4. Discussion
The pathogenesis of asthma is closely related to genetic, neurological, and immune factors, as well as airway inflammation [
23,
24]. Recently, metabolomics has emerged as a promising tool in multiple fields of biomedicine. In human and animal models, plasma- or serum-based metabolomic studies have elucidated the associations of amino acids, nucleic acid metabolites, and lipid derivatives with asthma and identified the metabolic characteristics of asthma [
25]. Serum lipid mediators, hormone concentrations, and blood uric acid levels are associated with asthma severity.
Asthma is a complex disease with multiple etiologies. Metabolomics provides a comprehensive platform to understand the pathophysiological basis of various diseases. As a bridge between the phenotype and genome [
26], metabolomics can amplify phenotypic or genomic differences, help identify differences in traits in addition to phenotypic and genetic differences between different germplasms, and improve our understanding of the differences between different germplasms.
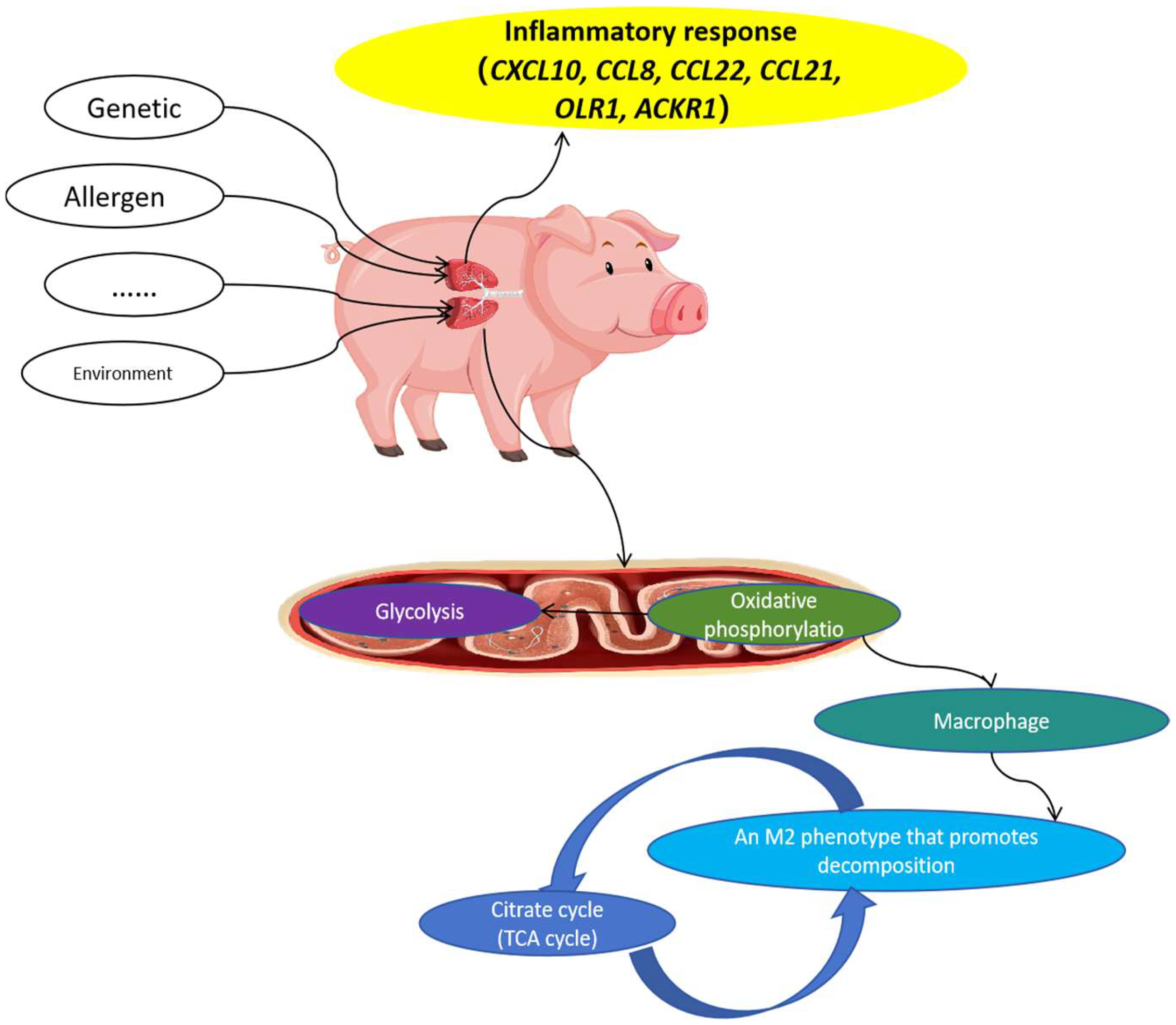
In this study, although we have focused on the molecular mechanisms underlying asthma in Meishan pigs through transcriptomics and metabolomics, the role of Mhp infection in the pathogenesis of asthma in these pigs cannot be overlooked.
Mitochondria are the main intracellular consumers of O
2 and are an important source of oxidative stress [
27]. We found that the oxidative phosphorylation pathway in lung mitochondria was disrupted in asthmatic Meishan pigs. Mitochondria produce ROS via electron leakage and molecular oxygen reduction in the respiratory chain [
28]. The precursor of ROS is O
2−, which is produced by mitochondria and NADPH oxidase. Under normal circumstances, 1–3% of the oxygen passing through the mitochondria is used to form ROS. A portion of the energy in all animal cells originates from oxidative phosphorylation in the mitochondria, and the reduction of O
2 to H
2O by complexes I and III of the mitochondrial respiratory chain produces intermediate ROS in the mitochondria [
29]. Various antioxidants exist in animals, including antioxidant enzymes, such as superoxide dismutase and glutathione peroxidase, and non-enzymatic systems. They can directly remove free oxygen radicals and other oxidative molecules, regenerate damaged biomolecules, and maintain a relative balance between ROS and antioxidant systems. Overproduction of ROS and/or reduced function of the antioxidant defense system leads to cellular oxidative stress, which promotes inflammation and asthma, including airflow obstruction, airway hyperreactivity, and airway remodeling. Our results suggest that the disruption of the oxidative phosphorylation pathway in lung mitochondria may lead to an imbalance between ROS production and antioxidant defense, contributing to the development of asthma in Meishan pigs.
The TCA cycle occurs in the cytoplasm and does not depend on oxygen [
30]. We observed significant changes in the contents of metabolites related to the TCA cycle, such as succinic acid, riboflavin—5—phosphate, fumaric acid, alpha-ketoglutaric acid, and citric acid, in asthmatic Meishan pigs compared to healthy ones. Lactic acid is produced under the action of lactate dehydrogenase, with much greater quantities of lactic acid produced in normal lung tissue than in other tissues [
31]. These alterations in the TCA cycle may affect energy metabolism and cellular function, further contributing to the pathogenesis of asthma. Previous studies have also shown that metabolite changes in the TCA cycle are associated with asthma severity [
32]. Our findings are in line with these reports and suggest that the disturbance of the TCA cycle may play an important role in the development of asthma in Meishan pigs.
Glutathione S-transferases (GSTs) are thought to protect cells from ROS [
33]. One of the latest concepts related to asthma to be investigated is the role of GSTs [
34]. GSTs are major phase II enzymes involved in metabolic detoxification [
35]. Inflammation is key to asthma, and reactive oxygen intermediates that play a role in inflammation are metabolized by GSTs; therefore, abnormalities in GSTs may lead to asthma [
36,
37]. Although we did not directly measure GST activity in this study, the observed oxidative stress and inflammation in asthmatic Meishan pigs suggest that GSTs may be involved in the pathophysiology of asthma. Future studies could focus on the expression and activity of GSTs in Meishan pigs with asthma to further clarify their role.
Mhp is a significant pathogen that can colonize the respiratory tract of pigs, leading to chronic respiratory diseases. Infection with Mhp can disrupt the normal physiological functions of the respiratory epithelium, which in turn may trigger a series of immune responses and inflammatory cascades. The immune response to Mhp infection is complex and involves both the innate and adaptive immune systems. Arginase 1 (
Arg1), an enzyme that catalyzes the conversion of arginine to ornithine, is a hallmark of immune-regulating M2 macrophages that produce IL-10 [
38]. Our study found that the expression of
Arg1 and related genes may be involved in the immune regulation of asthma in Meishan pigs. During influenza infection in mice, induction of
Arg1 expression is a key feature of lung CD4+ T cells. Conditional ablation of
Arg1 in CD4+ T cells accelerates the virus-specific Th1 response and its resolution, leading to effective viral clearance and reduced lung pathology. Unbiased transcriptomics and metabolomics studies have shown that
Arg1 deficiency, unlike
Arg2 deficiency, leads to alterations in glutamine metabolism [
39,
40]. Rebalancing this disturbed glutamine flux normalizes the cellular Th1 response. Normal
Arg1 activity allows arginine to produce ornithine, which is essential for cells, thus ensuring optimal glutamine flux into the TCA cycle [
41]. In the absence of
Arg1, the glutamine compensatory reaction reduces TCA activity for ornithine production, thereby affecting the kinetics of the Th1 response [
42]. Overall,
Arg1, which is inherent in CD4+ T cells, can be regarded as a rheostat that regulates the mammalian Th1 life cycle. Recently, Th17 cells, such as CD4+ T cells, have been shown to regulate the involvement of neutrophils in airway inflammation and airway reconstruction in asthma. Our results suggest that Arg1 may play a similar role in the immune regulation of asthma in Meishan pigs, and further studies are needed to elucidate its detailed mechanism.
RORC is an upstream regulatory gene and the main effector of Th17 cells [
43,
44].
RORC is involved in asthma pathogenesis via Th17 cells downstream of
RORC.
RORC mRNA expression in lung tissue is significantly upregulated in patients with asthma, while IL-17 in the peripheral blood, bronchial lavage fluid, and sputum is significantly increased and positively correlated with airway hyperreactivity [
45,
46]. In our study, we also observed changes in the expression of
RORC and related genes in asthmatic Meishan pigs. One study found that
RORC mRNA levels in peripheral blood mononuclear cells of children with asthma during acute attacks were significantly higher than those in the remitted and normal control groups.
RORC mRNA expression decreased to normal, whereas the IL-17 level was still significantly higher than that of the normal control group, indicating that
RORC aggravates airway inflammation and airway hyperreactivity by promoting transcription of the cytokine IL-17 during acute asthma attacks [
47]. Previous studies have shown that
RORC promoter methylation is low in obesity-related asthma. The immune response to allergic asthma is primarily regulated by Th2 lymphocytes, which release IL-4, IL-5, and IL-13. Thus, B cells and eosinophils recruit specific IgE molecules to the respiratory epithelium. RORC is required for naive CD4+ T cells to differentiate into Th17 lymphocytes;
RORC mRNA expression is higher in obesity-related asthma, while methylation of the
RORC promoter is lower [
48,
49]. It is speculated that
RORC expression is affected by the addition of a methyl group to the carbon of CpG cytosine. Our results are consistent with these previous findings and suggest that
RORC and Th17 cells may play an important role in the pathogenesis of asthma in Meishan pigs.
Argl is an important enzyme that regulates macrophage function [
50] and is a marker of selective macrophage activation. Alveolar macrophages are important sentinels in host lung defense, playing vital roles in maintaining immune regulation, pathogen clearance, and homeostasis [
51,
52]. M1 macrophages are an important source of many inflammatory cytokines, including TNF-α, IL-1, IL-12, IL-18, and IL-23, which have been identified as important mediators and drivers of chronic inflammatory and autoimmune diseases. The inflammatory response is caused by the aggregation of Th2 lymphocytes, mast cells, eosinophils, and macrophages in the lungs, and is related to the M2 polarization of macrophages [
53]. Macrophages are important regulators of allergic asthma and initiators of inflammatory responses associated with lung injury, fibrosis, and goblet cell proliferation [
54]. Pulmonary macrophages produce a variety of factors that directly stimulate airway, smooth muscle contractile force and extracellular matrix degradation, and participate in airway pathological remodeling. Analysis of bronchial biopsy specimens revealed an increase in CD206 macrophages in patients with asthma, demonstrating a correlation between the percentage of M2 macrophages and disease severity [
55,
56,
57]. Many circulating M2-like phenotypes have been observed in patients with allergic and bronchial asthma. Additionally, in response to bronchial allergens, macrophages in patients with asthma undergo M2 polarization, thereby supporting Th2-related inflammation. Although airway disease is associated with Th2/M2 inflammation, M1 macrophages may participate in the pathogenesis of asthma by releasing inflammatory cytokines and NO, thereby exacerbating lung injury and airway remodeling. In our study, we found that macrophage polarization and function may be altered in asthmatic Meishan pigs, which is consistent with previous reports. Future studies could further investigate the role of macrophages in the development and progression of asthma in Meishan pigs.
The Notch pathway plays an important role in the occurrence and development of asthma. The Notch pathway affects lung tissue development, determines the direction of cell differentiation, and regulates the development of alveoli and pulmonary blood vessels [
58]. The Notch pathway, which is known to be involved in cell differentiation and immune regulation, may also be modulated by Mhp infection. Mhp-induced cytokines and growth factors could potentially affect the expression and activation of Notch receptors and ligands, thereby altering the Notch signaling pathway. The Notch pathway is involved in T-cell regulation, which affects Th17 cells, Tregs, dendritic cell expression, and other pathways by altering the Th1/Th2 balance, leading to the occurrence and development of asthma [
59]. The Notch pathway also participates in the pathological changes in airway remodeling in asthma by altering the infiltration of various inflammatory cells, such as lymphocytes and eosinophils, promoting the metaplasia of airway goblet cells and airway mucus secretion [
60]. Notch signaling molecules are dynamically expressed during lung development and may play a key role in regulating the differentiation and development of the alveolar epithelium and vascular endothelial cells. Some studies have reported that the application of gamma-secretase inhibitor DAPT to inhibit the Notch signaling pathway can inhibit angiotensin II-induced pulmonary vascular remodeling and reduce pulmonary artery pressure [
61,
62,
63], providing a new idea for the treatment of pulmonary hypertension. However, the regulatory mechanisms of the Notch pathway have not been fully elucidated and more in-depth studies are needed. Our results suggest that the Notch pathway may be involved in the pathogenesis of asthma in Meishan pigs, and further studies are warranted to explore its detailed mechanism and potential therapeutic targets. However, the value of its application in clinical treatment requires further exploration.
YPEL family proteins are located in centrosomes near the interphase nucleolus and mitotic organs during mitosis [
64]. Based on their subcellular localization, YPEL4 may play an important role in the cell cycle and proliferation. YPEL4 may also mediate adrenal cell proliferation by regulating the mitogen-activated protein kinase signaling pathway [
65]. This molecular mechanism may also play an important role in lung diseases. Although the function of YPEL4 is largely unknown, further research may confirm its functional importance and the underlying molecular processes in the lungs and other diseases, which would make YPEL4 a therapeutic target. In our study, we observed changes in the expression of
YPEL4 and related genes in asthmatic Meishan pigs, suggesting that
YPEL4 may be involved in the cell proliferation and airway remodeling processes in asthma. Future studies could focus on elucidating the exact role of YPEL4 in the pathogenesis of asthma.
Asthma is characterized by an increased proliferation of smooth muscle cells in the airway walls [
66]. Hasaneen et al. studied the dual ERK and phosphatidylinositol 3-kinase (PI3K) pathways and their regulation of airway smooth muscle cell proliferation in asthmatics [
67]. In non-asthmatic cells, growth is controlled by mitogens, which pass through dual signaling pathways, namely the ERK-and PI3K-dependent pathways [
68]. In asthmatic cells, the PI3K pathway is dominant because of the upregulation of the endogenous MAPK inhibitor MAPK phosphatase-I. This inhibitor restricts the ERK signaling pathway in asthmatic cells under mitotic stimulation, making the PI3K pathway dominant. Ultimately, this study suggests that PI3K is an important target for smooth muscle hyperplasia in asthma. Notably, the PI3K signaling pathway plays an important role in the dual pathway between the non-asthmatic PI3K and MAPK signaling pathways. Naturally, the major vault protein (MVP) inhibits YPEL4’s ability to activate the transcription factor Elk-1 in the MAPK signaling pathway. In terms of interactions with YPEL4, specific inhibitors of MVP may allow the MAPK pathway to work alongside the PI3K pathway simultaneously, suggesting that specific inhibitors of MVP may serve as therapeutic targets for asthma by promoting the activity of YPEL4 [
69]. By interacting with MVP, YPEL4 participates in the activities and functions of Elk-1. Thus, YPEL4 plays an important role in regulating the MAPK transduction pathway. Studies have shown that YPEL4 mediates cell cycle progression and cell proliferation. The function of YPEL4 is not fully understood; therefore, further research may contribute to the understanding of YPEL4 as a potential therapeutic target.
The expression of
RAPGEF3, also known as
EPAC, plays a role in neutrophil dysfunction and airway smooth muscle remodeling [
70].
RAPGEF3 may work synergistically with another candidate gene, cadherin-6 (
CDH6), to influence asthma risk [
71]. Both genes have been found to be involved in cell–cell connectivity, which may play a key role in the pathophysiology of asthma. In our study, we also detected changes in the expression of
RAPGEF3 and related genes in asthmatic Meishan pigs, indicating that
RAPGEF3 may be involved in the airway smooth muscle remodeling process in asthma. Future studies could investigate the interaction between
RAPGEF3 and other genes in the context of asthma pathogenesis.
Airway mucus hypersecretion is an important pathophysiological change in asthma and the main cause of death in severe asthma [
72,
73], but the mechanism underlying the regulation of mucus hypersecretion in asthma is still unclear. Mucins are the main component of airway mucus and play important roles in viscoelasticity [
74]. A recent study found that myristoylated alanine-rich C kinase substrate (MARCKS) is a key molecule in mucus secretion [
75]. The secretory inductor PKC phosphorylates MARCKS, which is transferred from the serous membrane to the cell membrane, where MARCKS dephosphorylates, binds to actin and myosin and interacts with the mucous granular membrane [
76]. Through this mechanism, the mucins attach to the contractile cytoskeleton, migrate to the cell periphery, and undergo exocytosis. MARCKS regulates the extracellular secretion of mast cell particles in a PKC-dependent manner by regulating the availability of membrane inosine phosphate, which is required for particle fusion with the plasma membrane [
77]. In our study, we found that the expression of
MARCKS and related genes may be involved in the regulation of airway mucus secretion in asthmatic Meishan pigs. Future studies could further explore the role of
MARCKS in the pathophysiology of asthma and its potential as a therapeutic target.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}