Increasing Ambient Temperature and Age
4.3. Changes in the Parameters in BLM as Exposed to the Increasing RH
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, G.; Li, X.; Zhou, Y.; Feng, J.; Zhang, M.J.A. Effects of heat stress on gut-microbial metabolites, gastrointestinal peptides, glycolipid metabolism, and performance of broilers. Animals 2021, 11, 1286. [Google Scholar] [CrossRef] [PubMed]
- Hosseini-Vashan, S.J.; Safdari-Rostamabad, M.; Piray, A.H.; Sarir, H.J.A.F.S. The growth performance, plasma biochemistry indices, immune system, antioxidant status, and intestinal morphology of heat-stressed broiler chickens fed grape (Vitis vinifera) pomace. Anim. Feed. Sci. Technol. 2020, 259, 114343. [Google Scholar] [CrossRef]
- Hu, J.Y.; Mohammed, A.A.; Murugesan, G.R.; Cheng, H.W. Effect of a synbiotic supplement as an antibiotic alternative on broiler skeletal, physiological, and oxidative parameters under heat stress. Poult. Sci. 2022, 101, 101769. [Google Scholar] [CrossRef]
- Sun, S.; Li, B.; Wu, M.; Deng, Y.; Li, J.; Xiong, Y.; He, S. Effect of dietary supplemental vitamin C and betaine on the growth performance, humoral immunity, immune organ index, and antioxidant status of broilers under heat stress. Trop. Anim. Health Prod. 2023, 55, 96. [Google Scholar] [CrossRef]
- Wang, M.; Wang, L.; Wang, C.Q.; Wan, S.H.; Wang, H.; Sun, J.Y.; Tang, J.Q.; Zhang, X.C.; Zhang, H.J.; Li, J.; et al. Complete mitochondrial genome of the Partridge Shank chicken (Galliformes: Phasianidae). Mitochondrial DNA Part B Resour. 2019, 4, 3298–3300. [Google Scholar] [CrossRef]
- Yuan, C.; Jiang, Y.; Wang, Z.; Chen, G.; Bai, H.; Chang, G. Indigenous, Yellow-feathered chickens body measurements, carcass traits, and meat quality depending on marketable age. Animals 2022, 12, 2422. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.R.; Chen, Y.P.; Cheng, Y.F.; Qu, H.M.; Li, J.; Wen, C.; Zhou, Y.M. Effects of dietary phytosterols on growth performance, antioxidant status, and meat quality in Partridge Shank chickens. Poult. Sci. 2019, 98, 3715–3721. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, H.; Li, S.; Jiang, Y.; Deng, J.; Yang, C.; Chen, X.; Jiang, L. Effects of different levels of Citri Sarcodactylis Fructus by-products fermented feed on growth performance, serum biochemical, and intestinal health of cyan-shank partridge birds. Sci. Rep. 2023, 13, 20130. [Google Scholar] [CrossRef]
- Li, Y.; Ma, R.; Qi, R.; Li, H.; Li, J.; Liu, W.; Wan, Y.; Liu, Z.; Li, S.; Chang, X.; et al. Study on the changing patterns of production performance of laying hens and their relationships with environmental factors in a large-scale henhouse. Poult. Sci. 2024, 103, 104185. [Google Scholar] [CrossRef]
- Peng, H.; Wang, Y.; Zhang, Z.; Qin, W.; Li, B.; Zheng, W.; Yin, P.; Zhu, H. Effects of low-pressure systems on temperature, humidity, egg production, and feed utilization efficiency in large-scale poultry houses during summer. Animals 2024, 14, 2554. [Google Scholar] [CrossRef]
- Pontara Vilas Boas Ribeiro, B.; Yanagi Junior, T.; Duarte de Oliveira, D.; Ribeiro de Lima, R.; Gilberto Zangeronimo, M. Thermoneutral zone for laying hens based on environmental conditions, enthalpy and thermal comfort indexes. J. Therm. Biol. 2020, 93, 102678. [Google Scholar] [CrossRef] [PubMed]
- IUPS. Glossary of terms for thermal physiology. Jpn. J. Physiol. 2001, 51, 245. [Google Scholar]
- Pallubinsky, H.; Schellen, L.; van Marken Lichtenbelt, W.D. Exploring the human thermoneutral zone—A dynamic approach. J. Therm. Biol. 2019, 79, 199–208. [Google Scholar] [CrossRef]
- Huynh, T.T.T.; Aarnink, A.J.A.; Verstegen, M.W.A.; Gerrits, W.J.J.; Heetkamp, M.J.W.; Kemp, B.; Canh, T.T. Effects of increasing temperatures on physiological changes in pigs at different relative humidities. J. Anim. Sci. 2005, 83, 1385–1396. [Google Scholar] [CrossRef]
- Pilgrim, C. piecewise-regression (aka segmented regression) in Python. J. Open Source Softw. 2021, 6, 3859. [Google Scholar] [CrossRef]
- Gordon, C.J. Thermal physiology of laboratory mice: Defining thermoneutrality. J. Therm. Biol. 2012, 37, 654–685. [Google Scholar] [CrossRef]
- Chang, Y.; Wang, X.J.; Feng, J.H.; Zhang, M.H.; Diao, H.J.; Zhang, S.S.; Peng, Q.Q.; Zhou, Y.; Li, M.; Li, X. Real-time variations in body temperature of laying hens with increasing ambient temperature at different relative humidity levels. Poult. Sci. 2018, 97, 3119–3125. [Google Scholar] [CrossRef]
- Lin, H.; Zhang, H.; Du, R.; Gu, X.; Zhang, Z.; Buyse, J.; Decuypere, E. Thermoregulation responses of broiler chickens to humidity at different ambient temperatures. II. Four weeks of age. Poult. Sci. 2005, 84, 1173–1178. [Google Scholar] [CrossRef]
- Abo Ghanima, M.M.; Bin-Jumah, M.; Abdel-Moneim, A.-M.E.; Khafaga, A.F.; Abd El-Hack, M.E.; Allam, A.A.; El-Kasrawy, N.I. Impacts of strain variation on response to heat stress and boldo extract supplementation to broiler chickens. Animals 2019, 10, 24. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, X.; Zhang, M.; Feng, J. The upper limit temperature of thermoneutral zone estimated by the changes of temperature and respiration rate of the broilers. Sci. Agric. Sin. 2019, 52, 550–557. [Google Scholar]
- Bjerg, B.; Zhang, G.; Pedersen, P.; Morsing, S. Effective temperature for poultry and pigs in hot climate. In Animal Husbandry and Nutrition, 1st ed.; Yücel, B., Taşkin, T., Eds.; IntechOpen: London, UK, 2018; Chapter 2; pp. 23–40. [Google Scholar] [CrossRef]
- NRC. Nutrition Requirement of Poultry; National Academy Press: Washington, DC, USA, 2012. [Google Scholar]
- Yehia, M.; Askri, A.; Achour, A.; Prus, J.A.; Ouellet, V.; Alnahhas, N. Rectal temperature and heat transfer dynamics in the eye, face, and breast of broiler chickens exposed to moderate heat stress. Poult. Sci. 2024, 104, 104748. [Google Scholar] [CrossRef] [PubMed]
- Herborn, K.A.; Graves, J.L.; Jerem, P.; Evans, N.P.; Nager, R.; McCafferty, D.J.; McKeegan, D.E. Skin temperature reveals the intensity of acute stress. Physiol. Behav. 2015, 152, 225–230. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Wang, K.; Li, Y.; Nagaoka, K.; Li, C. Gut microbiota intervention attenuates thermogenesis in broilers exposed to high temperature through modulation of the hypothalamic 5-HT pathway. J. Anim. Sci. Biotechnol. 2023, 14, 159. [Google Scholar] [CrossRef]
- Nascimento, G.; Nääs, I.; Pereira, D.F.; Baracho, M.; Garcia, R. Assessment of broiler surface temperature variation when exposed to different air temperatures. Braz. J. Poult. Sci. 2011, 13, 259–263. [Google Scholar] [CrossRef]
- Kim, D.-H.; Lee, Y.-K.; Lee, S.-D.; Kim, S.-H.; Lee, K.-W. Physiological and behavioral responses of laying hens exposed to long-term high temperature. J. Therm. Biol. 2021, 99, 103017. [Google Scholar] [CrossRef] [PubMed]
- Tzschentke, B.; Nichelmann, M.; Postel, T. Effects of ambient temperature, age and wind speed on the thermal balance of layer-strain fowls. Br. Poult. Sci. 1996, 37, 501–520. [Google Scholar] [CrossRef]
- Perini, F.; Cendron, F.; Rovelli, G.; Castellini, C.; Cassandro, M.; Lasagna, E. Emerging genetic tools to investigate molecular pathways related to heat stress in chickens: A Review. Animals 2021, 11, 46. [Google Scholar] [CrossRef]
- Riber, A.B.; Wurtz, K.E. Impact of growth rate on the welfare of broilers. Animals 2024, 14, 3330. [Google Scholar] [CrossRef]
- Christensen, K.; Thaxton, Y.V.; Thaxton, J.P.; Scanes, C.G. Changes in body temperature during growth and in response to fasting in growing modern meat type chickens. Br. Poult. Sci. 2012, 53, 531–537. [Google Scholar] [CrossRef]
- Adu-Asiamah, P.; Zhang, Y.; Amoah, K.; Leng, Q.Y.; Zheng, J.H.; Yang, H.; Zhang, W.L.; Zhang, L. Evaluation of physiological and molecular responses to acute heat stress in two chicken breeds. Animals 2021, 15, 100106. [Google Scholar] [CrossRef]
- Soleimani, A.F.; Zulkifli, I.; Omar, A.R.; Raha, A.R. Physiological responses of 3 chicken breeds to acute heat stress. Poult. Sci. 2011, 90, 1435–1440. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Xing, C.H.; Wei, W.; Zhang, X.F.; Wei, Z.Y.; Ren, L.L.; Jiang, J.J.; Li, M.; Wang, J.X.; He, X.X.; et al. Genome-wide scan for selection signatures and genes related to heat tolerance in domestic chickens in the tropical and temperate regions in Asia. Poult. Sci. 2022, 101, 101821. [Google Scholar] [CrossRef] [PubMed]
- Ncho, C.M.; Gupta, V.; Goel, A. Effect of thermal conditioning on growth performance and thermotolerance in broilers: A systematic review and meta-analysis. J. Therm. Biol. 2021, 98, 102916. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | (IPT)/°C | Z | Constant/°C | |
|---|---|---|---|---|
| RH | 50% | 26.52 ± 1.69 | 0.11 ± 0.04 | 41.01 ± 0.30 |
| 65% | 27.02 ± 1.83 | 0.11 ± 0.04 | 41.07 ± 0.29 | |
| 80% | 26.71 ± 1.81 | 0.11 ± 0.03 | 41.02 ± 0.32 | |
| Age | 35 days | 26.64 ± 1.72 | 0.13 ± 0.03 a | 41.33 ± 0.20 a |
| 42 days | 26.93 ± 1.88 | 0.10 ± 0.03 b | 40.96 ± 0.20 b | |
| 49 days | 26.68 ± 1.77 | 0.10 ± 0.04 b | 40.80 ± 0.19 c | |
| p-value | ||||
| RH | 0.560 | 0.672 | 0.430 | |
| Age | 0.801 | <0.001 | <0.001 | |
| RH × Age | 0.659 | 0.150 | 0.728 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Chen, K.; Xu, H.; Liu, L.; Liu, L.; Li, C.; Li, Y. Changes in Core Temperature of Cyan-Shank Partridge Chickens Exposed to Continuously Increased Ambient Temperature at Different Relative Humidity Levels. Animals 2025, 15, 820. https://doi.org/10.3390/ani15060820
Wang C, Chen K, Xu H, Liu L, Liu L, Li C, Li Y. Changes in Core Temperature of Cyan-Shank Partridge Chickens Exposed to Continuously Increased Ambient Temperature at Different Relative Humidity Levels. Animals. 2025; 15(6):820. https://doi.org/10.3390/ani15060820
Chicago/Turabian StyleWang, Chen, Ketian Chen, Haocong Xu, Le Liu, Longshen Liu, Chunmei Li, and Yansen Li. 2025. "Changes in Core Temperature of Cyan-Shank Partridge Chickens Exposed to Continuously Increased Ambient Temperature at Different Relative Humidity Levels" Animals 15, no. 6: 820. https://doi.org/10.3390/ani15060820
APA StyleWang, C., Chen, K., Xu, H., Liu, L., Liu, L., Li, C., & Li, Y. (2025). Changes in Core Temperature of Cyan-Shank Partridge Chickens Exposed to Continuously Increased Ambient Temperature at Different Relative Humidity Levels. Animals, 15(6), 820. https://doi.org/10.3390/ani15060820