
Taxonomic Revision of the Relationship Between Coproptilia and Nosphistica (Lepidoptera: Lecithoceridae) with Descriptions of Two New Species and a New Record from China


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
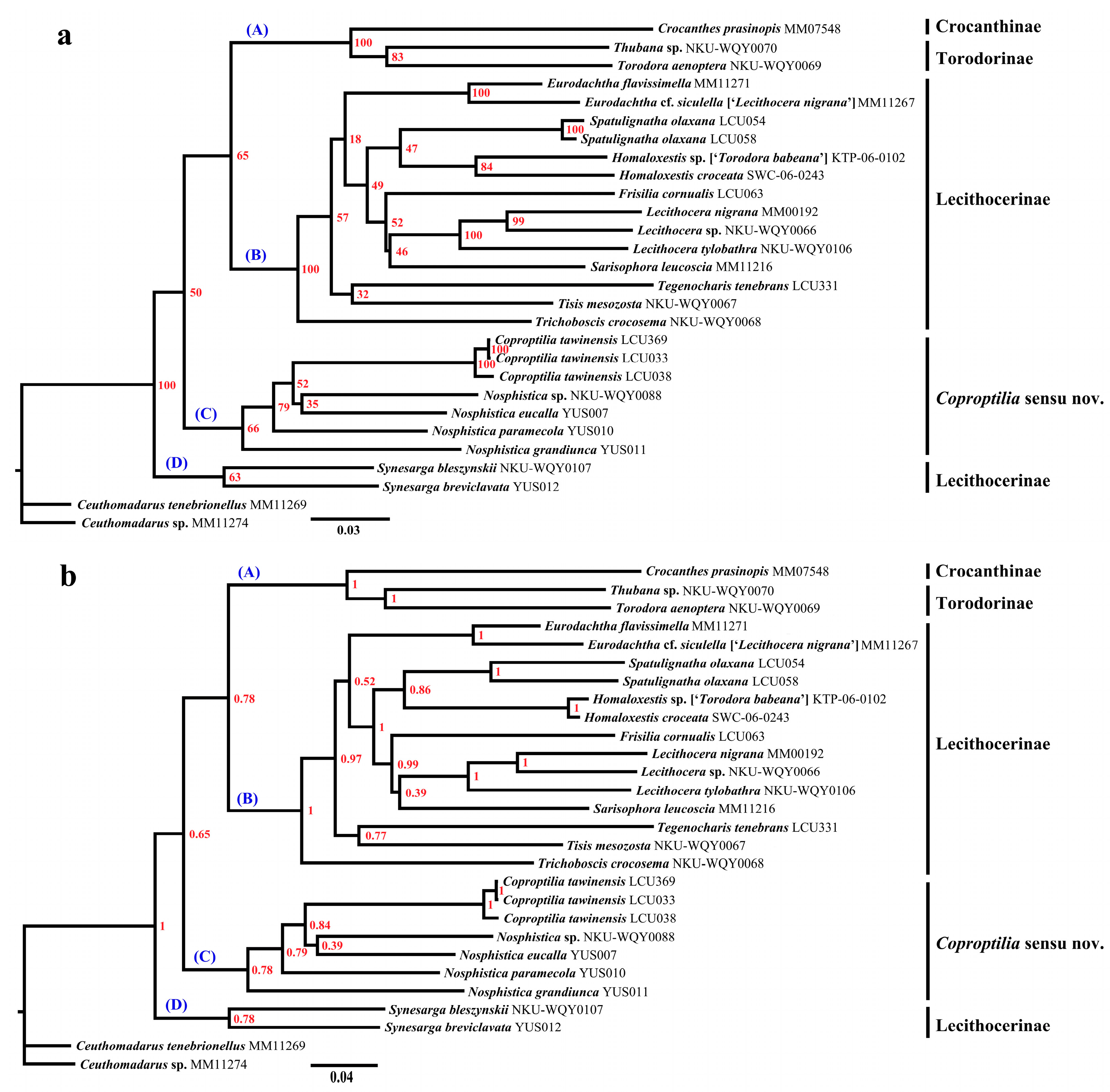
3.1. Molecular Analysis Results
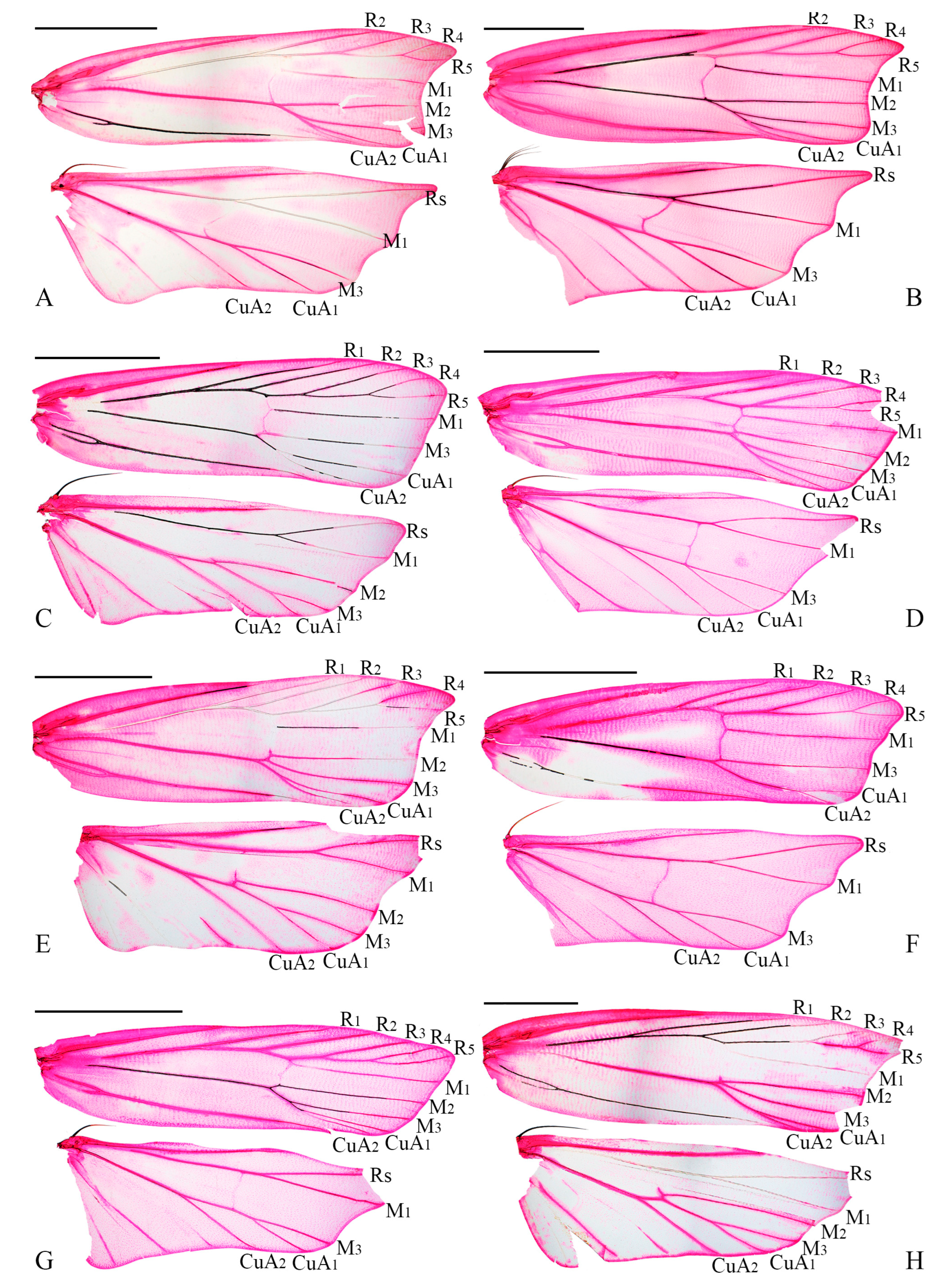
3.2. Morphological Results
3.2.1. Coproptilia tawiensis Park, 2009
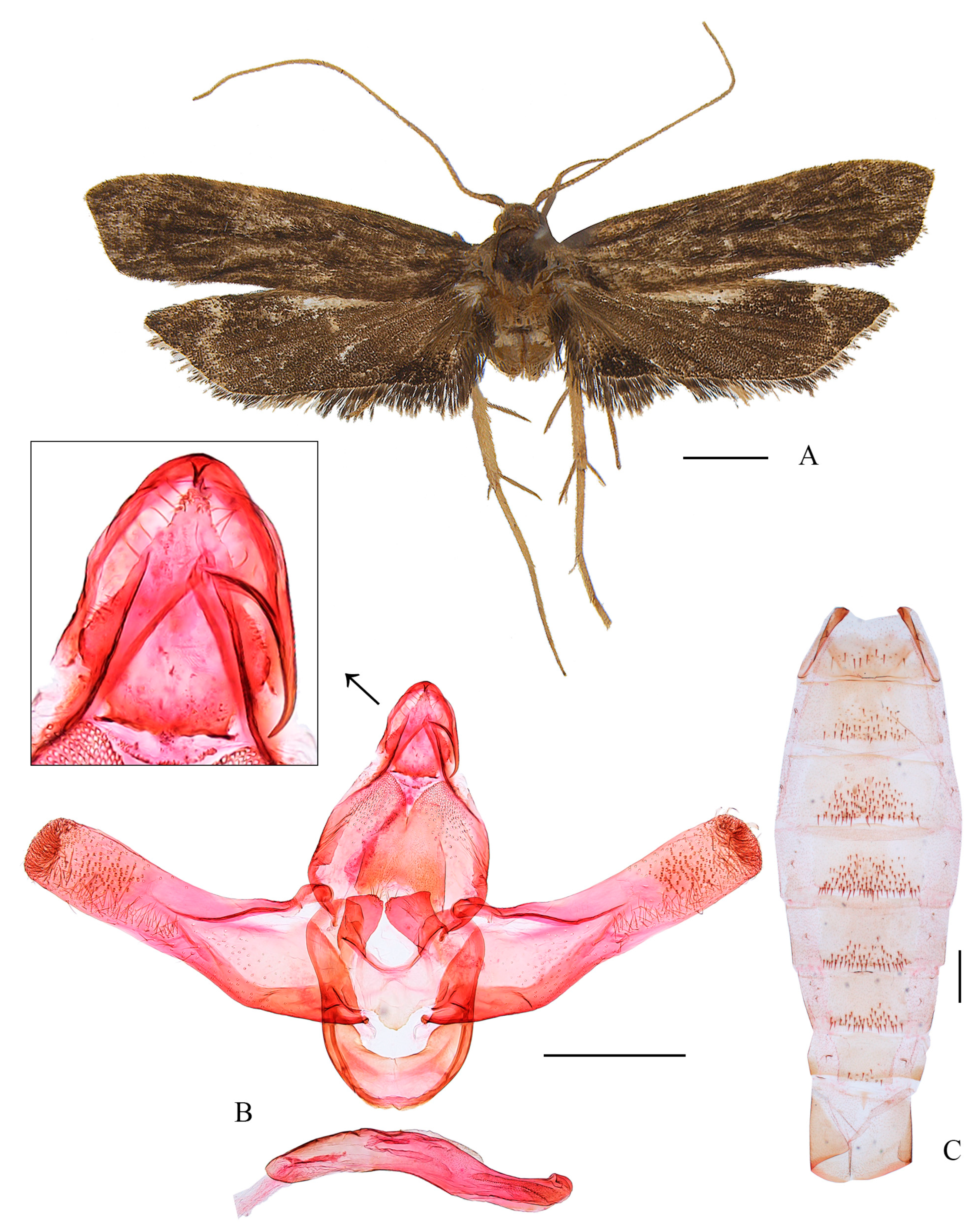
3.2.2. Coproptilia uniformis Yu, sp. nov. (Figure 3)
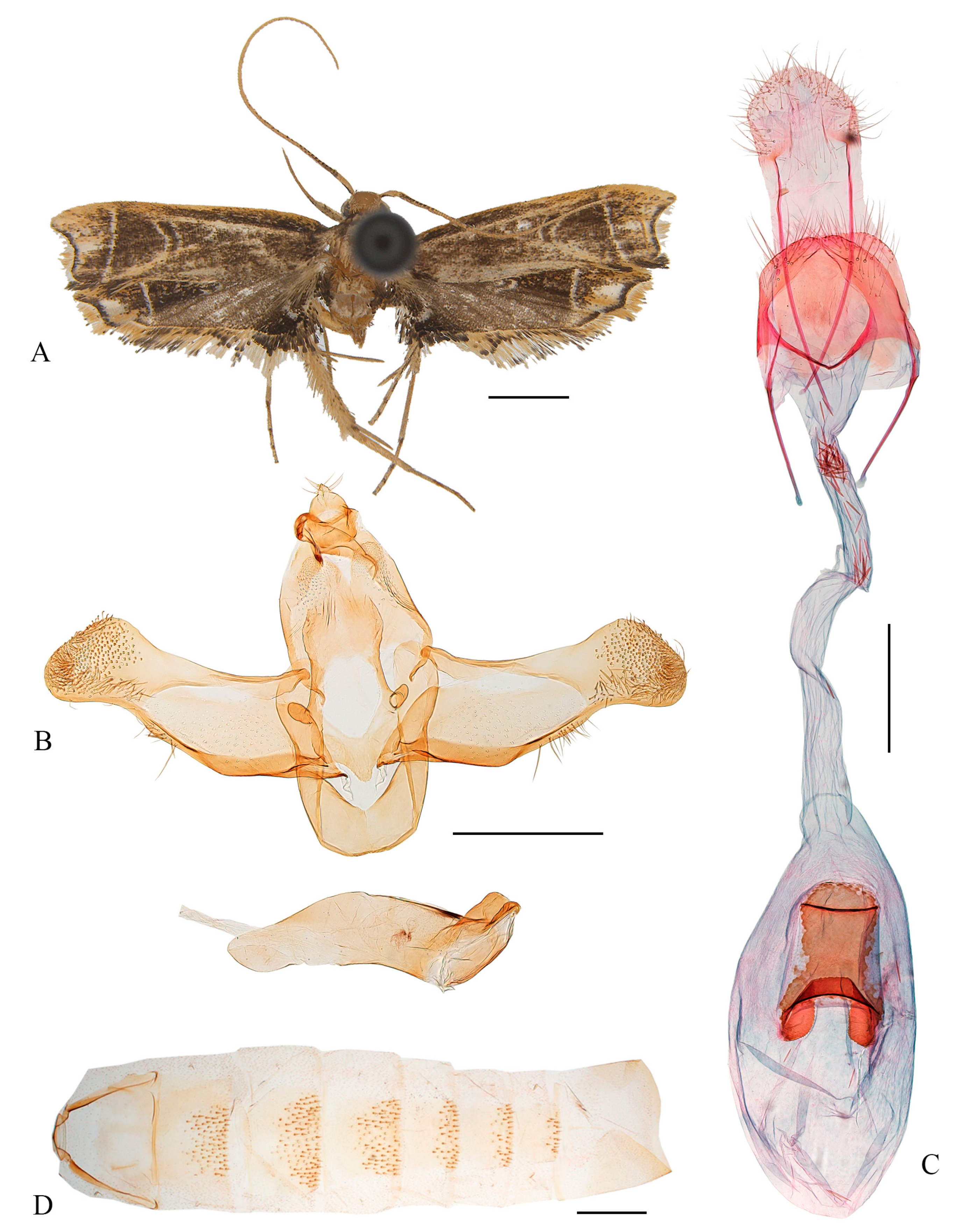
3.2.3. Coproptilia funiuensis Yu, sp. nov. (Figure 4)

4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Q.Y.; Li, H.H. Phylogeny of the superfamily Gelechioidea (Lepidoptera: Obtectomera), with an exploratory application on geometric morphometrics. Zool. Scr. 2020, 49, 307–328. [Google Scholar] [CrossRef]
- Park, K.T.; Cho, S.; Koo, J.M. The Subfamily Torodorinae of the World (Lepidoptera: Lecithoceridae); National Institute of Biological Resources: Incheon, Republic of Korea, 2022; pp. 1–584. [Google Scholar]
- Gozmány, L. Lecithoceridae. In Microlepidoptera Palaearctica; Amsel, H.G., Reisser, H., Gregor, F., Eds.; Georg Fromme & Co.: Vienna, Austria, 1978; Volume 5, pp. 1–306. [Google Scholar]
- Common, I.F.B. Moths of Australia; Melbourne University Press: Melbourne, Australia, 1990; pp. 1–544. [Google Scholar]
- Komai, F.; Yoshiyasu, Y.; Nasu, Y.; Saito, T. A Guide to the Lepidoptera of Japan; Tokai University Press: Tokyo, Japan, 2011; pp. 1–1308. [Google Scholar]
- Park, K.T.; Mey, W. A review of the genus Lecithocera Herrich-Schäffer, 1853 in the Philippines, with descriptions of seven new species (Lepidoptera: Lecithoceridae). SHILAP Revista de Lepidopterología 2016, 33, 339–352. [Google Scholar] [CrossRef]
- Snellen, P.C.T. Beschrjvingen van nieuwe exotische Tortricinen, Tincinen, en Pterophorinen benevens aanteekeningen over reeds bekend gemaakte soorten. Tikdschrift vorr Rntomologie 1903, 46, 25–26. [Google Scholar]
- Wu, C.S. The Lecithoeridae (Lepidoptera) of China, with descriptions of new taxa. Sinzoologica 1994, 11, 123–154. [Google Scholar]
- Park, K.T. Revision of the genus Coproptilia Snellen, with description of a new species from the Philippines (Lepidoptera: Lecithoceridae). Entomol. Res. 2009, 39, 239–242. [Google Scholar] [CrossRef]
- Wu, C.S. Fauna Sinica, Insecta, Lepidoptera, Lecithoceridae; Science Press: Beijing, China, 1997; pp. 1–306. [Google Scholar]
- Meyrick, E. Descriptions of Indian Micro-lepidoptera. J. Bombay Nat. Hist. Soc. 1911, 20, 706–736. [Google Scholar]
- Park, K.T. A revision of the genus Nosphistica Meyrick (Lepidoptera, Lecithoceridae). Zool. Stud. 2002, 41, 251–262. [Google Scholar]
- Yu, S.; Wang, S.X. Taxonomic study of the genus Nosphistica Meyrick, 1911 (Lepidoptera: Lecithoceridae) from China, with descriptions of seven new species. Zootaxa 2019, 4664, 497–517. [Google Scholar] [CrossRef] [PubMed]
- Sterling, M.J.; Lees, D.C.; Grundy, D. Xenotorodor stygioxanthus gen. nov., sp. nov. (Lepidoptera, Lecithoceridae, Torodorinae), described from an established population in Spain with discussion of taxonomic placement. Nota Lepidopterol. 2023, 46, 103–123. [Google Scholar] [CrossRef]
- Li, H.H. The Gelechiidae of China (I) (Lepidoptera: Gelechioidea); Nankai University Press: Tianjin, China, 2002. [Google Scholar]
- Folmer, O.; Black, M.B.; Hoch, W.; Lutz, R.A.; Vrijehock, R.C. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Cho, S.W.; Mitchell, A.; Regier, J.C.; Mitter, C.; Poole, R.W.; Friedlander, T.P.; Zhao, S.W. A highly conserved nuclear gene for low-level phylogenetics: Elongation factor-1α recovers morphology-based tree for heliothine moths. Mol. Biol. Evol. 1995, 12, 650–656. [Google Scholar] [PubMed]
- Brower, A.V.Z.; DeSalle, R. Patterns of mitochondrial versus nuclear DNA sequence divergence among nymphalid butterflies: The utility of wingless as a source of characters for phylogenetic inference. Insect Mol. Biol. 1998, 7, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Wahlberg, N.; Wheat, C.H. Genomic outposts serve the phylogenetic pioneers: Designing novel nuclear markers for genomic DNA extractions of Lepidoptera. Syst. Biol. 2008, 57, 231–242. [Google Scholar] [CrossRef]
- Kaila, L.; Mutanen, M.; Nyman, T. Phylogeny of the mega-diverse Gelechioidea (Lepidoptera): Adaptations and determinants of success. Mol. Phylogenet. Evol. 2011, 61, 801–809. [Google Scholar] [CrossRef]
- Regier, J.C.; Mitter, C.; Solis, M.A.; Hayden, J.E.; Landry, B.; Nuss, M.; Simonsen, T.J.; Yen, S.-H.; Zwick, A.; Cummings, M.P. A molecular phylogeny for the pyraloid moths (Lepidoptera: Pyraloidea) and its implications for higher-level classification. Syst. Entomol. 2012, 37, 635–656. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef]
- Meyrick, E. Exotic Microlepidoptera II; Thornhanger: Marlborough, UK, 1918; pp. 97–224. [Google Scholar]
- Yu, S.; Zhu, Y.M.; Wang, S.X. Eighteen new species and fifteen new records of the genus Torodora Meyrick (Lepidoptera: Lecithoceridae) from China. Zootaxa 2022, 5133, 1–39. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Region | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) | ||
|---|---|---|---|---|
| Elongation factor 1 alpha (EF-1α) | EF-1α-F | CCYGCCAAYATCACCACTGAAG | EF-1α-R | AGAGGHGGGAACTCYTGGAAGGA |
| Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | GAPDH-F | TCACTTGGAVGGTGGHGCCAAGAA | GAPDH-R | AGAGAGATACCAGCDGCAGCATC |
| Carbamoyl phosphate synthetase domain protein (CAD) | CAD-F | AGTTTRGACTACTGTGTAGTTAAAATA | CAD-R | TGATAAAATAACGCCATCAGGA |
| Cytosolic malate dehydrogenase (MDH) | MDH-F | TGTTGTCATGGAGCTTGCAGATT | MDH-R | CCCATATAACAACATTCTTWACATCC |
| Ribosomal protein S5 (RpS5) | RpS5-F | GCAGCATGGCCGTCGATAACAT | RpS5-R | TTGATGAACCCTTGGCAGCATTAAT |
| Wingless | wingless-F | TGCACAGTGAAAACTTGCTGGAT | wingless-R | GTTACACCTTTCCACAACGAACATG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Yu, S. Taxonomic Revision of the Relationship Between Coproptilia and Nosphistica (Lepidoptera: Lecithoceridae) with Descriptions of Two New Species and a New Record from China. Animals 2025, 15, 426. https://doi.org/10.3390/ani15030426
Li H, Yu S. Taxonomic Revision of the Relationship Between Coproptilia and Nosphistica (Lepidoptera: Lecithoceridae) with Descriptions of Two New Species and a New Record from China. Animals. 2025; 15(3):426. https://doi.org/10.3390/ani15030426
Chicago/Turabian StyleLi, Haotian, and Shuai Yu. 2025. "Taxonomic Revision of the Relationship Between Coproptilia and Nosphistica (Lepidoptera: Lecithoceridae) with Descriptions of Two New Species and a New Record from China" Animals 15, no. 3: 426. https://doi.org/10.3390/ani15030426
APA StyleLi, H., & Yu, S. (2025). Taxonomic Revision of the Relationship Between Coproptilia and Nosphistica (Lepidoptera: Lecithoceridae) with Descriptions of Two New Species and a New Record from China. Animals, 15(3), 426. https://doi.org/10.3390/ani15030426