
Immunological Responses and Protection in the Largemouth Bass (Microterus salmoides) Immunized with Inactivated Vaccine Against Largemouth Bass Ranavirus (LMBRaV)

 , , ,
, , ,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cell Lines and Virus
2.3. Inactivation of LMBRaV
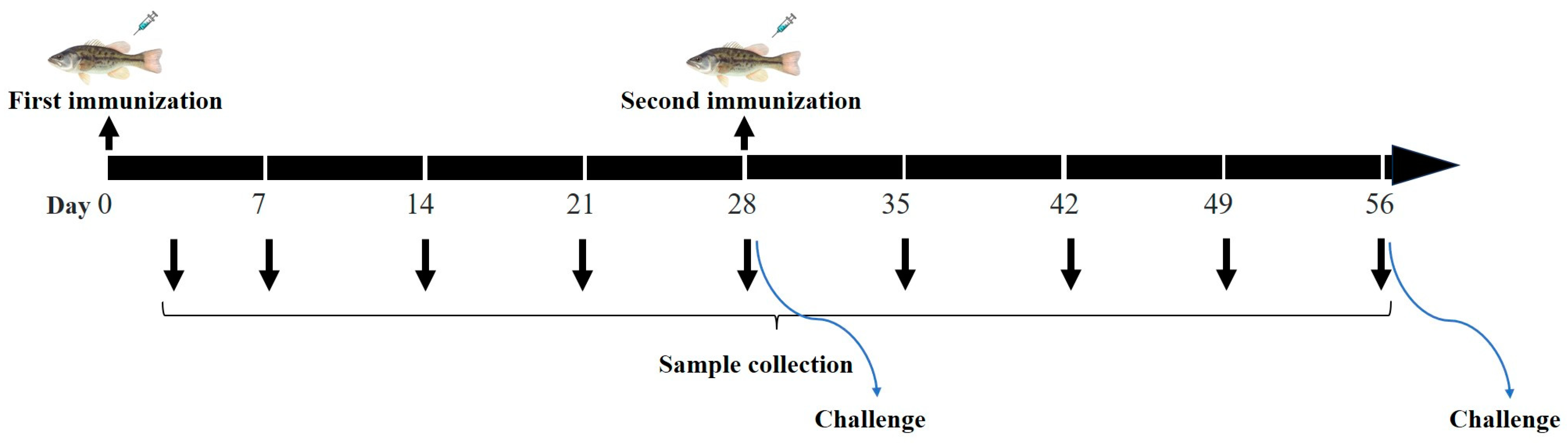
2.4. Immunization and Samples Collection
2.5. Blood Parameters
2.6. Serum Neutralization Test
2.7. Quantitative Real-Time Reverse Transcriptase PCR (qRT-PCR)
2.8. LMBRaV Challenge
2.9. Statistical Analysis
3. Results
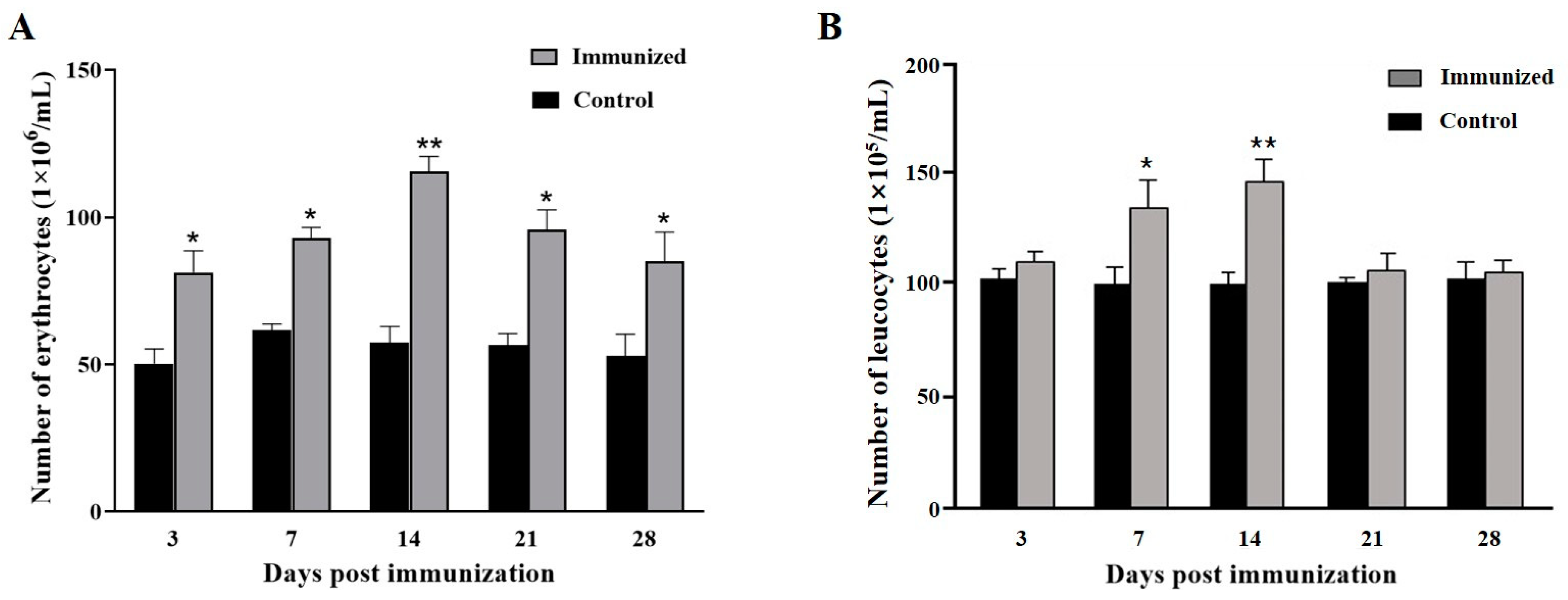
3.1. Blood Cell Counting
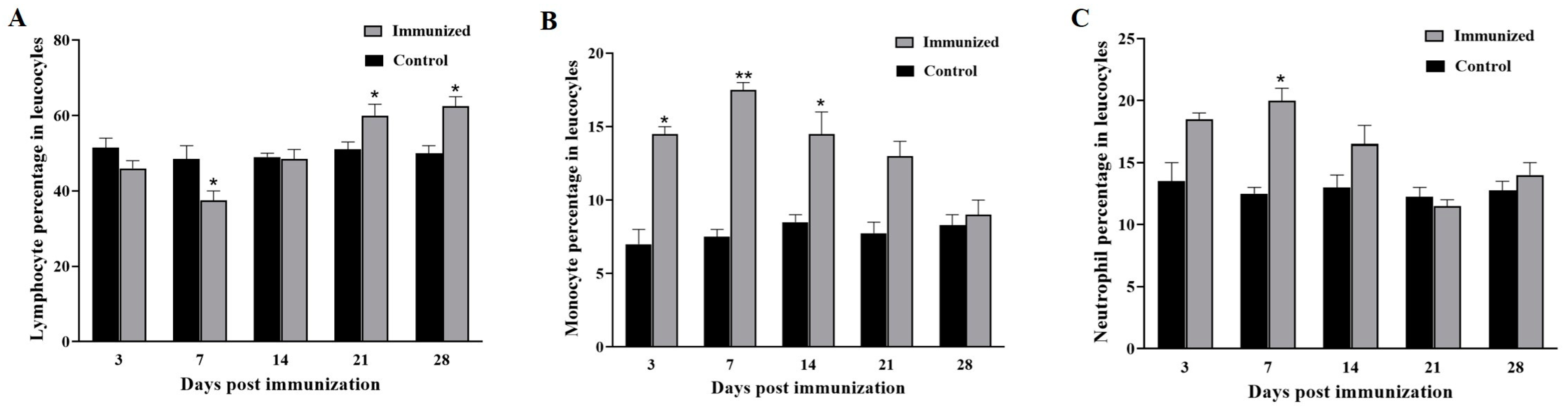
3.2. Different Leucocytes Count
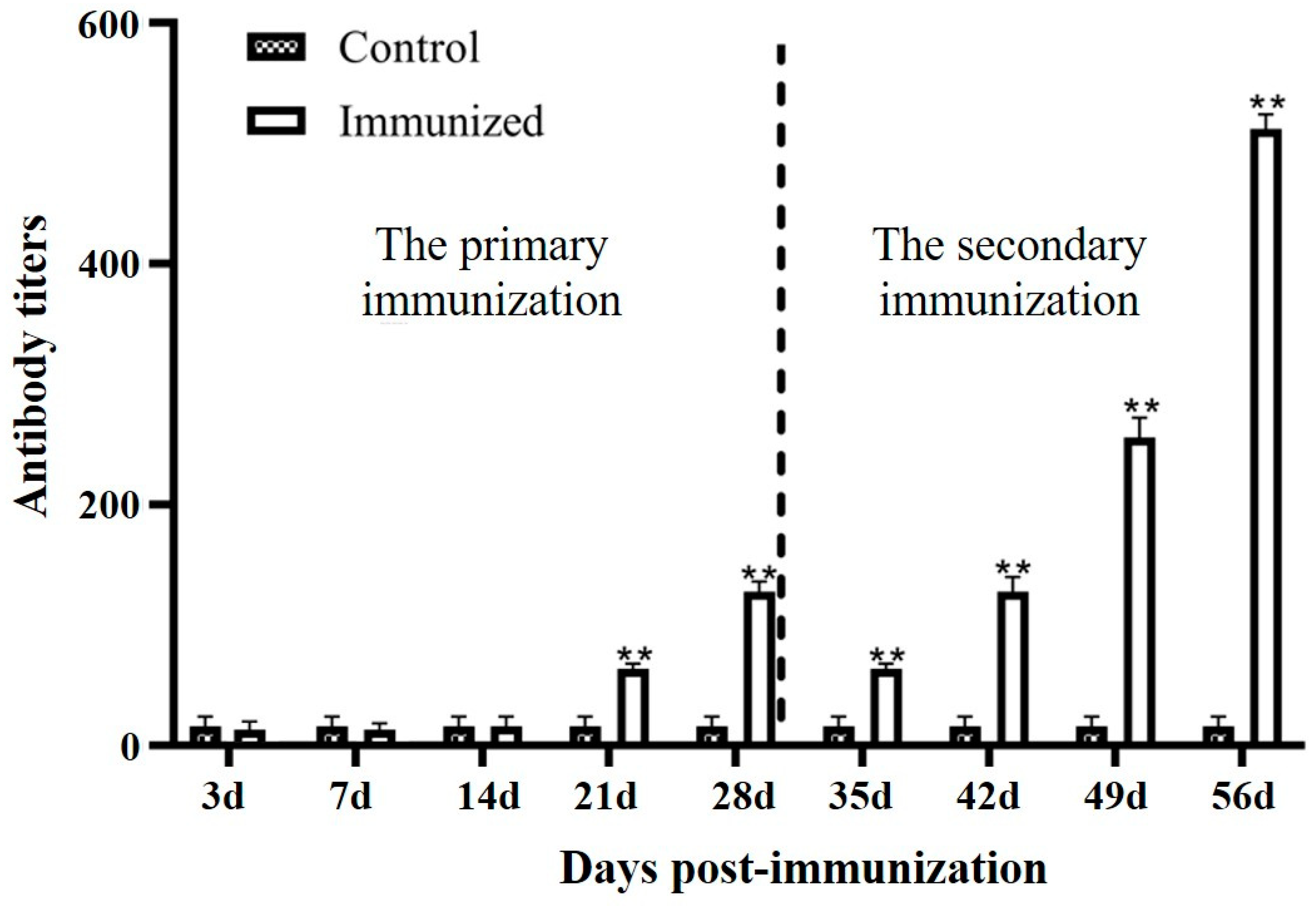
3.3. Serum Antibody Levels
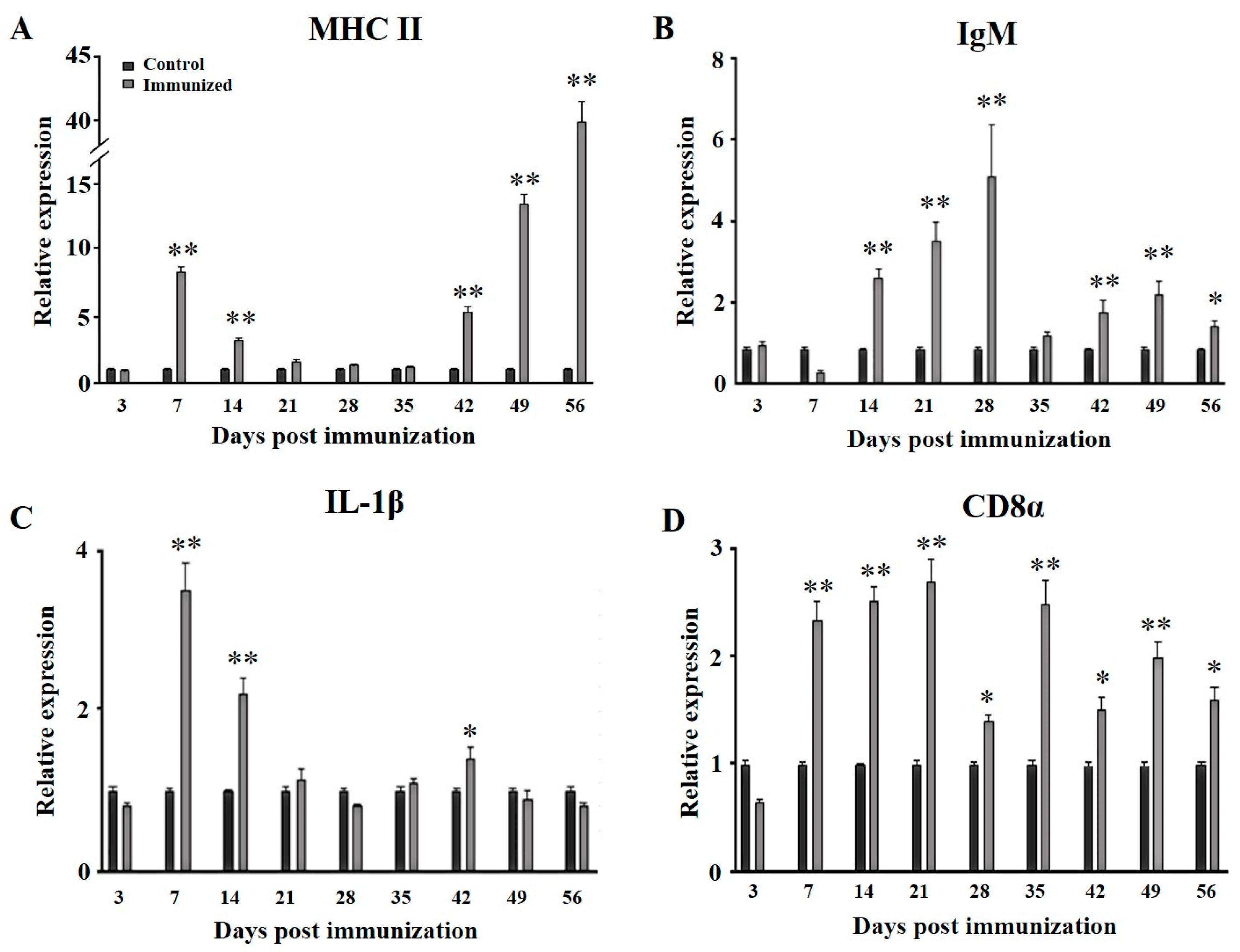
3.4. Transcription Changes of Immune Related Genes
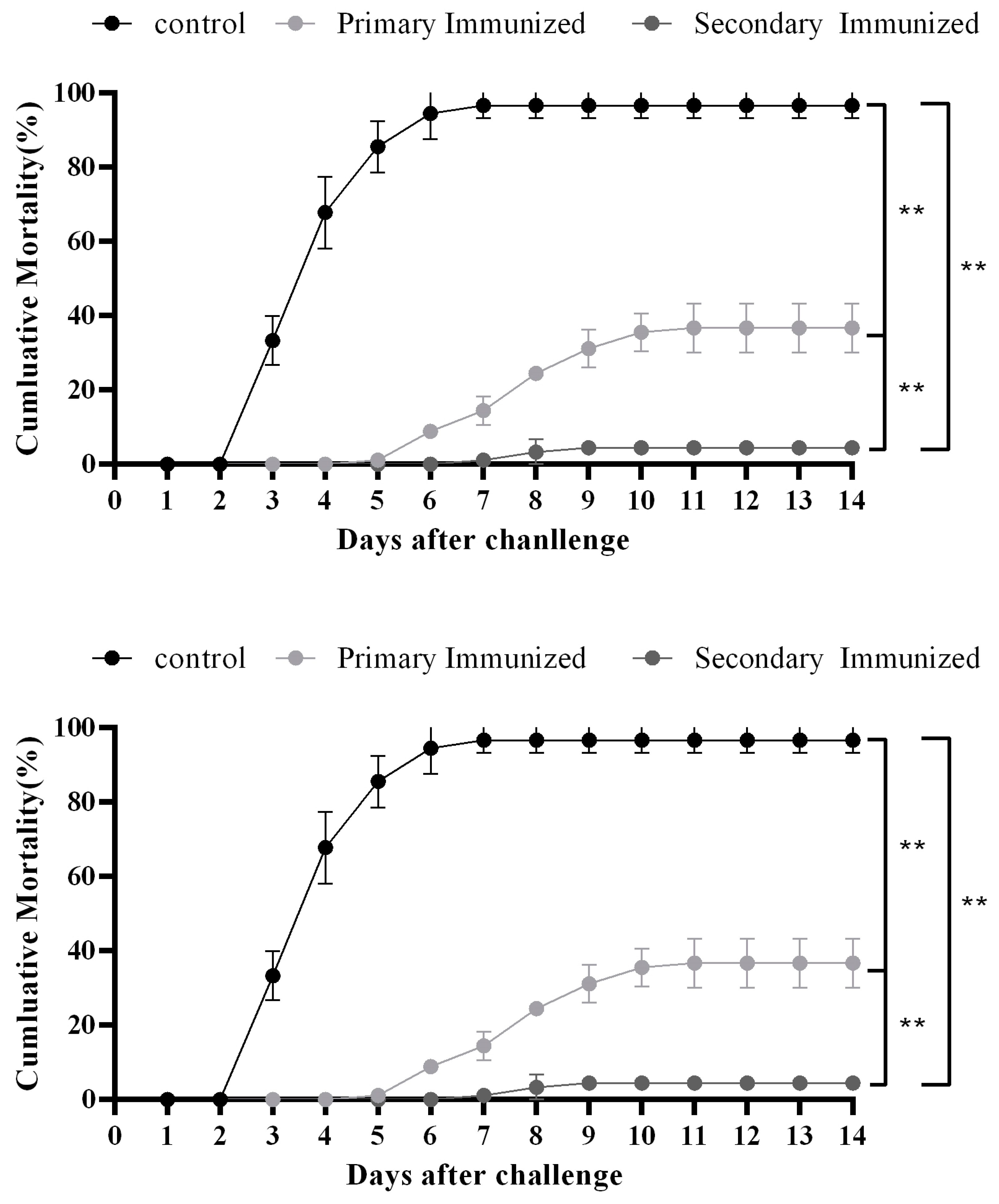
3.5. Challenge Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Q.; Li, K.; Zeng, W.; Liu, C.; Shi, C.; Wu, S. Progress on viral disease caused by largemouth bass ranavirus. Prog. Vet. Med. 2011, 32, 73–76. [Google Scholar] [CrossRef]
- Fisheries Bureau of the Ministry of Agriculture and Rural Affairs, National Fisheries Technology Extension Center & China Society of Fisheries. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2023; p. 25. [Google Scholar]
- Deng, G.C.; Li, S.J.; Xie, J.; Bai, J.J.; Chen, K.C.; Ma, D.M.; Jiang, X.Y.; Lao, H.H.; Yu, L.Y. Characterization of a ranavirus isolated from cultured largemouth bass (Micropterus salmoides) in China. Aquaculture 2011, 312, 198–204. [Google Scholar] [CrossRef]
- Zhao, R.X.; Geng, Y.; Qin, Z.Y.; Wang, K.Y.; Ouyang, P.; Chen, D.F.; Huang, X.L.; Zuo, Z.C.; He, C.L.; Guo, H.R.; et al. A new ranavirus of the Santee-Cooper group invades largemouth bass (Micropterus salmoides) culture in southwest China. Aquaculture 2020, 526, 735363. [Google Scholar] [CrossRef]
- Luo, X.W.; Shen, J.Y.; Yang, T.; Zeng, L.B.; Zhou, Y.; Fan, Y.D.; Liu, W.Z.; Li, Y.Q.; Xue, M.Y.; Meng, Y.; et al. Isolation and identification of largemouth bass ranavirus from Hubei Province. J. Fish Sci. China 2022, 29, 494–502. [Google Scholar] [CrossRef]
- Plumb, J.A.; Grizzle, J.M.; Young, H.E.; Noyes, A.D.; Lamprecht, S. An iridovirus isolated from wild largemouth bass. J. Aquat. Anim. Health 1996, 8, 265–270. [Google Scholar] [CrossRef]
- Mao, J.H.; Wang, J.; Chinchar, G.D.; Chinchar, V.G. Molecular characterization of a ranavirus isolated from largemouth bass Micropterus salmoides. Dis. Aquat. Org. 1999, 37, 107–114. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Hick, P.; Ince, I.A.; Jancovich, J.K.; Marschang, R.; Qin, Q.W.; Subramaniam, K.; Waltzek, T.B.; Whittington, R.; Williams, T.; et al. ICTV virus taxonomy profile: Iridoviridae. J. Gen. Virol. 2017, 98, 890–891. [Google Scholar] [CrossRef]
- Getchell, R.G.; Groocock, G.H.; Schumacher, V.L.; Grimmett, S.G.; Wooster, G.A.; Bowser, P.R. Quantitative polymerase chain reaction assay for largemouth bass virus. J. Aquat. Anim. Health 2007, 19, 226–233. [Google Scholar] [CrossRef]
- Ma, D.; Bai, J.; Deng, G.C.; Li, S.J.; Ye, X.; Jiang, X. Sequence analysis of MCP gene from largemouth bass ulcerative syndrome virus and rapid detection by PCR assay. J. Fish Sci. China 2010, 17, 1149–1156. [Google Scholar]
- Zhu, Q.C.; Wang, Y.; Feng, J.L. Rapid diagnosis of largemouth bass ranavirus in fish samples using the loop-mediated isothermal amplification method. Mol. Cell. Probes. 2020, 52, 101569. [Google Scholar] [CrossRef]
- Jiang, N.; Shen, J.Y.; Zhou, Y.; Liu, W.Z.; Meng, Y.; Li, Y.Q.; Xue, M.Y.; Xu, C.; Fan, Y.D. Development of a droplet digital PCR method for the sensitive detection and quantification of largemouth bass ranavirus. J. Fish Dis. 2023, 46, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Z.L.; Wang, P.R.; Huang, X.H.; Cai, J.; Huang, Y.H.; Wei, S.; Ji, H.S.; Wei, J.G.; Zhou, Y.C.; Qin, Q.W. Immunogenicity and protective effects of inactivated Singapore grouper iridovirus (SGIV) vaccines in orange-spotted grouper, Epinephelus coioides. Dev. Comp. Immunol. 2012, 38, 254–261. [Google Scholar] [CrossRef]
- Liu, W.Z.; Xu, J.; Ma, J.; LaPatra, S.E.; Meng, Y.; Fan, Y.; Zhou, Y.; Yang, X.; Zeng, L. Immunological responses and protection in Chinese giant salamander Andrias davidianus immunized with inactivated iridovirus. Vet. Microbiol. 2014, 174, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Yi, W.Y.; Zhang, X.; Zeng, K.; Xie, D.F.; Song, C.; Tam, K.; Liu, Z.J.; Zhou, T.H.; Li, W. Construction of a DNA vaccine and its protective effect on largemouth bass (Micropterus salmoides) challenged with largemouth bass virus (LMBV). Fish Shellfish. Immunol. 2020, 106, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.Y.; Zhang, C.S.; Yuan, X.M.; Huang, L.; Hu, D.Y.; Yu, Z.; Yin, W.L.; Lin, L.Y.; Pan, X.Y.; Yang, G.L.; et al. Oral vaccination with recombinant Pichia pastoris expressing iridovirus major capsid protein elicits protective immunity in largemouth bass (Micropterus salmoides). Front. Immunol. 2022, 13, 852300. [Google Scholar] [CrossRef]
- Zhang, M.J.; Chen, X.Y.; Xue, M.Y.; Jiang, N.; Li, Y.Q.; Fan, Y.D.; Zhang, P.; Liu, N.C.; Xiao, Z.D.; Zhang, Q.H.; et al. Oral vaccination of largemouth bass (Micropterus salmoides) against largemouth bass ranavirus (LMBV) using yeast surface display technology. Animals 2023, 13, 7. [Google Scholar] [CrossRef]
- Wang, Q.W.; Liang, X.; Ning, Y.S.; Liu, S.J.; Liang, Z.J.; Zhang, Z.M.; Chen, Y.J.; Cao, J.Q.; Wang, F.B.; Lan, L.F.; et al. Surface display of major capsid protein on Bacillus subtilis spores against largemouth bass virus (LMBV) for oral administration. Fish Shellfish. Immunol. 2023, 135, 108627. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 18, 493–497. [Google Scholar] [CrossRef]
- Jiang, N.; Fan, Y.D.; Zhou, Y.; Liu, W.Z.; Ma, J.; Meng, Y.; Xie, C.X.; Zeng, L.B. Characterization of Chinese giant salamander iridovirus tissue tropism and inflammatory response after infection. Dis. Aquat. Org. 2015, 114, 229–237. [Google Scholar] [CrossRef]
- Ho, P.Y.; Byadqi, O.; Wang, P.C.; Tsai, M.A.; Liaw, L.L.; Chen, S.C. Identification, molecular cloning of IL-1β and its expression profile during Nocardia seriolae infection in largemouth bass, Micropterus salmoides. Int. J. Mol. Sci. 2016, 17, 1670. [Google Scholar] [CrossRef]
- Amend, D.F. Potency testing of fish vaccines. International Symposium in Fish Biologics: Serodiagnostics and Vaccines. Dev. Biol. Stand. 1981, 49, 447–454. [Google Scholar]
- Ma, J.; Bruce, T.J.; Jones, E.M.; Cain, K.D. A review of fish vaccine development strategies: Conventional methods and modern biotechnological approaches. Microorganisms 2019, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Ma, J.; Fan, Y.D.; Zhou, Y.; Xu, J.; Liu, W.Z.; Gu, Z.M.; Zeng, L.B. Immune response and protection in gibel carp, Carassius gibelio, after vaccination with β-propiolactone inactivated cyprinid herpesvirus 2. Fish Shellfish. Immunol. 2016, 49, 344–350. [Google Scholar] [CrossRef]
- Luo, X.; Wang, R.X.; Ma, B.F.; Chang, O.Q.; Fu, X.Z.; Liang, H.R.; Niu, Y.J.; Lin, Q.; Liu, L.H.; Lv, A.; et al. Inactive LMBV vaccine with nano-aluminum adjuvant provided immune protection in largemouth bass (Micropterus salmoides). Aquaculture 2025, 595, 741634. [Google Scholar] [CrossRef]
- Shinmoto, H.; Taniguchi, K.; Ikawa, T.; Kawai, K.; Oshima, S. Phenotypic diversity of infectious red sea bream iridovirus isolates from cultured fish in Japan. Appl. Environ. Microbiol. 2009, 75, 3535–3541. [Google Scholar] [CrossRef]
- Cannon, M.S.; Mollenhauer, H.H.; Eurell, T.E.; Lewis, D.H.; Cannon, A.M.; Tompkins, C. An ultrastructural study of the leukocytes of the channel catfish, Ictalurus punctatus. J. Morphol. 1980, 164, 1–23. [Google Scholar] [CrossRef]
- Wang, X.D.; Rao, J.R. The discovery and studies of the extensive erythrocytic phagocytosis. J. Fish. China 1996, 20, 72–75. [Google Scholar]
- Burton, D.R. Antibodies, viruses and vaccines. Nat. Rev. Immunol. 2002, 2, 706–713. [Google Scholar] [CrossRef]
- Li, S.; Chen, J.N.; Zhao, L.N.; Huang, C.P.; Huang, J.L.; Wang, G.P.; Zhong, Y. Preparation of anti-largemouth bass ranavirus egg yolk antibody and establishment of indirect ELISA method. South China Fish. Sci. 2024, 20, 129–139. [Google Scholar] [CrossRef]
- Bernasconi, N.L.; Traggiai, E.; Lanzavecchia, A. Maintenance of serological memory by polyclonal activation of human memory B cells. Science 2002, 298, 2199–2202. [Google Scholar] [CrossRef]
- Ito, T.; Maeno, Y. Effect of booster shot and investigation of vaccination efficacy period against herpesviral haematopoietic necrosis (HVHN) in goldfish Carassius auratus. Vet. Microbiol. 2015, 175, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Tort, L.; Balasch, J.C.; Mackenzie, S. Fish immune system. A crossroads between innate and adaptive responses. Inmunología 2003, 22, 277–286. [Google Scholar]
- Bernatchez, L.; Landry, C. MHC studies in nonmodel vertebrates: What have we learned about natural selection in 15 years? J. Evol. Biol. 2003, 16, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Van den Hoorn, T.; Paul, P.; Jongsma, M.L.M.; Neefjes, J. Routes to manipulate MHC class II antigen presentation. Curr. Opin. Immunol. 2011, 23, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Fan, Y.D.; Zhou, Y.; Meng, Y.; Liu, W.Z.; Li, Y.Q.; Xue, M.Y.; Robert, J.; Zeng, L.B. The immune system and the antiviral responses in Chinese giant salamander, Andrias davidianus. Front. Immunol. 2021, 12, 718627. [Google Scholar] [CrossRef]
- Hercend, T.; Reinherz, E.L.; Meuer, S.; Schlossman, S.F.; Ritz, J. Phenotypic and functional heterogeneity of human cloned natural killer cell lines. Nature 1983, 301, 158–160. [Google Scholar] [CrossRef]
- Jia, Y.J.; Xia, J.Y.; Jiang, F.Y.; Li, Y.; Chen, G.; Zhu, B. Antigenic epitope screening and functional modification of mannose enhance the efficacy of largemouth bass virus subunit vaccines. J. Fish Dis. 2022, 45, 1635–1643. [Google Scholar] [CrossRef]
- Jaafar, R.M.; Al-Jubury, A.; Chettri, J.K.; Dalsgaard, I.; Kania, P.W.; Buchmann, K. Secondary immune response of rainbow trout following repeated immersion vaccination. J. Fish Dis. 2018, 41, 117–123. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′–3′) | Accession no. or Reference | Usage |
|---|---|---|---|
| il-1β | CCGTGCCAACAGTGTGAAGA | XM_038733429 | qRT-PCR |
| GGGTGCTGTGTCCACCTTGC | |||
| igM | CGACTACGATATGAACTGGG | XM_038698199 | qRT-PCR |
| GCTGTTGTCTCTGGAGATGG | |||
| mhc II | GGGATGGAGACCAGGCGATA | XM_038735684 XM_038696403 | qRT-PCR qRT-PCR |
| cd8α | CCCGCTTGACAGCACATCCT CCAAGTCAGTGCACATCTAC GGGCCCAGTATGATTGAAGG | ||
| β-actin | CCACCACAGCCGAGAGGGAA | [21] | qRT-PCR |
| TCATGGTGGATGGGGCCAGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, T.; Zhai, J.; Li, C.; Zeng, L.; Li, Y.; Liu, W.; Meng, Y.; Fan, Y.; Huang, Z.; Zhou, Y.; et al. Immunological Responses and Protection in the Largemouth Bass (Microterus salmoides) Immunized with Inactivated Vaccine Against Largemouth Bass Ranavirus (LMBRaV). Animals 2025, 15, 803. https://doi.org/10.3390/ani15060803
Yang T, Zhai J, Li C, Zeng L, Li Y, Liu W, Meng Y, Fan Y, Huang Z, Zhou Y, et al. Immunological Responses and Protection in the Largemouth Bass (Microterus salmoides) Immunized with Inactivated Vaccine Against Largemouth Bass Ranavirus (LMBRaV). Animals. 2025; 15(6):803. https://doi.org/10.3390/ani15060803
Chicago/Turabian StyleYang, Tao, Jiale Zhai, Chenyang Li, Lingbing Zeng, Yiqun Li, Wenzhi Liu, Yan Meng, Yuding Fan, Zhenyu Huang, Yong Zhou, and et al. 2025. "Immunological Responses and Protection in the Largemouth Bass (Microterus salmoides) Immunized with Inactivated Vaccine Against Largemouth Bass Ranavirus (LMBRaV)" Animals 15, no. 6: 803. https://doi.org/10.3390/ani15060803
APA StyleYang, T., Zhai, J., Li, C., Zeng, L., Li, Y., Liu, W., Meng, Y., Fan, Y., Huang, Z., Zhou, Y., & Jiang, N. (2025). Immunological Responses and Protection in the Largemouth Bass (Microterus salmoides) Immunized with Inactivated Vaccine Against Largemouth Bass Ranavirus (LMBRaV). Animals, 15(6), 803. https://doi.org/10.3390/ani15060803