
Evaluation of the Therapeutic Effect of Levamisole on Subclinical Mastitis in Bovine Leukemia Virus-Infected Cows Classified by Proviral Load

 , , , , , , , and
, , , , , , , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Bacterial Culture of Milk Samples
2.3. Measurement of SCC in Milk
2.4. Measurement of Leukocyte Population in Milk
2.5. CBC Analysis
2.6. Blood Biochemistry Analysis
2.7. Measurement of Blood PVL
2.7.1. Isolation of Genomic DNA from Blood
2.7.2. Construction of Molecular Clones
2.7.3. qPCR Analysis for BLV Determination
2.8. Statistical Analysis
- Above-PVL groups: LMS-treated vs. LMS-untreated;
- LMS-treated groups: BLV-positive vs. BLV-negative;
- LMS-treated, BLV-positive groups: Above-PVL vs. Below-PVL.
3. Results
3.1. Cows
3.2. Comparison of Host, Pathogen Factors, CBC, and Blood Biochemistry Parameters (Day 0)
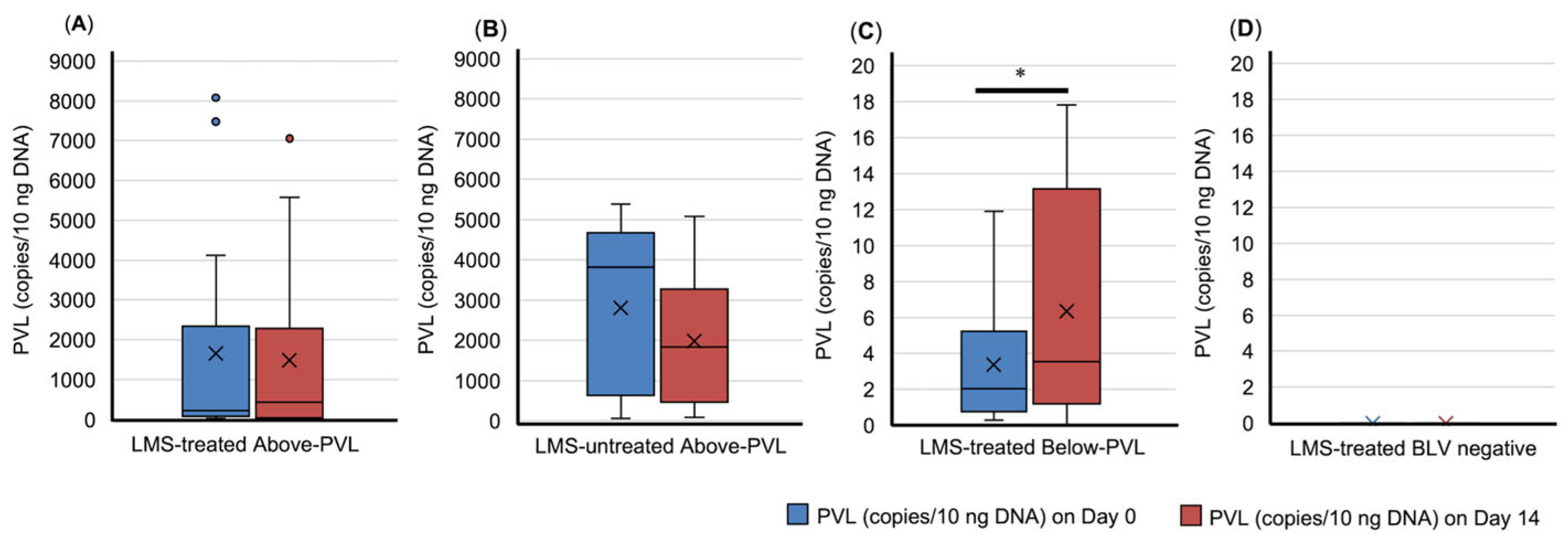
3.3. Dynamics of PVL and Trends in CBC in Peripheral Blood
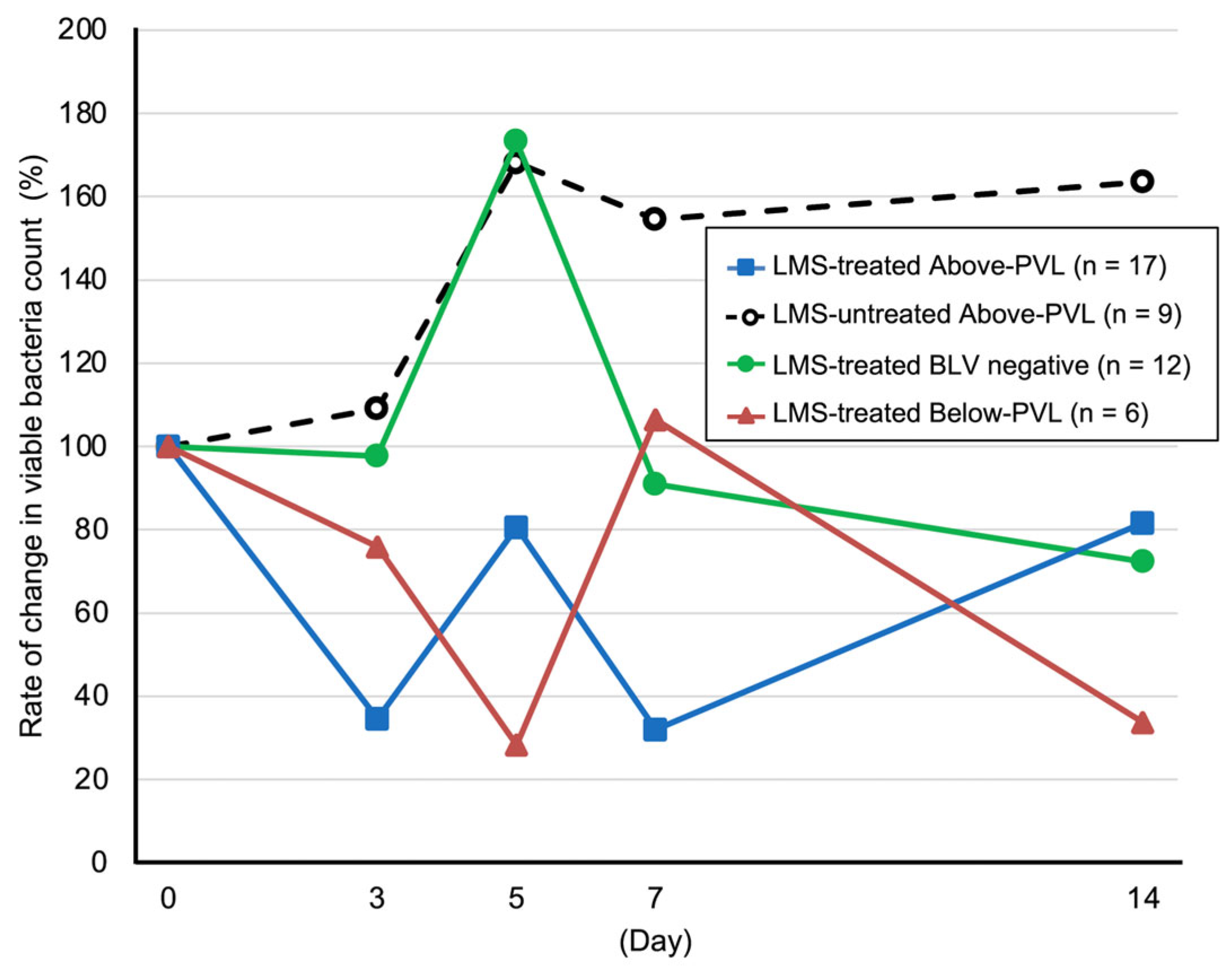
3.4. Rate of Change in the Number of Viable Bacteria
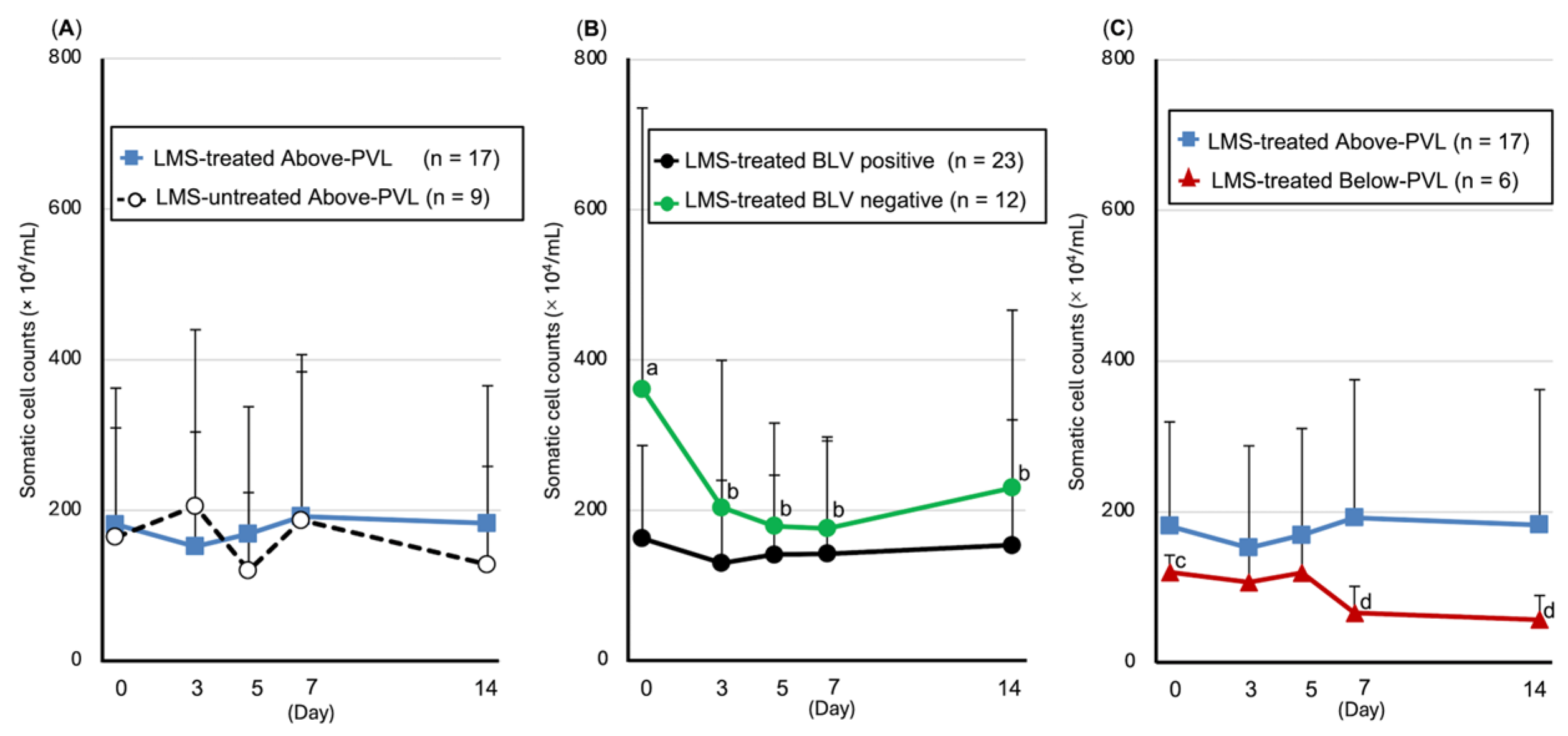
3.5. Trends in SCC and Leukocyte Populations in Milk
3.5.1. SCC
3.5.2. Leukocyte Populations in Milk
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SCC | Somatic cell counts |
| BLV | Bovine leukemia virus |
| EBL | Enzootic bovine leukosis |
| AL | Aleukemic |
| PL | Persistent lymphocytosis |
| PVL | Proviral load |
| LMS | Levamisole |
| CMT | California Mastitis Test |
| CBC | Complete blood count |
| qPCR | Quantitative PCR |
| NMC | National Mastitis Council |
| PBS | Phosphate-buffered saline |
| Hct | Hematocrit |
| WBCs | White blood cells |
| BUN | Blood urea nitrogen |
References
- Kumari, T.; Bhakat, C.; Choudhary, R. A review on subclinical mastitis in dairy cattle. Int. J. Pure Appl. Biosci. 2018, 6, 1291–1299. [Google Scholar] [CrossRef]
- Kawai, K.; Miyazaki, Y.; Obayashi, T.; Oshida, T. Employment of mastitis prevention management program based on HACCP method. Jpn. J. Anim. Hyg. 2010, 35, 147–156. [Google Scholar]
- Murakami, K.; Kobayashi, S.; Konishi, M.; Kameyama, K.; Tsutsui, T. Nationwide survey of bovine leukemia virus infection among dairy and beef breeding cattle in Japan from 2009–2011. J. Vet. Med. Sci. 2013, 75, 1123–1126. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, I.; Lévy, D. Pathobiology of bovine leukemia virus. Vet. Res. 1994, 25, 521–536. [Google Scholar] [PubMed]
- World Organization for Animal Health. Chapter 3.4.9. Enzootic bovine leukosis. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organization for Animal Health (WOAH): Paris, France, 2018; pp. 1–12. [Google Scholar]
- Da, Y.; Shanks, R.D.; Stewart, J.A.; Lewin, H.A. Milk and fat yields decline in bovine leukemia virus-infected Holstein cattle with persistent lymphocytosis. Proc. Natl. Acad. Sci. USA 1993, 90, 6538–6541. [Google Scholar] [CrossRef] [PubMed]
- Emanuelson, U.; Scherling, K.; Pettersson, H. Relationships between herd bovine leukemia virus infection status and reproduction, disease incidence, and productivity in Swedish dairy herds. Prev. Vet. Med. 1992, 12, 121–131. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, W.; Mao, Y.; Yang, Z.; Lu, G.; Zhang, R.; Zhang, H.; Szeto, C.; Wang, C. Bovine leukemia virus infection in cattle of China: Association with reduced milk production and increased somatic cell score. J. Dairy Sci. 2016, 99, 3688–3697. [Google Scholar] [CrossRef] [PubMed]
- Sandev, N.; Koleva, M.; Binev, R.; Ilieva, D. Influence of enzootic bovine leukosis virus upon the incidence of subclinical mastitis in cows at a different stage of infection. Vet. Arhiv. 2004, 74, 411–416. [Google Scholar]
- Murakami, H.; Yamada, T.; Suzuki, M.; Nakahara, Y.; Suzuki, K.; Sentsui, H. Bovine leukemia virus integration site selection in cattle that develop leukemia. Virus Res. 2011, 156, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Rosewick, N.; Durkin, K.; Artesi, M.; Marçais, A.; Hahaut, V.; Griebel, P.; Arsic, N.; Avettand-Fenoel, V.; Burny, A.; Charlier, C.; et al. Cis-perturbation of cancer drivers by the HTLV-1/BLV proviruses is an early determinant of leukemogenesis. Nat. Commun. 2017, 8, 15264. [Google Scholar] [CrossRef] [PubMed]
- Blankenstein, P.; Fechner, H.; Looman, A.C.; Beier, D.; Marquardt, O.; Ebner, D. Polymerase chain reaction (PCR) for detection of BLV provirus—A practical complement for BLV diagnosis? Berl. Munch. Tierarztl. Wochenschr. 1998, 111, 180–186. [Google Scholar] [PubMed]
- Fechner, H.; Kurg, A.; Geue, L.; Blankenstein, P.; Mewes, G.; Ebner, D.; Beier, D. Evaluation of polymerase chain reaction (PCR) application in diagnosis of bovine leukemia virus (BLV) infection in naturally infected cattle. J. Vet. Med. Ser. B 1996, 43, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Juliarena, M.A.; Gutierrez, S.E.; Ceriani, C. Determination of proviral load in bovine leukemia virus-infected cattle with and without lymphocytosis. Am. J. Vet. Res. 2007, 68, 1220–1225. [Google Scholar] [CrossRef] [PubMed]
- Ohno, A.; Takeshima, S.; Matsumoto, Y.; Aida, Y. Risk factors associated with increased bovine leukemia virus proviral load in infected cattle in Japan from 2012 to 2014. Virus Res. 2015, 210, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Mekata, H.; Yamamoto, M.; Hayashi, T.; Kirino, Y.; Sekiguchi, S.; Konnai, S.; Horii, Y.; Norimine, J. Cattle with a low bovine leukemia virus proviral load are rarely an infectious source. Jpn. J. Vet. Res. 2018, 66, 157–163. [Google Scholar] [CrossRef]
- Ruggiero, V.J.; Norby, B.; Benitez, O.J.; Hutchinson, H.; Sporer, K.R.B.; Droscha, C.; Swenson, C.L.; Bartlett, P.C. Controlling bovine leukemia virus in dairy herds by identifying and removing cows with the highest proviral load and lymphocyte counts. J. Dairy Sci. 2019, 102, 9165–9175. [Google Scholar] [CrossRef] [PubMed]
- Axis, T.M.; DeJong, T.N.; Swenson, C.L.; Sporer, K.R.B.; Droscha, C.; Niles, D.; Bartlett, P.C. Reducing bovine leukemia virus prevalence on a large midwestern dairy farm by using lymphocyte counts, ELISA antibody testing, and proviral load. Bovine Pract. 2020, 54, 136–144. [Google Scholar] [CrossRef]
- Shrestha, S.; Orsel, K.B.; Herman, W.; Martins, L.; Shrestha, S.; Van der Meer, F. Effects of bovine leukemia virus seropositivity and proviral load on milk, fat, and protein production of dairy cows. J. Dairy Sci. 2024, 107, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Murakami, H.; Kakinuma, S.; Murao, K.; Ohmae, K.; Isobe, N.; Akamatsu, H.; Seto, T.; Hashimura, S.; Konda, K.; et al. Association between bovine leukemia virus proviral load and severity of clinical mastitis. J. Vet. Med. Sci. 2019, 81, 1431–1437. [Google Scholar] [CrossRef] [PubMed]
- Nakada, S.; Fujimoto, Y.; Kohara, J.; Makita, K. Economic losses associated with mastitis due to bovine leukemia virus infection. J. Dairy Sci. 2023, 106, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Kawai, K.; Iwano, H.; Usui, M.; Shinozuka, Y.; Hayashi, T.; Higuchi, H. Guidebook for Antimicrobial Treatment of Bovine Mastitis; Ministry of Agriculture, Forestry and Fisheries: Tokyo, Japan, 2017. (In Japanese) [Google Scholar]
- Lago, A.; Godden, S.M.; Bey, R.; Ruegg, P.L.; Leslie, K. The selective treatment of clinical mastitis based on on-farm culture results: I. Effects on antibiotic use, milk withholding time, and short-term clinical and bacteriological outcomes. J. Dairy Sci. 2011, 94, 4441–4456. [Google Scholar] [CrossRef] [PubMed]
- Kiku, Y.; Ozawa, T.; Takahashi, H.; Kushibiki, S.; Inumaru, S.; Shingu, H.; Nagasawa, Y.; Watanabe, A.; Hata, E.; Hayashi, T. Effect of intramammary infusion of recombinant bovine GM-CSF and IL-8 on CMT score, somatic cell count, and milk mononuclear cell populations in Holstein cows with Staphylococcus aureus subclinical mastitis. Vet. Res. Commun. 2017, 41, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Sakai, Y.; Matsuda, K.; Ohtsuka, H.; Muramatsu, Y.; Hara, H.; Nakagawa-Yagi, Y.; Hara, T. Effect of Sasa senanensis extract (SanSTAGE) on Holstein cows with chronic mastitis. Jpn. J. Large Anim. Clin. 2012, 3, 174–180. [Google Scholar] [CrossRef]
- Brunner, C.J.; Muscoplat, C.C. Immunomodulatory effects of levamisole. J. Am. Vet. Med. Assoc. 1980, 176, 1159–1162. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.C. Levamisole and bovine mastitis. Vet. Rec. 1984, 114, 138–140. [Google Scholar] [CrossRef] [PubMed]
- Hadden, J.W.; England, A.; Sadlik, J.R.; Hadden, E.M. The comparative effects of isoprinosine, levamisole, muramyl dipeptide and SM1213 on lymphocyte and macrophage proliferation and activation in vitro. Int. J. Immunopharmacol. 1979, 1, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Onodera, T.; Tsukamoto, S.; Kume, T. Efficacy of levamisole treatment of bovine chronic mastitis. J. Jpn. Vet. Med. Assoc. 1980, 33, 375–378. [Google Scholar] [CrossRef]
- Son, B.W.; Kim, C.S.; Joo, L.S. Efficacy of levamisole treatment of bovine chronic mastitis. J. Korean Vet. Med. Assoc. 1983, 19, 39–44. [Google Scholar]
- Ishikawa, H.; Shimizu, T.; Hirano, H.; Saito, N.; Nakano, T. Protein composition of whey from subclinical mastitis and effect of treatment with levamisole. J. Dairy Sci. 1982, 65, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Hakogi, E.; Ishida, S.; Okada, H. Efficiency of levamisole on bovine subclinical mastitis. J. Jpn. Vet. Med. Assoc. 1983, 36, 382–387. [Google Scholar] [CrossRef]
- Yousaf, A.; Sarfaraz, I.; Zafar, M.A.; Abbas, R.Z.; Hussain, A.; Manzoor, A. Effect of treatment with tri-sodium citrate alone and in combination with levamisole HCl on total milk bacterial count in dairy buffalo suffering from sub-clinical mastitis. Rev. Vet. 2010, 21, 187–189. [Google Scholar]
- Lima, E.S.; Blagitz, M.G.; Batista, C.F.; Alves, A.J.; Fernandes, A.C.C.; Sanchez, E.M.R.; Torres, H.F.; Diniz, S.A.; Silva, M.X.; Della Libera, A.M.M.P.; et al. Milk macrophage function in bovine leukemia virus-infected dairy cows. Front. Vet. Sci. 2021, 8, 650021. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Murakami, H.; Kakinuma, S.; Murao, K.; Oomae, K.; Akamatsu, H.; Seto, T.; Shinozuka, Y.; Kawai, K. Predicting an increased risk of severe clinical mastitis and economic loss using a threshold value of bovine leukemia virus proviral load. Am. J. Vet. Res. 2024, 85, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Middleton, J.R.; Fox, L.K.; Pghetti, G.; Petersson-Wolfe, C. Laboratory Handbook on Bovine Mastitis, 3rd ed.; National Mastitis Council Inc.: Madison, WI, USA, 2017; pp. 19–24. [Google Scholar]
- Kawai, K.; Hayashi, T.; Kiku, Y.; Chiba, T.; Nagahata, H.; Higuchi, H.; Obayashi, T.; Itoh, S.; Onda, K.; Arai, S.; et al. Reliability in somatic cell count measurement of clinical mastitis milk using DeLaval cell counter. Anim. Sci. J. 2013, 84, 805–807. [Google Scholar] [CrossRef] [PubMed]
- Kakinuma, S.; Maeda, Y.; Ohtsuka, H.; Ohmae, K.; Ayabe, K.; Konnai, S.; Oikawa, M. The leukocyte population in the peripheral blood and the colostrum of cows infected with bovine leukemia virus is skewed towards humoral immunity. Int. J. Appl. Res. Vet. Med. 2012, 10, 323–327. [Google Scholar]
- Murakami, H.; Uchiyama, J.; Nikaido, S.; Sato, R.; Sakaguchi, M.; Tsukamoto, K. Inefficient viral replication of bovine leukemia virus induced by spontaneous deletion mutation in the G4 gene. J. Gen. Virol. 2016, 97, 2753–2762. [Google Scholar] [CrossRef] [PubMed]
- Murakami, H.; Uchiyama, J.; Suzuki, C.; Nikaido, S.; Shibuya, K.; Sato, R.; Maeda, Y.; Tomioka, M.; Takeshima, S.-N.; Kato, H.; et al. Variations in the viral genome and biological properties of bovine leukemia virus wild-type strains. Virus Res. 2018, 253, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Murakami, H.; Todaka, H.; Uchiyama, J.; Sato, R.; Sogawa, K.; Sakaguchi, M.; Tsukamoto, K. A point mutation to the long terminal repeat of bovine leukemia virus related to viral productivity and transmissibility. Virology 2019, 537, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, J.; Murakami, H.; Sato, R.; Mizukami, K.; Suzuki, T.; Shima, A.; Ishihara, G.; Sogawa, K.; Sakaguchi, M. Examination of the fecal microbiota in dairy cows infected with bovine leukemia virus. Vet. Microbiol. 2020, 240, 108547. [Google Scholar] [CrossRef] [PubMed]
- Osame, S.; Seo, H.; Ichijo, S.; Inada, I.; Eguchi, T.; Sarashina, T. Effects of vitamin A and E on somatic cell counts in milk of dairy cows. J. Jpn. Vet. Med. Assoc. 1990, 43, 453–458. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Qi, G.; Brand, D.; Zheng, S.G. Role of vitamin A in the immune system. J. Clin. Med. 2018, 7, 258. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.D.; Meydani, S.N.; Wu, D. Regulatory role of vitamin E in the immune system and inflammation. IUBMB Life 2019, 71, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Randhawa, S.S.; Singh, R.S. The effect of biotin and zinc supplementation on dairy cow hoof health and milk quality. Vet. Arhiv 2019, 89, 799–820. [Google Scholar] [CrossRef]
- Kuroishi, T. Regulation of immunological and inflammatory functions by biotin. Can. J. Physiol. Pharmacol. 2015, 93, 1091–1096. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, M.; Agrawal, R.; Singh, R.; Pande, N.; Bhavna. Effect of prepartum levamisole therapy on postpartum udder health in dairy cows. Haryana Vet. 2020, 59, 96–98. [Google Scholar]
- Ovadia, H.; Flesh, J.; Nelken, D. Prevention of bovine mastitis by treatment with levamisole. Isr. J. Med. Sci. 1978, 14, 394–396. [Google Scholar] [PubMed]
- Kume, T. Bovine Mastitis: Mammary Gland Infection and Resistance—One Aspect; Kindai Shuppan Co.: Tokyo, Japan, 1984; pp. 53–64. (In Japanese) [Google Scholar]
- Ignarro, L.J. Regulation of Polymorphonuclear Leukocyte, Macrophage, and Platelet Function. In Immunopharmacology (Comprehensive Immunology), 3rd ed.; Hadden, J.W., Coffey, R.G., Spreafico, F., Eds.; Springer: Boston, MA, USA, 1977; Volume 3, pp. 61–86. [Google Scholar] [CrossRef]
- Hernandes, M.R.G.; Moraes, L.C.A.; Ribeiro, E.B.; Fagundes, D.L.G.; Honorio-França, A.C.; França, E.L. In vitro immunomodulatory effects of microemulsions with levamisole delivery systems on blood phagocyte interacting with Giardia lamblia. Parasitol. Int. 2017, 66, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Rainard, P.; Riollet, C. Innate immunity of the bovine mammary gland. Vet. Res. 2006, 37, 369–400. [Google Scholar] [CrossRef] [PubMed]
- Ladera Gómez, M.E.; Nieto Farias, M.V.; Vater, A.; Ceriani, M.C.; Dolcini, G.L. Study of the proviral load levels and mRNA expression of cytokines in peripheral blood mononuclear cells and somatic milk cells in cattle with different BLV infection profiles. Vet. Ital. 2023, 59, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, S. Concept of parasite control throughout the lifetime of cattle. J. Clin. Vet. Med. 2003, 21, 10–12. (In Japanese) [Google Scholar]
- Kordalis, N.G.; Arsenopoulos, K.; Vasileiou, N.G.C.; Papadopoulos, E.; Fthenakis, G.C. Field evidence for association between increased gastrointestinal nematode burden and subclinical mastitis in dairy sheep. Vet. Parasitol. 2019, 265, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Mavrogianni, V.S.; Papadopoulos, E.; Gougoulis, D.A.; Gallidis, E.; Fthenakis, G.C. Gastrointestinal trichostrongylosis can predispose ewes to clinical mastitis after experimental mammary infection. Vet. Parasitol. 2017, 245, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Arsenopoulos, K.; Gelasakis, A.I.; Delistamatis, V.; Papadopoulos, E. Evaluation of the pour-on administration of eprinomectin on milk yield and somatic cell counts in dairy ewes naturally infected with gastrointestinal nematodes. Vet. Parasitol. Open Access 2019, 276S, 100016. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Shimizu, T. Depression of B-lymphocytes by mastitis and treatment with levamisole. J. Dairy Sci. 1983, 66, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Renoux, G.; Kassel, R.L.; Renoux, M.; Fiore, N.C.; Guillamin, J.M.; Palat, A. Immunomodulation by levamisole in normal and leukemia mice: Evidence for a serum factor. In Modulation of Host Immune Resistance in the Prevention or Treatment of Induced Neoplasias; Chirigos, M.A., Ed.; Fogarty International Proceedings No. 28; US Government Printing Office: Washington, DC, USA, 1977; pp. 45–51. [Google Scholar]
- Van Der Maaten, M.J.; Schmerr, M.J.; Miller, J.M.; Sacks, J.M. Levamisole does not affect the virological and serological responses of bovine leukemia virus-infected cattle and sheep. Can. J. Comp. Med. 1983, 47, 474–479. [Google Scholar] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LMS-Treated | LMS-Untreated | ||||
|---|---|---|---|---|---|
| Above-PVL | Below-PVL | Above-PVL | p-Value | ||
| Variables | 17 quarters, 15 cows | 6 quarters, 6 cows | 9 quarters, 9 cows | (1) | (3) |
| Age (year) | 5.5 ± 1.7 | 4.3 ± 1.3 | 4.7 ± 1.3 | 0.347 | 0.154 |
| Parity | 3.3 ± 1.5 | 2.0 ± 0.9 | 2.4 ± 1.5 | 0.138 | 0.055 |
| Days in Milk | 248.3 ± 93.6 | 308.8 ± 69.7 | 252.2 ± 141.7 | 0.953 | 0.267 |
| Pathogen | |||||
| Streptococcus spp. | 2 | 2 | 0 | ||
| Enterococcus spp. | 2 | 3 | 0 | ||
| Coagulase-negative Staphylococcus spp. | 2 | 0 | 2 | ||
| Coliform bacteria | 0 | 0 | 1 | 0.315 | 0.224 |
| Corynebacterium bovis | 1 | 0 | 1 | ||
| Staphylococcus aureus | 0 | 0 | 0 | ||
| No Growth | 9 | 1 | 4 | ||
| Unknown | 1 | 0 | 1 | ||
| LMS-Treated | |||
|---|---|---|---|
| BLV-Positive | BLV-Negative | p-Value | |
| Variables | 23 quarters, 21 cows | 16 quarters, 12 cows | (2) |
| Age (year) | 5.2 ± 1.7 | 5.1 ± 2.1 | 0.699 |
| Parity | 2.9 ± 1.4 | 2.9 ± 1.4 | 0.868 |
| Days in Milk | 265.6 ± 90.2 | 229.0 ± 89.5 | 0.258 |
| Pathogen | |||
| Streptococcus spp. | 2 | 2 | |
| Enterococcus spp. | 5 | 5 | |
| Coagulase-negative Staphylococcus spp. | 2 | 2 | |
| Coliform bacteria | 0 | 0 | 0.780 |
| Corynebacterium bovis | 1 | 0 | |
| Staphylococcus aureus | 0 | 2 | |
| No Growth | 10 | 7 | |
| Unknown | 1 | 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, A.; Maeda, Y.; Murakami, H.; Miyoshi, S.; Miura, M.; Murao, K.; Shinozuka, Y.; Kurumisawa, T.; Kawai, K. Evaluation of the Therapeutic Effect of Levamisole on Subclinical Mastitis in Bovine Leukemia Virus-Infected Cows Classified by Proviral Load. Animals 2025, 15, 2145. https://doi.org/10.3390/ani15142145
Watanabe A, Maeda Y, Murakami H, Miyoshi S, Miura M, Murao K, Shinozuka Y, Kurumisawa T, Kawai K. Evaluation of the Therapeutic Effect of Levamisole on Subclinical Mastitis in Bovine Leukemia Virus-Infected Cows Classified by Proviral Load. Animals. 2025; 15(14):2145. https://doi.org/10.3390/ani15142145
Chicago/Turabian StyleWatanabe, Aiko, Yosuke Maeda, Hironobu Murakami, Shiro Miyoshi, Michisaburo Miura, Koki Murao, Yasunori Shinozuka, Tomomi Kurumisawa, and Kazuhiro Kawai. 2025. "Evaluation of the Therapeutic Effect of Levamisole on Subclinical Mastitis in Bovine Leukemia Virus-Infected Cows Classified by Proviral Load" Animals 15, no. 14: 2145. https://doi.org/10.3390/ani15142145
APA StyleWatanabe, A., Maeda, Y., Murakami, H., Miyoshi, S., Miura, M., Murao, K., Shinozuka, Y., Kurumisawa, T., & Kawai, K. (2025). Evaluation of the Therapeutic Effect of Levamisole on Subclinical Mastitis in Bovine Leukemia Virus-Infected Cows Classified by Proviral Load. Animals, 15(14), 2145. https://doi.org/10.3390/ani15142145