
Missense Mutations in FDNC5 Associated with Morphometric Traits and Meat Quality in Hainan Black Goats
 , , , , and
, , , , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Total RNA and DNA Extraction
2.3. Primer Design
2.4. Statistical Analysis
3. Results
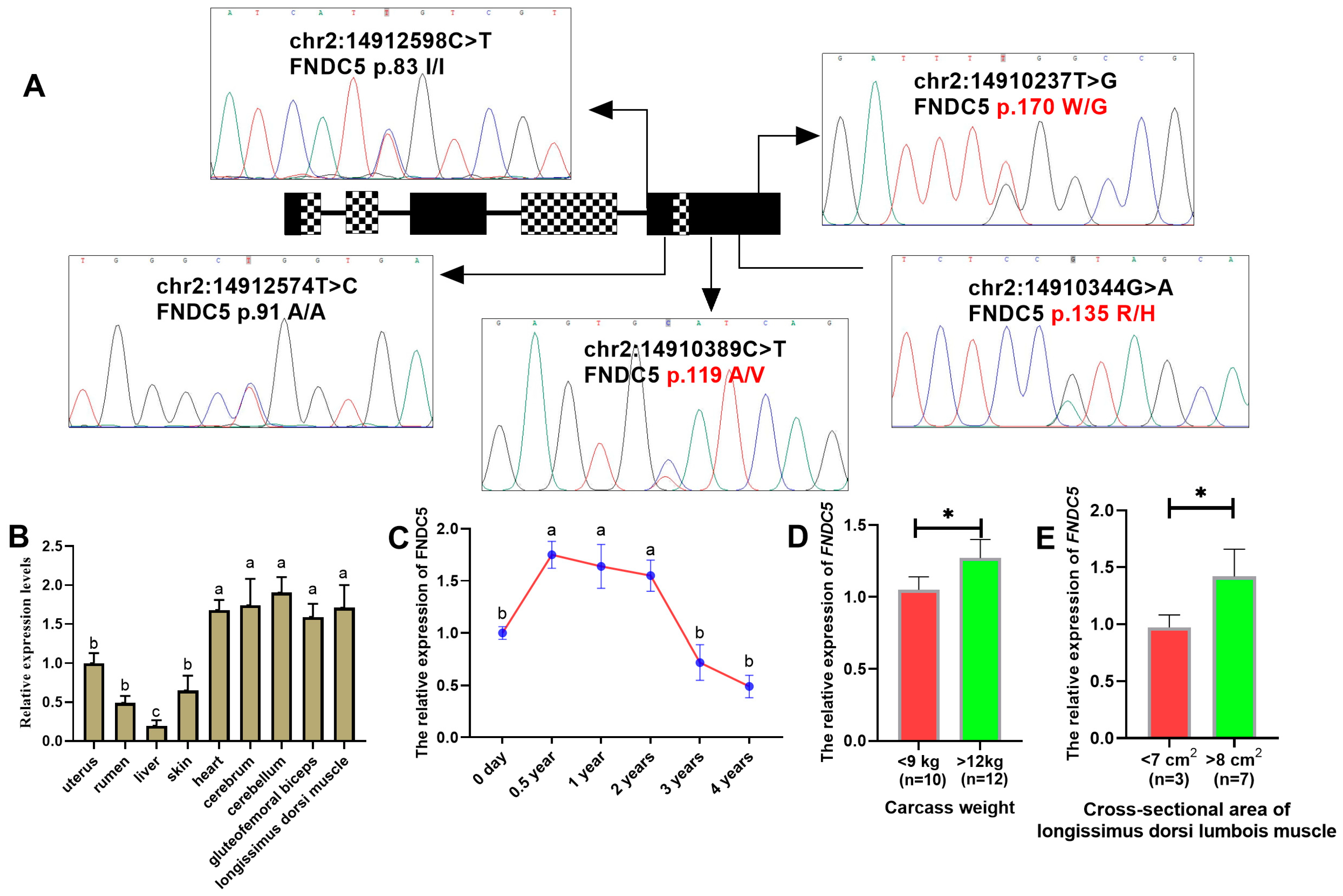
3.1. Identification of Missense Mutations in the Goat FNDC5 Gene
3.2. The Impact of Missense Mutations on the Structure of the FNDC5 Protein
3.3. The mRNA Expression of FNDC5 in HNBGs
3.4. Association Analysis Between the FNDC5 Missense Mutations and Traits
3.5. Missense Mutations Haplotype and Combination Genotype Analysis of the FNDC5 Gene
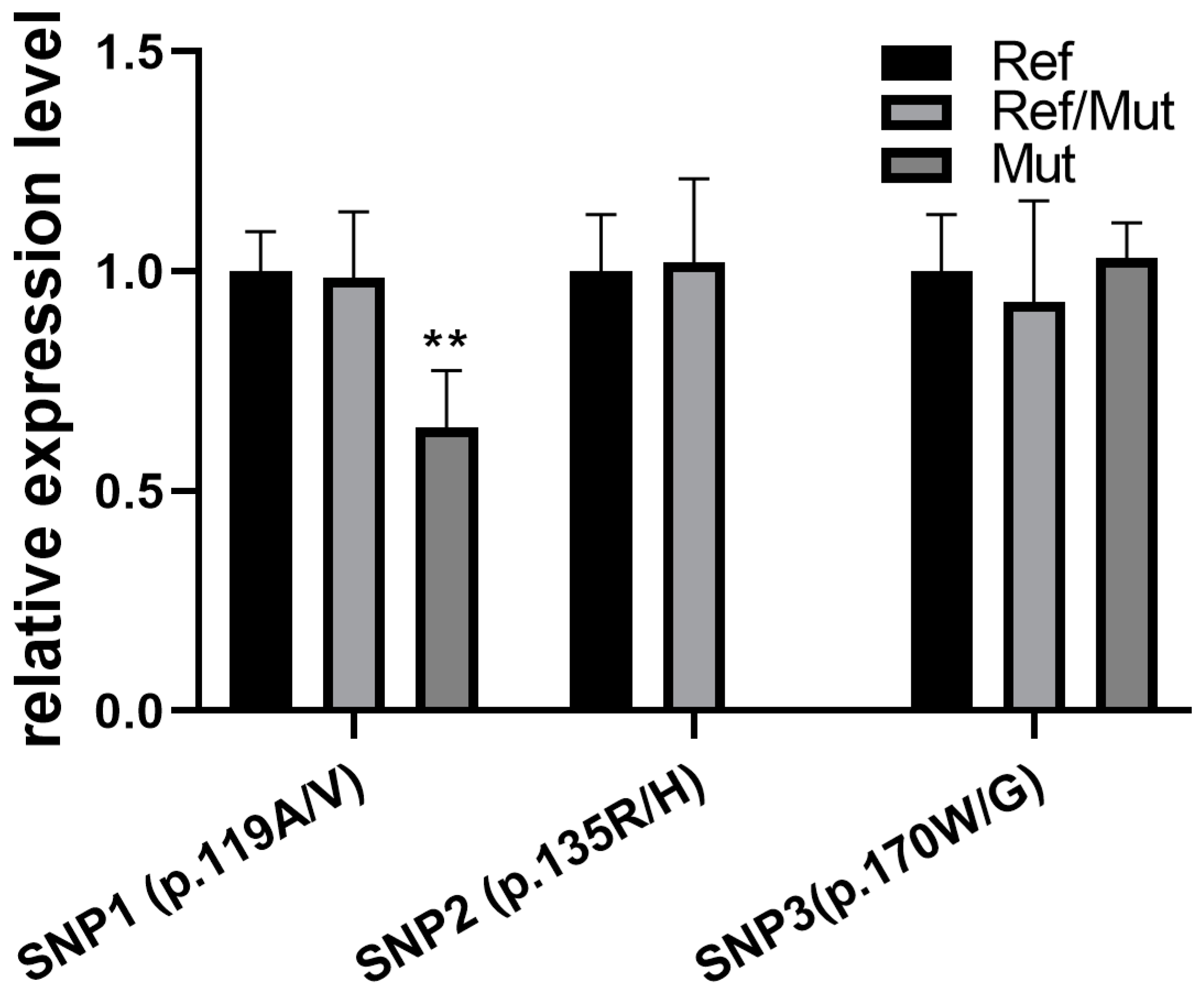
3.6. Missense Mutations Affect the Expression of the FNDC5 Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haile, A.; Gizaw, S.; Getachew, T.; Mueller, J.P.; Amer, P.; Rekik, M.; Rischkowsky, B. Community-based breeding programmes are a viable solution for Ethiopian small ruminant genetic improvement but require public and private investments. J. Anim. Breed. Genet. 2019, 136, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.; Almeida, S.; Pereira, E.; Mangachaia, F.; Rodrigues, S. Physicochemical characteristics of sheep and goat pâtés. differences between fat sources and proportions. Heliyon 2019, 5, e02119. [Google Scholar] [CrossRef]
- Banskalieva, V.V.; Sahlu, T.; Goetsch, A.L. Fatty acid composition of goat muscles and fat depots: A review. Small Rumin. Res. 2000, 37, 255–268. [Google Scholar] [CrossRef]
- Wu, Q.; Han, X.; Zhang, Y.; Liu, H.; Zhou, H.; Wang, K.; Han, J. One Copy Number Variation within the Angiopoietin-1 Gene Is Associated with Leizhou Black Goat Meat Quality. Animals 2024, 14, 2682. [Google Scholar] [CrossRef]
- Feng, H.; Shi, H.; Yang, F.; Yun, Y.; Wang, X. Impact of anthocyanins derived from Dioscorea alata L. on growth performance, carcass characteristics, antioxidant capacity, and immune function of Hainan black goats. Front. Vet. Sci. 2023, 10, 1283947. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Chai, Y.; Zhang, W.; Cheng, Y.; Zhang, Z.; An, Q.; Chen, S.; Man, C.; Du, L.; Zhang, W.; et al. Whole-Genome Sequencing Reveals the Genomic Characteristics and Selection Signatures of Hainan Black Goat. Genes 2022, 13, 1539. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Peng, W.; Mao, K.; Yang, Y.; Wu, Q.; Wang, K.; Zeng, M.; Han, X.; Han, J.; Zhou, H. The Changes in Fecal Bacterial Communities in Goats Offered Rumen-Protected Fat. Microorganisms 2024, 12, 822. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Feng, G.; Guo, C.; Li, Z.; Liu, D.; Liu, G.; Zou, X.; Sun, B.; Guo, Y.; Deng, M.; et al. Identification of functional circRNAs regulating ovarian follicle development in goats. BMC Genom. 2024, 25, 893. [Google Scholar] [CrossRef] [PubMed]
- Maak, S.; Norheim, F.; Drevon, C.A.; Erickson, H.P. Progress and Challenges in the Biology of FNDC5 and Irisin. Endocr. Rev. 2021, 42, 436–456. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Cui, F.; Ning, K.; Wang, Z.; Fu, P.; Wang, D.; Xu, H. Role of irisin in physiology and pathology. Front. Endocrinol. 2022, 13, 962968. [Google Scholar] [CrossRef] [PubMed]
- Waseem, R.; Shamsi, A.; Mohammad, T.; Hassan, M.I.; Kazim, S.N.; Chaudhary, A.A.; Rudayni, H.A.; Al-Zharani, M.; Ahmad, F.; Islam, A. FNDC5/Irisin: Physiology and Pathophysiology. Molecules 2022, 27, 1118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, K.; Jin, Y.; Liu, J.; Wang, Y.; Xue, Y.; Liu, H.; Chen, Q.; Cao, Z.; Jia, X.; et al. Explore novel molecular mechanisms of FNDC5 in ischemia-reperfusion (I/R) injury by analyzing transcriptome changes in mouse model of skeletal muscle I/R injury with FNDC5 knockout. Cell Signal. 2024, 113, 110959. [Google Scholar] [CrossRef]
- Shimonty, A.; Pin, F.; Prideaux, M.; Peng, G.; Huot, J.; Kim, H.; Rosen, C.J.; Spiegelman, B.M.; Bonewald, L.F. Deletion of FNDC5/irisin modifies murine osteocyte function in a sex-specific manner. eLife 2024, 12, RP92263. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Yao, J.; Li, J.; Zhang, J.; Wang, D.; Zuo, H.; Zhang, Y.; Xu, B.; Zhong, Y.; Shen, F.; et al. Irisin ameliorates age-associated sarcopenia and metabolic dysfunction. J. Cachexia Sarcopenia Muscle 2023, 14, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Men, X.M.; Xu, Z.W.; Tao, X.; Deng, B.; Qi, K.K. FNDC5 expression closely correlates with muscle fiber types in porcine longissimus dorsi muscle and regulates myosin heavy chains (MyHCs) mRNA expression in C2C12 cells. PeerJ 2021, 9, e11065. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Wang, X.; Zhang, Q.; He, Y.; Zhang, X.; Yang, L.; Shi, J. Comparative Transcriptome Analysis Identifying the Different Molecular Genetic Markers Related to Production Performance and Meat Quality in Longissimus dorsi Tissues of MG × STH and STH Sheep. Genes 2020, 11, 183. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Xu, M.; Han, X.; Liu, H.; Han, J.; Sun, W.; Zhou, H. Transcriptome analysis of muscle atrophy in Leizhou black goats: Identification of key genes and insights into limb-girdle muscular dystrophy. BMC Genom. 2025, 26, 80. [Google Scholar] [CrossRef]
- Xu, T.; Xu, F.; Gu, L.; Rong, G.; Li, M.; Qiao, F.; Shi, L.; Wang, D.; Xia, W.; Xun, W.; et al. Landscape of alternative splicing in Capra_hircus. Sci. Rep. 2018, 8, 15128. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, Y.; Han, X.; Wu, Q.; Liu, H.; Han, J.; Zhou, H. Effects of Copy Number Variations in the Plectin (PLEC) Gene on the Growth Traits and Meat Quality of Leizhou Black Goats. Animals 2023, 13, 3651. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, L.; Lin, X.; Peng, P.; Shen, W.; Tang, S.; Lan, X.; Wan, F.; Yin, Y.; Liu, M. Effects of Genetic Variation of the Sorting Nexin 29 (SNX29) Gene on Growth Traits of Xiangdong Black Goat. Animals 2022, 12, 3461. [Google Scholar] [CrossRef]
- Wang, K.; Kang, Z.; Jiang, E.; Yan, H.; Zhu, H.; Liu, J.; Qu, L.; Lan, X.; Pan, C. Genetic effects of DSCAML1 identified in genome-wide association study revealing strong associations with litter size and semen quality in goat (Capra hircus). Theriogenology 2020, 146, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Rousset, F. genepop’007: A complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yan, H.; Xu, H.; Yang, Q.; Zhang, S.; Pan, C.; Chen, H.; Zhu, H.; Liu, J.; Qu, L.; et al. A novel indel within goat casein alpha S1 gene is significantly associated with litter size. Gene 2018, 671, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, Y.; Cao, Z.; Du, M.; Hao, Y.; Pan, J.; He, H. Irisin promotes cementoblast differentiation via p38 MAPK pathway. Oral. Dis. 2020, 26, 974–982. [Google Scholar] [CrossRef]
- Ye, X.; Shen, Y.; Ni, C.; Ye, J.; Xin, Y.; Zhang, W.; Ren, Y. Irisin reverses insulin resistance in C2C12 cells via the p38-MAPK-PGC-1α pathway. Peptides 2019, 119, 170120. [Google Scholar] [CrossRef]
- Fukushima, Y.; Kurose, S.; Shinno, H.; Cao Thi Thu, H.; Tamanoi, A.; Tsutsumi, H.; Hasegawa, T.; Nakajima, T.; Kimura, Y. Relationships between serum irisin levels and metabolic parameters in Japanese patients with obesity. Obes. Sci. Pract. 2016, 2, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Post, K.L.; Belmadani, M.; Ganguly, P.; Meili, F.; Dingwall, R.; McDiarmid, T.A.; Meyers, W.M.; Herrington, C.; Young, B.P.; Callaghan, D.B.; et al. Multi-model functionalization of disease-associated PTEN missense mutations identifies multiple molecular mechanisms underlying protein dysfunction. Nat. Commun. 2020, 11, 2073. [Google Scholar] [CrossRef] [PubMed]
- Mitui, M.; Nahas, S.A.; Du, L.T.; Yang, Z.; Lai, C.H.; Nakamura, K.; Arroyo, S.; Scott, S.; Purayidom, A.; Concannon, P.; et al. Functional and computational assessment of missense variants in the ataxia-telangiectasia mutated (ATM) gene: Mutations with increased cancer risk. Hum. Mutat. 2009, 30, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Spanakis, E.; Milord, E.; Gragnoli, C. AVPR2 variants and mutations in nephrogenic diabetes insipidus: Review and missense mutation significance. J. Cell Physiol. 2008, 217, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Corso, G.; Feroce, I.; Intra, M.; Toesca, A.; Magnoni, F.; Sargenti, M.; Naninato, P.; Caldarella, P.; Pagani, G.; Vento, A.; et al. BRCA1/2 germline missense mutations: A systematic review. Eur. J. Cancer Prev. 2018, 27, 279–286. [Google Scholar] [CrossRef] [PubMed]
- GB/T 35892-2018; Laboratory Animal—Guideline for Ethical Review of Animal Welfare. National Laboratory Animal Standardization Technical Committee: Beijing, China, 2018.


{kind=link}
{kind=link}
{kind=link}
| Loci | Size | Genotypic Frequencies | HWE p-Value | Population Parameters | |||||
|---|---|---|---|---|---|---|---|---|---|
| n | Ref | Ref/Mut | Mut | Ho | He | Ne | PIC | ||
| SNP1 p.119A/V | 792 | 0.866 | 0.128 | 0.006 | p < 0.0.5 | 0.870 | 0.130 | 1.150 | 0.122 |
| SNP2 p.135R/H | 795 | 0.883 | 0.117 | - | p < 0.0.5 | 0.890 | 0.110 | 1.124 | 0.104 |
| SNP3 p.170W/G | 790 | 0.876 | 0.118 | 0.006 | p < 0.0.5 | 0.884 | 0.116 | 1.132 | 0.110 |
| Traits | Genotypes (Mean ± SE) | p-Values | ||
|---|---|---|---|---|
| Ref | Ref/Mut | Mut | ||
| body height (cm) | 52.24 ± 0.14 | 52.07 ± 0.19 | 51.88 ± 0.14 | 0.317 |
| body oblique length (cm) | 54.67 ± 0.21 | 54.72 ± 0.14 | 54.92 ± 0.18 | 0.288 |
| chest circumference (cm) | 59.07 a ± 0.19 | 58.88 a ± 0.21 | 57.79 b ± 0.12 | 0.034 |
| body weight (kg) | 20.29 a ± 0.27 | 19.31 b ± 0.18 | 19.57 b ± 0.07 | 0.011 |
| cannon circumference (cm) | 7.27 ± 0.05 | 7.19 ± 0.03 | 7.23 ± 0.04 | 0.152 |
| carcass weight (kg) | 9.71 a ± 0.19 | 9.84 a ± 0.23 | 9.01 b ± 0.15 | 0.027 |
| cross-section area of longissimus dorsi lumbois muscle (cm2) | 7.84 a ± 0.17 | 7.75 a ± 0.06 | 7.24 b ± 0.12 | 0.043 |
| water loss rate (%) | 4.85 ± 0.10 | 4.88 ± 0.24 | 4.77 ± 0.31 | 0.302 |
| water-holding capacity (%) | 4.82 ± 0.24 | 4.78 ± 0.13 | 4.84 ± 0.16 | 0.424 |
| shear stress (N) | 48.09 ± 0.37 | 48.38 ± 0.22 | 47.59 ± 0.41 | 0.087 |
| Traits | Genotypes (Mean ± SE) | p-Values | ||
|---|---|---|---|---|
| Ref | Ref/Mut | - | ||
| body height (cm) | 51.64 ± 0.30 | 51.97 ± 0.14 | 0.425 | |
| body oblique length (cm) | 54.73 ± 0.25 | 54.34 ± 0.10 | 0.174 | |
| chest circumference (cm) | 57.94 ± 0.13 | 57.23 ± 0.28 | 0.027 | |
| body weight (kg) | 19.99 ± 0.24 | 19.27 ± 0.09 | 0.043 | |
| cannon circumference (cm) | 7.18 ± 0.04 | 7.21 ± 0.05 | 0.684 | |
| carcass weight (kg) | 9.75 ± 0.24 | 9.31 ± 0.26 | 0.058 | |
| cross-section area of longissimus dorsi lumbois muscle (cm2) | 7.87 ± 0.22 | 7.11 ± 0.29 | 0.007 | |
| water loss rate (%) | 4.95 ± 0.21 | 4.31 ± 0.17 | 0.041 | |
| water-holding capacity (%) | 4.83 ± 0.14 | 4.69 ± 0.19 | 0.026 | |
| shear stress (N) | 49.10 ± 0.26 | 48.98 ± 0.37 | 0.073 | |
| SNP Haplotype | SNP1 p.119A/V C>T | SNP2p.135R/H G>A | SNP3p.170W/G T>G | Frequency |
|---|---|---|---|---|
| HI | C | G | T | 0.872 |
| H2 | C | G | G | 0.046 |
| H3 | C | A | T | 0.011 |
| H4 | T | A | T | 0.032 |
| H5 | T | A | G | 0.020 |
| H6 | T | G | T | 0.019 |
| Combined Genotype | BH (cm) | BOL (cm) | CC (cm) | BW (kg) | CAC (cm) | CW (kg) | CALM (cm2) | WLR (%) | WHC (%) | SS (N) |
|---|---|---|---|---|---|---|---|---|---|---|
| CCGGTT | 51.79 ± 0.27 | 54.83 a ± 0.32 | 58.78 a ± 0.37 | 20.09 a ± 0.27 | 7.21 ± 0.08 | 9.81 a ± 0.20 | 7.80 a ± 0.29 | 4.73 a ± 0.18 | 4.68 ± 0.24 | 48.89 ± 0.23 |
| CCGGTG | 51.82 ± 0.19 | 54.71 a ± 0.27 | 58.69 ab ± 0.25 | 19.88 a ± 0.31 | 7.20 ± 0.07 | 9.68 a ± 0.33 | 7.73 a ± 0.29 | 4.69 a ± 0.21 | 4.72 ± 0.18 | 49.01 ± 0.15 |
| CCGATT | 51.77 ± 0.34 | 54.68 ab ± 0.38 | 58.47 b ± 0.21 | 19.75 a ± 0.28 | 7.23 ± 0.09 | 9.72 a ± 0.14 | 7.88 a ± 0.29 | 4.80 a ± 0.12 | 4.62 ± 0.31 | 48.79 ± 0.38 |
| CTGATT | 51.62 ± 0.22 | 54.75 a ± 0.40 | 58.23 bc ± 0.41 | 19.64 ab ± 0.16 | 7.22 ± 0.09 | 9.60 ab ± 0.19 | 7.28 ab ± 0.29 | 4.73 a ± 0.25 | 4.42 ± 0.27 | 48.88 ± 0.27 |
| TTGATT | 51.80 ± 0.43 | 54.70 a ± 0.38 | 57.62 c ± 0.53 | 19.48 b ± 0.38 | 7.19 ± 0.21 | 8.95 b ± 0.49 | 7.01 b ± 0.41 | 4.27 b ± 0.42 | 4.66 ± 0.36 | 48.75 ± 0.51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Xu, M.; Zhang, Y.; Han, J.; Zhou, H.; Wang, K. Missense Mutations in FDNC5 Associated with Morphometric Traits and Meat Quality in Hainan Black Goats. Animals 2025, 15, 565. https://doi.org/10.3390/ani15040565
Huang J, Xu M, Zhang Y, Han J, Zhou H, Wang K. Missense Mutations in FDNC5 Associated with Morphometric Traits and Meat Quality in Hainan Black Goats. Animals. 2025; 15(4):565. https://doi.org/10.3390/ani15040565
Chicago/Turabian StyleHuang, Jing, Mengning Xu, Yuelang Zhang, Jiancheng Han, Hanlin Zhou, and Ke Wang. 2025. "Missense Mutations in FDNC5 Associated with Morphometric Traits and Meat Quality in Hainan Black Goats" Animals 15, no. 4: 565. https://doi.org/10.3390/ani15040565
APA StyleHuang, J., Xu, M., Zhang, Y., Han, J., Zhou, H., & Wang, K. (2025). Missense Mutations in FDNC5 Associated with Morphometric Traits and Meat Quality in Hainan Black Goats. Animals, 15(4), 565. https://doi.org/10.3390/ani15040565