
Papillary and Callous Scales in the Integument of Agamid Lizards (Agamidae, Sauria) as a Phenomenon of Extraordinary Development of the Corneous Layers

 , , ,
, , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Gross and Microscopic Investigation
2.3. Terminology
3. Results
3.1. Distribution of the Modified Scales
3.2. Morphology of the Unspecialized Scaled Epidermis
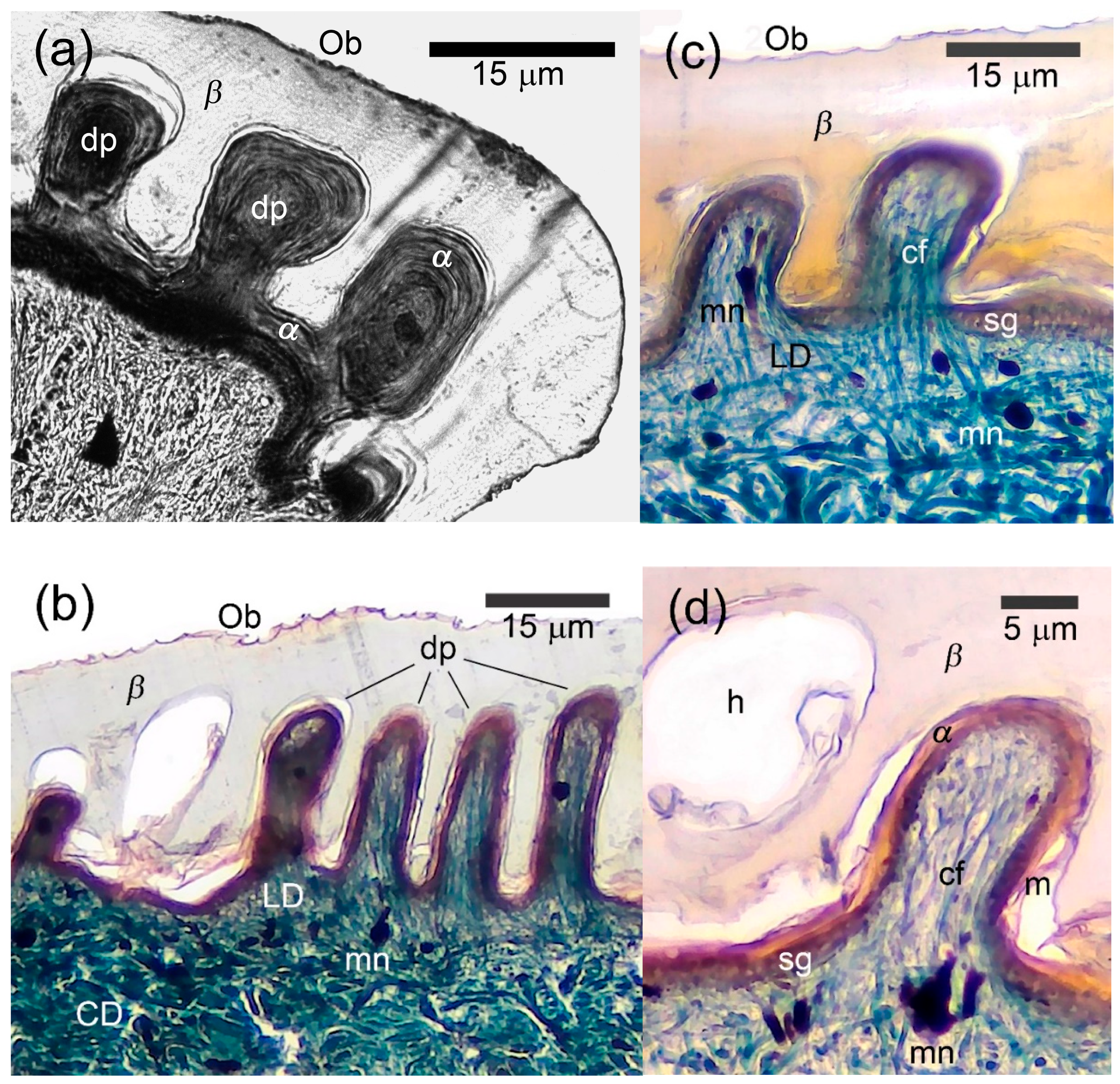
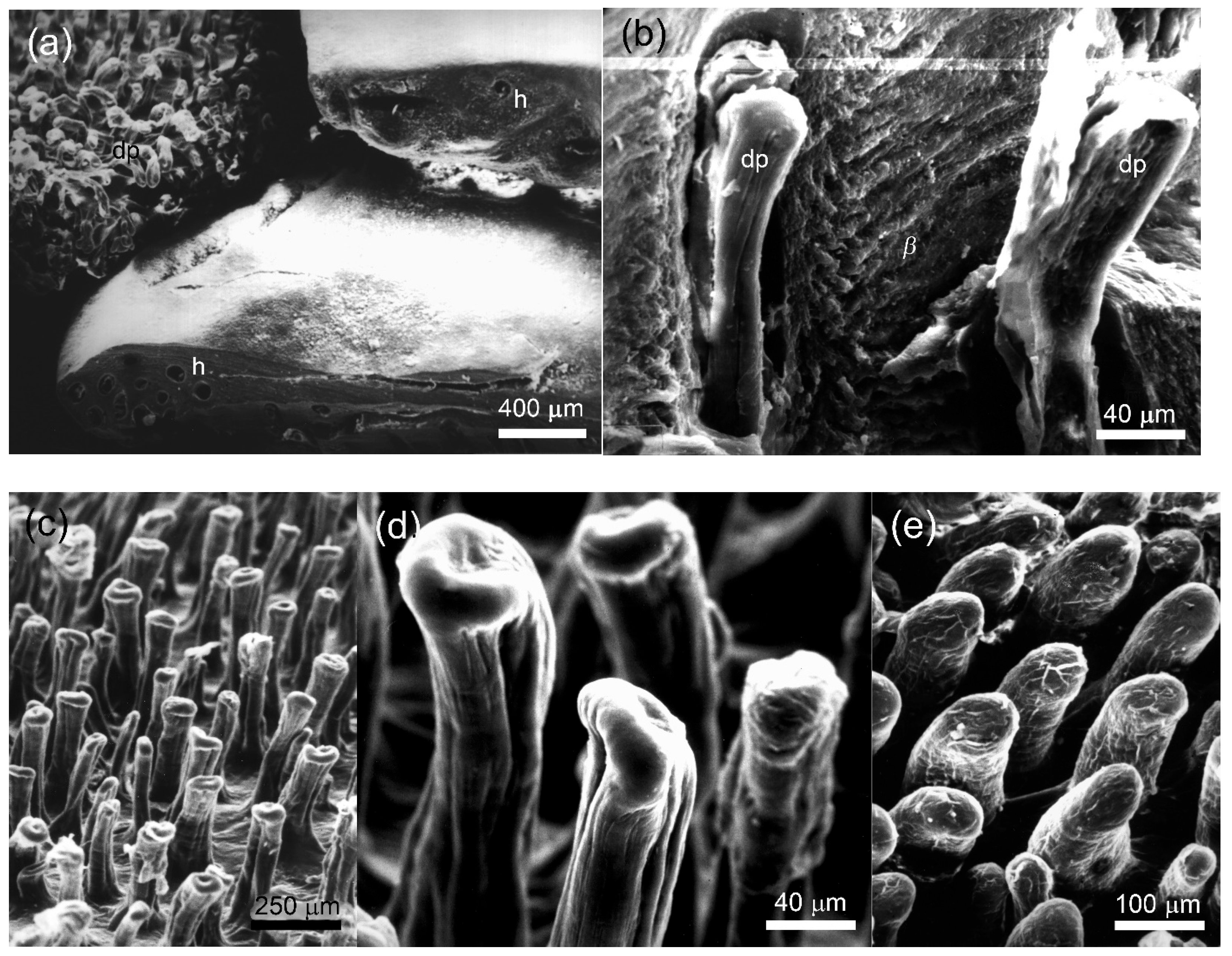
3.3. Morphology of the Papillary Scales
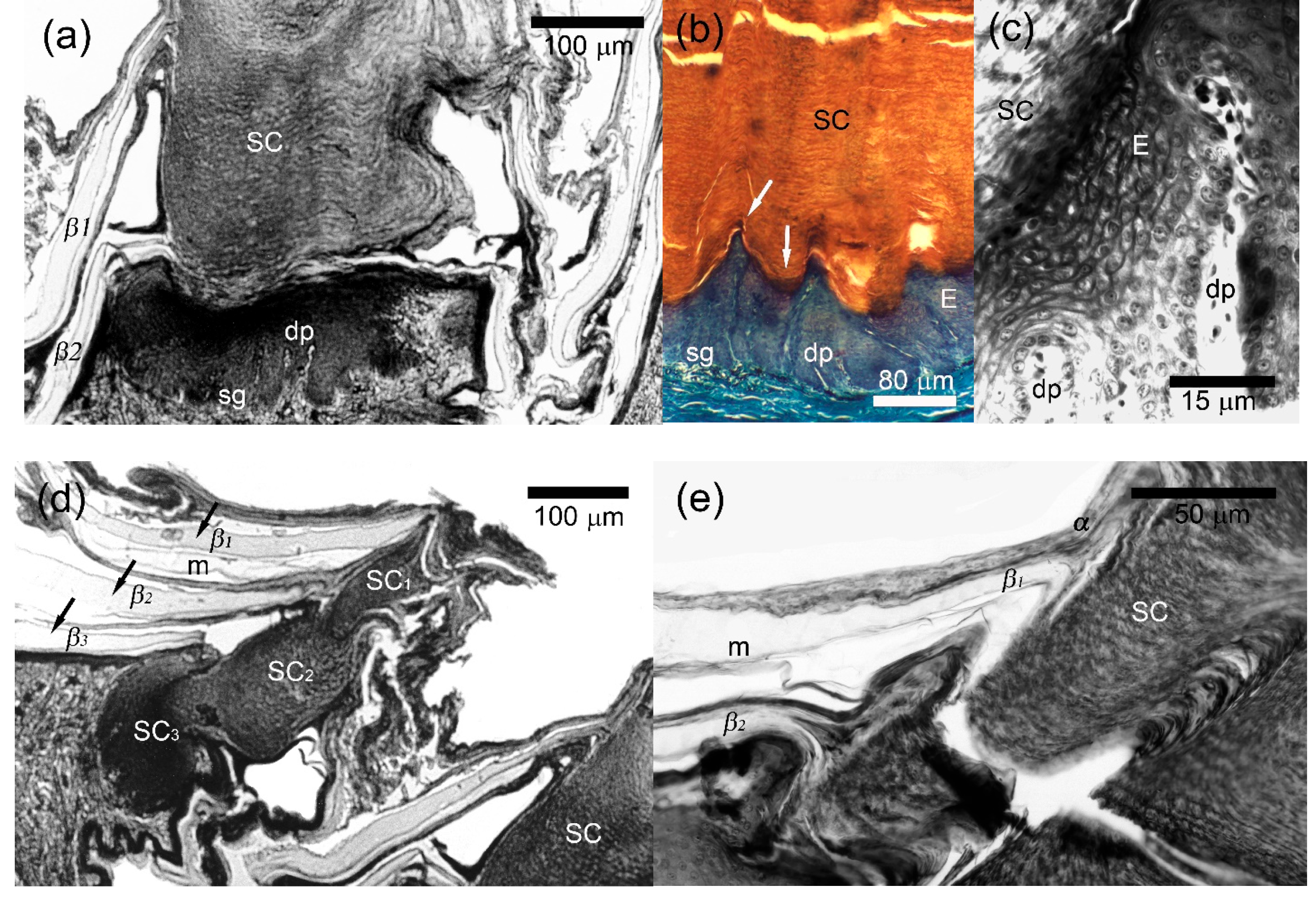
3.4. Morphology of the Callous Scales
4. Discussion
4.1. Papillary Scales and Their Functional Value
4.2. Callous Scales: Affinity to Epidermal Glands, Functional and Taxonomic Value
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Acanthocercus adramitanus (2): ZISP 32061, 32221.
- A. atricollis (7): ZFMK 2691, 2692, 7423, 7424, 18394, 29397; ZISP 19699.
- A. gregori (26): ZFMK 2685, 2689, 2690, 15865–15868, 19161, 20854, 21073–21076, 21078, 21079, 21081–21086, 41655, 61661, 61662, 61644, 61665.
- A. minutus (7): ZFMK 2683, 2684, 2686–2688, 16229, 16230.
- A. ugandaensis (7): ZFMK 55710–55716.
- A. yemensis (3): ZISP 32062, 32115, 32220.
- Laudakia nupta (58): ZFMK 2682, 7924, 8627, 8628, 20824–20826; GNM Museum numbers: Re. ex. 5521–5560; SMF 63217–63220, 63222, 63224, 63225, 63228–63230; ZISP 24661.
References
- Boulenger, G.A. Catalogue of the Lizards in the British Museum (Natural History), 2nd ed.; Taylor & Francis: London, UK, 1885; Volume I: Geckonidae, Eublepharidae, Uroplatidae, Pygopodidae, Agamidae. [Google Scholar]
- Baig, K.J.; Wagner, P.; Ananjeva, N.B.; Böhme, W. A morphology-based taxonomic revision of Laudakia Gray, 1845 (Squamata: Agamidae). Vertebr. Zool. 2012, 62, 213–260. [Google Scholar] [CrossRef]
- Cooper, W., Jr.; van Wyk, J.H.; Mouton, P.F.N.; Al-Johany, A.M.; Lemos-Espinal, J.A.; Paulissen, M.A.; Flowers, M. Lizard antipredatory behaviors preventing extraction from crevices. Herpetologica 2000, 56, 394–401. [Google Scholar]
- Ramm, T.; Roycroft, E.J.; Müller, J. Convergent evolution of tail spines in squamate reptiles driven by microhabitat use. Biol. Lett. 2020, 16, 20190848. [Google Scholar] [CrossRef]
- Iohanssen, L.K. On the Comparative Morphology of Epidermal Receptors of Agamid and Geckonid Lizards. Diploma Thesis, Leningrad State University, St-Petersburg, Russia, 1987. [Google Scholar]
- Ananjeva, N.B.; Peters, G.; Macey, J.B.; Papenfuss, T.J. Stellio sacra (Smith 1935)—A distinct species of Asiatic Rock agamid from Tibet. Asiat. Herpetol. Res. 1990, 3, 104–115. [Google Scholar]
- Ananjeva, N.B.; Iohanssen, L.K.; Duisebayeva, T.N. Skin receptors of Laudakia (Agamidae, Sauria) with some comments about scalation of Asian rock agamids. Russ. J. Herpetol. 2000, 7, 51–60. [Google Scholar]
- Smirnova, Y. Dermal papillous scales at the rock agamas of the genus Laudakia (Sauria, Agamidae). Curr. Res. Herpetol. 2003, 2, 124–136, (In Russian with English summary). [Google Scholar]
- Tölg, F. Beiträge zur Kenntnis drüsenartiger Epidermoidalorgane der Eidechsen. Arb. Aus Dem Zool. Inst. Der Univ. Wien 1905, 15, 119–154. [Google Scholar]
- Gabe, M.; Saint-Girons, H. Contribution à la morphologie comparée du cloaque et des glandes épidermoïdes de la région cloacale chez les Lépidosauriens. Mém. Mus. Hist. Nat. Série A Zool. (1950–1992) 1965, 33, 149–292. [Google Scholar]
- Smith, M.A. The Fauna of British India: Reptilia and Amphibia. II Sauria; Taylor and Francis: London, UK, 1935. [Google Scholar]
- Baig, K.J. A new species of Agama (Sauria: Agamidae) from Northern Pakistan. Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist. 1989, 9, 117–122. [Google Scholar]
- Baig, K.J.; Böhme, W. Callous scalation in female agamid lizards (Stellio-group of Agama) and its functional implications. Bonn. Zool. Beitr. 1991, 42, 275–281. [Google Scholar]
- Dujsebayeva, T.N. The histology of callous scales of the males of asian rock agamas, Laudakia caucasia and Laudakia himalayana (Reptilia: Agamidae). Russ. J. Herpetol. 1998, 5, 160–164. [Google Scholar]
- Dujsebayeva, T.N.; Ananjeva, N.B.; Miroschnichenko, L.V. Studies on specialized epidermal derivatives in iguianian lizards. I. Gross morphology, topography and histology of callose scales in the Asian Rock Agama, Laudakia himalayana (Steindachner, 1869) (Squamata: Agamidae). Amphibia-Reptilia 2007, 28, 537–546. [Google Scholar] [CrossRef]
- Harris, V.A. The Anatomy of the Rainbow Lizard; Hutchinson & Co.: London, UK, 1963. [Google Scholar]
- Moody, S. Phylogenetic and Historical Biogeographical Relationships of the Genera in the Family Agamidae (Reptilia: Lacertilia). Ph.D. Thesis, University of Michigan, Ann Arbor, MI, USA, 1980. [Google Scholar]
- Maderson, P.F.A.; Chiu, K.W. Epidermal glands in gekkonid lizards: Evolution and phylogeny. Herpetologica 1970, 26, 233–238. [Google Scholar]
- Maderson, P.F.A.; Chiu, K.W. Endocrine Relationships of Secondary Sexual Derivatives in Tetrapods. In Current Trends in Comparative Endocrinology; Lofts, B., Holmes, W.N., Eds.; Hong Kong University Press: Hong Kong, China, 1985; pp. 1191–1194. [Google Scholar]
- Searby, C.A. Histological Description of Generation Glands and Their Functional Relationship to the Shedding Cycle in Cordylid Lizards. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2002. [Google Scholar]
- Ribeiro, L.B.; Kolodiuk, M.F.; Freire, E.M.X. Ventral colored patches in Tropidurus semitaeniatus (Squamata, Tropiduridae): Sexual dimorphism and association with reproductive cycle. J. Herpetol. 2010, 44, 177–182. [Google Scholar] [CrossRef]
- Mayerl, C.; Baeckens, S.; van Damme, R. Evolution and role of the follicular epidermal gland system in non-ophidian squamates. Amphibia-Reptilia 2015, 36, 185–206. [Google Scholar] [CrossRef]
- Panov, E.N.; Zykova, L.U. Eurasian Rock Agamas; KMK Press: Moscow, Russia, 2016. [Google Scholar]
- Dujsebayeva, T.N.; Ananjeva, N.; Böhme, W.; Wagner, P. Studies on specialized epidermal derivatives in iguanian lizards: II. New data on the scalation of the Malagasy iguanas of the genus Oplurus (Sauria: Iguanidae). Amphibia-Reptilia 2009, 30, 89–97. [Google Scholar] [CrossRef]
- Dujsebayeva, T.N.; Kalyabina, S.A.; Ananjeva, N.B.; Rastegar-Pouyani, N. Papillous scales of the skin of Laudakia nupta and Acanthocercus atricollis (Sauria, Agamidae). In Proceedings of the Fifth International Congress of Vertebrate Morphology, Bristol, UK, 12–17 July 1997; p. 74. [Google Scholar]
- Romeis, B. Mikroskopische Technik, 16th ed.; R. Oldenburg Verlag: München, Germany, Wien, Austria, 1968. [Google Scholar]
- Martoja, R.; Martoja-Pierson, U. Initiation aux Techniques de L’histologie Animali; Masson: Paris, France, 1967. [Google Scholar]
- Irish, F.J.; Williams, E.E.; Seling, E. Scanning electron microscopy of changes in epidermal structure occurring during the shedding cycle in squamate reptiles. J. Morphol. 1988, 197, 105–126. [Google Scholar] [CrossRef]
- Uetz, P.; Freed, P.; Aguilar, R.; Hošek, J. (Eds.) The Reptile Database. 2022. Available online: http://www.reptile-database.org/ (accessed on 30 December 2024).
- Wagner, P.; Butler, B.O.; Ceríaco, L.M.P.; Bauer, A.M. A new species of the Acanthocercus atricollis complex (Squamata: Agamidae). Salamandra 2021, 57, 449–463. [Google Scholar]
- Maderson, P.F.A. The structure and evolution of holocrine epidermal glands in sphaerodactyline and eublepharine gekkonid lizards. Copeia 1972, 3, 559–571. [Google Scholar] [CrossRef]
- Maderson, P.F.A. Some developmental problems of the Reptilian integument. In Biology of the Reptilia, Volume 14, Development A; Gans, C., Billett, F., Maderson, P.F.A., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 1985; pp. 571–598. [Google Scholar]
- IUCN. 2024 The IUCN Red List of Threatened Species. Version 2024-2. Available online: https://www.iucnredlist.org (accessed on 14 February 2025).
- Maderson, P.F.A.; Mayhew, W.W.; Sprague, G. Observations on the epidermis of desert-living iguanids. J. Morphol. 1970, 130, 25–35. [Google Scholar] [CrossRef]
- Montagna, W. Cutaneous Comparative Biology. Arch. Dermatol. 1971, 104, 577–591. [Google Scholar] [CrossRef]
- Sokolov, V.E. Integument of Mammalia; Nauka: Moscow, Russia, 1973. (In Russian) [Google Scholar]
- Lavker, R.M.; Sun, T.-T. Heterogeneity in epidermal basal keratinocytes: Morphological and functional correlation. Science 1982, 215, 1239–1241. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, P.W.; Davies, C.; Lucey, A.D. Hydrodynamics and compliant walls: Does the dolphin have a secret? Curr. Sci. 2000, 79, 758–765. [Google Scholar]
- Alibardi, L. The process of cornification evolved from the initial keratinization in the epidermis and epidermal derivatives of vertebrates: A new synthesis and the case of sauropsids. Int. Rev. Cell Mol. Biol. 2016, 327, 263–319. [Google Scholar] [PubMed]
- Alibardi, L. Vertebrate keratinization evolved into cornification mainly due to transglutaminase and sulfhydryl oxidase activities on epidermal proteins: An immunohistochemical survey. Anat. Rec. 2021, 305, 333–358. [Google Scholar] [CrossRef]
- Alibardi, L. Immunocytochemical analysis of the process of keratinization of the epidermas of snakes. J. Zool. 2002, 258, 541–552. [Google Scholar] [CrossRef]
- Alibardi, L.; DeNardo, D.F. Ultrastructural and immunocytochemical features of the epidermis of the lizard Heloderma suspectum indicate richness in lipids and lack of a specialized shedding complex. Acta Zool. Stockh. 2013, 94, 35–43. [Google Scholar] [CrossRef]
- Ananjeva, N.B.; Tuniev, B.S. Some aspects of historical biogeography of Asian rock agamids. Russ. J. Herpetol. 1994, 1, 3–41. [Google Scholar]
- Khisroon, M.; Farooq, J.; Masroor, R. Systematics, Ecology and Distribution of Caucasian Rock Agama, Paralaudakia caucasia in District Chitral, Khyber Pakhtunkhwa Province, Pakistan. Putaj Sci. 2013, 19, 107–126. [Google Scholar]
- Mazanayeva, L.; Ananjeva, N. New data on habitats and distribution of the Caucasian agama, Paralaudakia caucasia (Eichwald, 1831) in Dagestan. Russ. J. Herpetol. 2016, 23, 293–304. [Google Scholar]
- Avilova, K.V. Spatial organization of the epithelial structures in the bill tip organ of Waterfowl (Anseriformes, Aves). Biol. Bull. Rev. 2018, 8, 234–244. [Google Scholar] [CrossRef]
- Fedorenko, A.G.; Avilova, K.V.; Lebedeva, N.V. The fine structure of the bill tip organ in waterfowl (Anseriformes, Aves) with the Mallard (Anas plathrhynchos) as an example. In Evolutionary and Functional Morphology of Vertebrates; Conference and Workshop dedicated to Felix Yanovich Dzerzhinsky; KMK Press: Moscow, Russia, 2017; pp. 283–288. [Google Scholar]
- Montagna, W. The Evolution of Human Skin. J. Hum. Evol. 1985, 14, 3–22. [Google Scholar] [CrossRef]
- Wu, P.; Hou, L.; Plikus, M.; Hughes, M.; Scehnet, J.; Suksaweang, S.; Widelitz, R.; Jiang, T.; Chuong, C.M. Evo-Devo of amniote integuments and appendages. Int. J. Dev. Biol. 2004, 48, 249–270. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, P.W. Hydrodynamics of dolphin skin and other compliant surfaces. WIT Trans. State Art Sci. Eng. 2006, 4, 447–456. [Google Scholar]
- Dujsebayeva, T.N. The skin development in the Pallas’ Coluber, Elaphe dione (Pallas, 1773) (Serpentes, Colubridae). Russ. J. Herpetol. 2008, 15, 44–54. [Google Scholar]
- Di-Poï, N.; Milinkovitch, M.C. The anatomical placode in reptile scale morphogenesis indicates shared ancestry among skin appendages in amniotes. Sci. Adv. 2016, 2, e1600708. [Google Scholar] [CrossRef]
- Alibardi, L.; Thompson, M.B. Epidermal differentiation in the developing scales of embryos of the Australian scincid lizard Lampropholis guichenoti. J. Morphol. 1999, 241, 139–152. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species, 6th ed.; P.F. Collier & Son: New York, NY, USA, 1872. [Google Scholar]
- Maderson, P.F.A. Lizard glands and lizard hands: Models for evolutionary study. Forma Funct. 1970, 3, 179–204. [Google Scholar]
- Maderson, P.F.A. The tetrapod epidermis: A system protoadapted as a semiochemical source. In Chemical Signals in Vertebrates 4: Ecology, Evolution, and Comparative Biology; Duvall, D., Muller-Schwarze, D., Silverstein, R.M., Eds.; Plenum Press: New York, NY, USA, 1986; pp. 13–25. [Google Scholar]
- van Wyk, J.H.; Maderson, P.F.A. Epidermal glands and the shedding cycle in the cordylid lizard. Cordylus giganteus (Sauria: Cordylidae). In Proceedings of the American Society of Ichthyologists and Herpetologists Meeting, Edmonton, AB, Canada, 15–19 June 1995; pp. 195–196. [Google Scholar]
- Louw, S.; Burger, B.V.; Le Roux, M.; van Wyk, J.H. Lizard Epidermal Gland Secretions. II. Chemical Characterization of the Generation Gland Secretion of the Sungazer, Cordylus giganteus. J. Nat. Prod. 2011, 74, 1364–1369. [Google Scholar]
- le Mouton, P.F.N.; Flemming, A.F.; Broeckhoven, C. Generation Gland Morphology in Cordylid Lizards: An Evolutionary Perspective. J. Morphol. 2014, 275, 456–464. [Google Scholar] [CrossRef]
- Cole, C.J. Femoral glands of the lizard, Crotaphytus collaris. J. Morphol. 1966, 118, 119–135. [Google Scholar] [CrossRef] [PubMed]
- De-Lima, A.K.S.; Domingos, F.M.C.B.; Chaves, S.B.; Pic-Taylor, A.; Sebben, A.; Klaczko, J. A new scent organ for Gymnodactylus lizards (Squamata: Phyllodactylidae) and an updated evolutionary scenario for the origin of squamate epidermal glands. Biol. J. Linn. Soc. 2018, 125, 561–575. [Google Scholar] [CrossRef]
- Anderson, S.C. The Lizards of Iran. In Contribution to Herpetology; Adler, C., Ed.; SSAR: Ithaca, NY, USA, 1999; Volume 15. [Google Scholar]
- Schatti, B.; Gasperetti, J. A Contribution to the Herpetofauna of Southwest Arabia. Fauna Saudi Arab. 1994, 14, 348–423. [Google Scholar]
- de Villiers, A.; le Mouton, P.F.N.; Flemming, A. Generation glands of cordylid lizards: Mechanism of secretion transfer to the environment. Amphibia-Reptilia 2015, 36, 351–360. [Google Scholar] [CrossRef]
- van Wyk, J.H.; Maderson, P.F.A. The histology of pre-cloacal epidermal glands in the rock lizard, Agama atra (Sauria: Agamidae). In Proceedings of the Abstracts of the Third Asian Herpetological Meeting, Almaty, Kazakhstan, 1–5 September 1998; pp. 42–43. [Google Scholar]
- van Wyk, J.H.; le Mouton, P.F.N. Glandular epidermal structures in cordylid lizards. Amphibia-Reptilia 1992, 13, 1–12. [Google Scholar] [CrossRef]
- Taylor, E.H.; Leonard, A.B. Concerning the relationship of certain Neotropical gekkonid lizard genera, with comments on the microscopical structure of their glandular scales. Univ. Kans. Sci. Bull. 1956, 38, 1019–1029. [Google Scholar]
- Kluge, A.G. Higher taxonomic categories of gekkonid lizards and their evolution. Bull. Am. Mus. Nat. Hist. 1967, 135, 1–59. [Google Scholar]
- Kluge, A.G. Epidermal gland evolution in gekkonoid lizards. J. Herpetol. 1983, 17, 89–90. [Google Scholar] [CrossRef]
- Maderson, P.F.A. The histology of the escutcheon scales of Gonatodes (Gekkonidae) with a comment on a squamate sloughing cycle. Copeia 1967, 4, 743–752. [Google Scholar] [CrossRef]
- Maderson, P.F.A.; Chiu, K.W. The effects of androgens on the β-glands of the Tokay (Gekko gecko): Modification of an hypothesis. J. Morphol. 1981, 167, 109–118. [Google Scholar] [CrossRef]
- Underwood, G. On the lizards of the family Pygopodidae. A contribution to the morphology and phylogeny of the Squamata. J. Morphol. 1957, 100, 207–268. [Google Scholar] [CrossRef]
- Bauer, A.M.; Russell, A.P. Hoplodactylus delcourti n. sp. (Reptilia: Gekkonidae), the largest known gecko. N. Z. J. Zool. 1986, 13, 141–148. [Google Scholar] [CrossRef]
- Ananjeva, N.B.; Dujsebayeva, T.N. Femoral pores of agamid lizards (Agamidae, Sauria, Reptilia). Curr. Res. Herpetol. 2007, 7, 3–15, (In Russian with English summary). [Google Scholar]
- Athavale, N.V.; Asnani, M.V.; Pilo, B.; Shan, R.V. Histo-morphology of the femoral glands in the agamid lizard, Uromastix hardwickii (Gray). J. Anim. Morphol. Physiol. 1977, 24, 51–55. [Google Scholar]
- Dujsebayeva, T.N.; Miroshnichenko, L.V.; Nigay, N. Histological description of the callous epidermis of Uromastyx lizards (Squamata; Agamidae). In Proceedings of the Abstracts of the Third Asian Herpetological Meeting, Almaty, Kazakhstan, 1–5 September 1998; p. 56. [Google Scholar]
- Walker, J.M. Accessory femoral pores in a colony of the Collared Lizard, Crotaphytus collaris in Texas. J. Herpetol. 1980, 14, 417–418. [Google Scholar] [CrossRef]
- Cole, C.J. Femoral glands in lizards: A review. Herpetologica 1966, 22, 199–206. [Google Scholar]
- Khannoon, E.R.R.; Dollahon, N.R.; Bauer, A.M. Comparative study of the pheromone-manufacturing femoral glands in two sympatric species of lacertid lizards (Acanthodactylus). Zool. Sci. 2013, 30, 110–117. [Google Scholar] [CrossRef]
- Antoniazzi, M.M.; Jared, C.; Pellegrini, C.M.R.; Machan, N. Epidermal glands in Squamata: Morphology and histochemistry of the pre-cloacal glands in Amphisbaena alba (Amphisbaenia). Zoomorphology 1993, 113, 199–203. [Google Scholar] [CrossRef]
- Vences, M.; Müller-Jung, J.; Glaw, F.; Böhme, W. Review of the Zonosaurus aeneus species group, with resurrection of Zonosaurus subunicolor (Boettger 1881). Senckenb. Biol. 1996, 76, 47–59. [Google Scholar]
- Carvalho, A.L.G.; Jeckel, A.M.; Nisa, C.; Luna, M.C.; Piantoni, C. A novel epidermal gland type in lizards (α-gland): Structural organization, histochemistry, protein profile and phylogenetic origins. Zool. J. Linn. Soc. 2021, 192, 1137–1166. [Google Scholar] [CrossRef]
- Zheng, Y.; Wiens, J.J. Combining phylogenomic and supermatrix approaches, and a time-calibrated phylogeny for squamate reptiles (lizards and snakes) based on 52 genes and 4162 species. Mol. Phylogenet. Evol. 2016, 94, 537–547. [Google Scholar] [CrossRef]
- Dilmukhamedov, M.E. On the secretory function of the reptilian integument. In The Problems of Herpetology; Nauka: Leningrad, Russia, 1985; p. 71. (In Russian) [Google Scholar]
- Brady, S.P. Arakeratosis. JAAD 2004, 50, 77–84. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Number of Specimens Studied in Parentheses | Sex/Age | N | Callous Scales | Papillary Scales | Habitats | |||
|---|---|---|---|---|---|---|---|---|
| PC | MV | P/P | C | |||||
| Rows | Scale/Row | +/– | +/– | +/– | ||||
| Acanthocercus adramitanus (2) | M | 1 | 1 | 8 | – | + | + | Rocky areas |
| F | 1 * | 1 | 10 | – | ˂ | ˂ | ||
| A. atricollis (7) | M | 5 | 2–3 | 3–6 | – | + | + | Savanna, woodland, forest clearings |
| F | 1 | – | – | – | – | – | ||
| subad M? | 1 | 1 | ? | – | ˂ | ˂ | ||
| A. gregori (26) | M | 13 | 2–3 | 2–6 | – | + | + | Grassland, savanna |
| F | 6 | – | – | – | ˂ | ˂ | ||
| subad M? | 4 | ? | ? | – | – | – | ||
| subad F? | 2 | – | – | – | – | – | ||
| juv | 1 | ? ** | – | – | – | – | ||
| A. minutus (7) | M | 2 | 3–4 | 1–6 | – | + | + | Savanna |
| F | 2 | – | – | – | – | – | ||
| juv | 2 | ? ** | – | – | – | – | ||
| juv | 1 | – | – | – | – | – | ||
| A. ugandaensis (7) | M | 3 | 2–3 | 4–7 | – | + | + | Forest, artificial |
| F | 1 | – | – | – | – | – | ||
| subad M? | 2 | 2 | 4–6 | – | – | – | ||
| subad F? | 1 | – | – | – | – | – | ||
| A. yemensis (3) | M | 1 | 2 | 8, 10 | – | + | + | Rocky areas |
| F ** | 2 | 2 | 8 | – | ˂ | ˂ | ||
| Laudakia nupta (58) | M | 33 | 1–7 | 8–10 | + | + | + | Rocky areas, shrubland |
| F | 12 | – | – | – | – | – | ||
| subad M? | 3 | + | + | + | + | + | ||
| subad F? | 7 | – | – | – | – | – | ||
| juv | 3 | – | – | – | – | – | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dujsebayeva, T.N.; Ananjeva, N.B.; Rastegar-Pouyani, N.; Al-Johany, A.M.; Melnikov, D.A. Papillary and Callous Scales in the Integument of Agamid Lizards (Agamidae, Sauria) as a Phenomenon of Extraordinary Development of the Corneous Layers. Animals 2025, 15, 743. https://doi.org/10.3390/ani15050743
Dujsebayeva TN, Ananjeva NB, Rastegar-Pouyani N, Al-Johany AM, Melnikov DA. Papillary and Callous Scales in the Integument of Agamid Lizards (Agamidae, Sauria) as a Phenomenon of Extraordinary Development of the Corneous Layers. Animals. 2025; 15(5):743. https://doi.org/10.3390/ani15050743
Chicago/Turabian StyleDujsebayeva, Tatjana N., Natalia B. Ananjeva, Nasrullah Rastegar-Pouyani, Awadh M. Al-Johany, and Daniel A. Melnikov. 2025. "Papillary and Callous Scales in the Integument of Agamid Lizards (Agamidae, Sauria) as a Phenomenon of Extraordinary Development of the Corneous Layers" Animals 15, no. 5: 743. https://doi.org/10.3390/ani15050743
APA StyleDujsebayeva, T. N., Ananjeva, N. B., Rastegar-Pouyani, N., Al-Johany, A. M., & Melnikov, D. A. (2025). Papillary and Callous Scales in the Integument of Agamid Lizards (Agamidae, Sauria) as a Phenomenon of Extraordinary Development of the Corneous Layers. Animals, 15(5), 743. https://doi.org/10.3390/ani15050743