
Improving Farmed Juvenile Gilthead Seabream (Sparus aurata) Stress Response to Marine Heatwaves and Vibriosis Through Seaweed-Based Dietary Modulation

 , , ,
, , ,  , , ,
, , ,  ,
,  ,
,  , and
, and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Fish Rearing and Acclimation Conditions
2.3. Experimental Design
2.4. MHW Modulation
2.5. V. harveyi Challenge
2.6. Sampling
2.7. Growth Performance Calculations
2.8. Plasma Biochemistry Parameters
2.9. Oxidative Stress Response
2.10. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Plasma Biochemistry
3.3. Oxidative Stress
4. Discussion
4.1. Supplementation Under Control Temperature and MHW Conditions
4.2. Combined Effects of MHW and V. harveyi Challenge
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
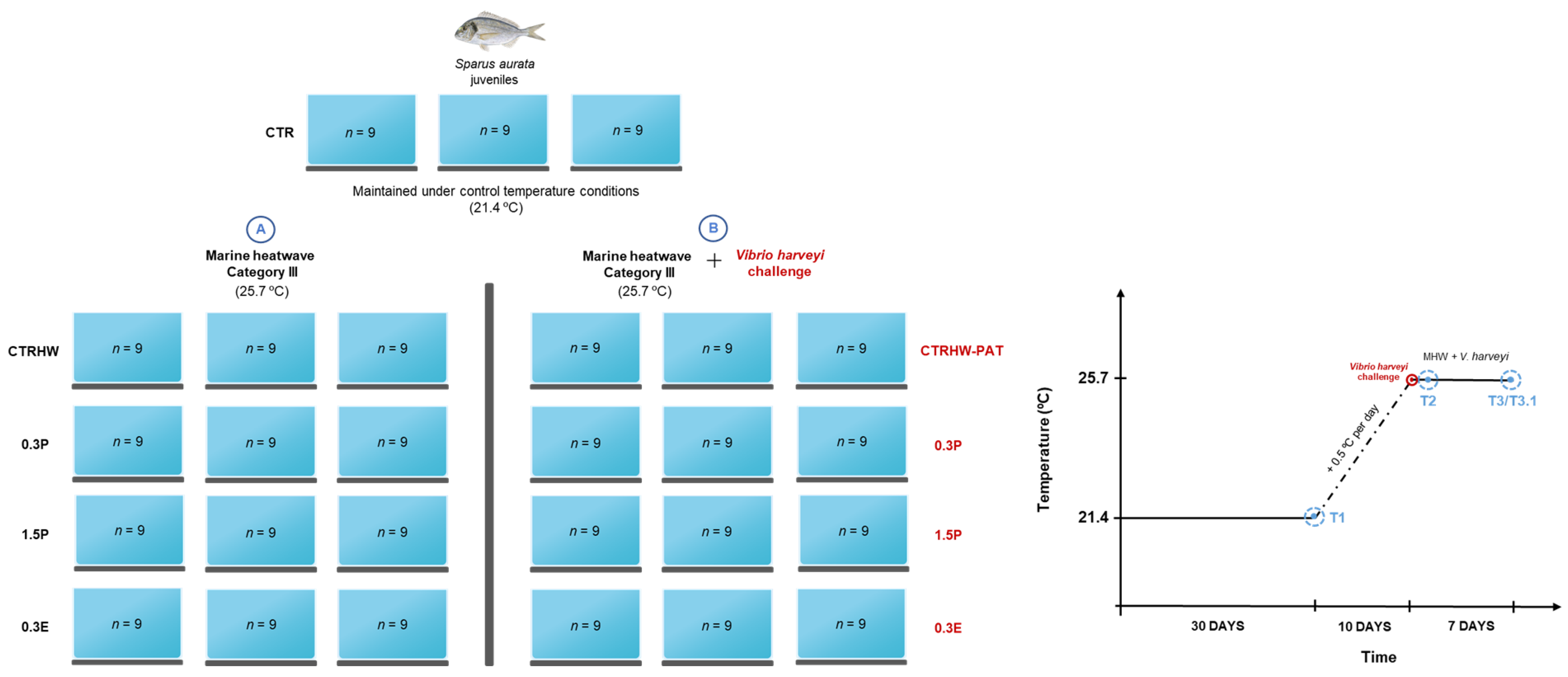
Appendix A. Experimental Design
References
- Passos, R.; Correia, A.P.; Ferreira, I.; Pires, P.; Pires, D.; Gomes, E.; Do Carmo, B.; Santos, P.; Simões, M.; Afonso, C.; et al. Effect on Health Status and Pathogen Resistance of Gilthead Seabream (Sparus aurata) Fed with Diets Supplemented with Gracilaria gracilis. Aquaculture 2021, 531, 735888. [Google Scholar] [CrossRef]
- Bondad-Reantaso, M.G.; MacKinnon, B.; Karunasagar, I.; Fridman, S.; Alday-Sanz, V.; Brun, E.; Le Groumellec, M.; Li, A.; Surachetpong, W.; Karunasagar, I.; et al. Review of Alternatives to Antibiotic Use in Aquaculture. Rev. Aquac. 2023, 15, 1421–1451. [Google Scholar] [CrossRef]
- Reverter, M.; Sarter, S.; Caruso, D.; Avarre, J.-C.; Combe, M.; Pepey, E.; Pouyaud, L.; Vega-Heredía, S.; De Verdal, H.; Gozlan, R.E. Aquaculture at the Crossroads of Global Warming and Antimicrobial Resistance. Nat. Commun. 2020, 11, 1870. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Esteban, M.Á. Beneficial Roles of Feed Additives as Immunostimulants in Aquaculture: A Review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Okocha, R.C.; Olatoye, I.O.; Adedeji, O.B. Food Safety Impacts of Antimicrobial Use and Their Residues in Aquaculture. Public. Health Rev. 2018, 39, 21. [Google Scholar] [CrossRef]
- Farías, D.R.; Ibarra, R.; Estévez, R.A.; Tlusty, M.F.; Nyberg, O.; Troell, M.; Avendaño-Herrera, R.; Norden, W. Towards Sustainable Antibiotic Use in Aquaculture and Antimicrobial Resistance: Participatory Experts’ Overview and Recommendations. Antibiotics 2024, 13, 887. [Google Scholar] [CrossRef]
- Frölicher, T.L.; Fischer, E.M.; Gruber, N. Marine Heatwaves under Global Warming. Nature 2018, 560, 360–364. [Google Scholar] [CrossRef]
- Oliver, E.C.J.; Burrows, M.T.; Donat, M.G.; Sen Gupta, A.; Alexander, L.V.; Perkins-Kirkpatrick, S.E.; Benthuysen, J.A.; Hobday, A.J.; Holbrook, N.J.; Moore, P.J.; et al. Projected Marine Heatwaves in the 21st Century and the Potential for Ecological Impact. Front. Mar. Sci. 2019, 6, 734. [Google Scholar] [CrossRef]
- Marques, A.; Marçal, R.; Pereira, V.; Pereira, P.; Mieiro, C.; Guilherme, S.; Marques, C.; Santos, M.A.; Pereira, R.; Abreu, H.; et al. Macroalgae-Enriched Diet Protects Gilthead Seabream (Sparus aurata) against Erythrocyte Population Instability and Chromosomal Damage Induced by Aqua-Medicines. J. Appl. Phycol. 2020, 32, 1477–1493. [Google Scholar] [CrossRef]
- Rosa, R.; Marques, A.; Nunes, M.L. Impact of Climate Change in Mediterranean Aquaculture. Rev. Aquac. 2012, 4, 163–177. [Google Scholar] [CrossRef]
- Cascarano, M.C.; Stavrakidis-Zachou, O.; Mladineo, I.; Thompson, K.D.; Papandroulakis, N.; Katharios, P. Mediterranean Aquaculture in a Changing Climate: Temperature Effects on Pathogens and Diseases of Three Farmed Fish Species. Pathogens 2021, 10, 1205. [Google Scholar] [CrossRef]
- Austin, B.; Austin, D.A. Vibrionaceae Representatives. In Bacterial Fish Pathogens: Disease of Farmed and Wild Fish; Springer: Dordrecht, The Netherlands, 2012; pp. 357–411. [Google Scholar]
- Gauger, E.; Gómez-Chiarri, M. 16S Ribosomal DNA Sequencing Confirms the Synonymy of Vibrio harveyi and V. carchariae. Dis. Aquat. Org. 2002, 52, 39–46. [Google Scholar] [CrossRef]
- Cano-Gomez, A.; Bourne, D.G.; Hall, M.R.; Owens, L.; Høj, L. Molecular Identification, Typing and Tracking of Vibrio harveyi in Aquaculture Systems: Current Methods and Future Prospects. Aquaculture 2009, 287, 1–10. [Google Scholar] [CrossRef]
- Austin, B.; Zhang, X.-H. Vibrio Harveyi: A Significant Pathogen of Marine Vertebrates and Invertebrates. Lett. Appl. Microbiol. 2006, 43, 119–124. [Google Scholar] [CrossRef]
- Arias, C.R.; Macian, M.C.; Aznar, R.; Garay, E.; Pujalte, M.J. Low Incidence of Vibrio vulnificus among Vibrio Isolates from Sea Water and Shellfish of the Western Mediterranean Coast. J. Appl. Microbiol. 1999, 86, 125–134. [Google Scholar] [CrossRef]
- Pujalte, M.J.; Ortigosa, M.; Macián, M.C.; Garay, E. Aerobic and Facultative Anaerobic Heterotrophic Bacteria Associated to Mediterranean Oysters and Seawater. Int. Microbiol. 1999, 2, 259–266. [Google Scholar]
- Pujalte, M.; Sitjà-Bobadilla, A.; Álvarez-Pellitero, P.; Garay, E. Carriage of Potentially Fish-Pathogenic Bacteria in Sparus aurata Cultured in Mediterranean Fish Farms. Dis. Aquat. Org. 2003, 54, 119–126. [Google Scholar] [CrossRef]
- Oliva-Teles, A. Nutrition and Health of Aquaculture Fish. J. Fish. Dis. 2012, 35, 83–108. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Yousefi, S.; Capillo, G.; Paknejad, H.; Khalili, M.; Tabarraei, A.; Van Doan, H.; Spanò, N.; Faggio, C. Mucosal Immune Parameters, Immune and Antioxidant Defence Related Genes Expression and Growth Performance of Zebrafish (Danio rerio) Fed on Gracilaria gracilis Powder. Fish. Shellfish. Immunol. 2018, 83, 232–237. [Google Scholar] [CrossRef]
- Hassaan, M.S.; El Nagar, A.G.; Salim, H.S.; Fitzsimmons, K.; El-Haroun, E.R. Nutritional Mitigation of Winter Thermal Stress in Nile Tilapia by Propolis-Extract: Associated Indicators of Nutritional Status, Physiological Responses and Transcriptional Response of Delta-9-Desaturase Gene. Aquaculture 2019, 511, 734256. [Google Scholar] [CrossRef]
- Turchini, G.M.; Nie, P. The Climate Is Still Changing. Rev. Aquac. 2021, 13, 3–4. [Google Scholar] [CrossRef]
- Costa, M.; Cardoso, C.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Current Knowledge and Future Perspectives of the Use of Seaweeds for Livestock Production and Meat Quality: A Systematic Review. Anim. Physiol. Nutr. 2021, 105, 1075–1102. [Google Scholar] [CrossRef]
- Marmelo, I.; Dias, M.; Grade, A.; Pousão-Ferreira, P.; Diniz, M.S.; Marques, A.; Maulvault, A.L. Immunomodulatory and Antioxidant Effects of Functional Aquafeeds Biofortified with Whole Laminaria digitata in Juvenile Gilthead Seabream (Sparus aurata). Front. Mar. Sci. 2024, 11, 1325244. [Google Scholar] [CrossRef]
- Morais, T.; Inácio, A.; Coutinho, T.; Ministro, M.; Cotas, J.; Pereira, L.; Bahcevandziev, K. Seaweed Potential in the Animal Feed: A Review. J. Mar. Sci. Eng. 2020, 8, 559. [Google Scholar] [CrossRef]
- Silva, A.; Silva, S.A.; Carpena, M.; Garcia-Oliveira, P.; Gullón, P.; Barroso, M.F.; Prieto, M.A.; Simal-Gandara, J. Macroalgae as a Source of Valuable Antimicrobial Compounds: Extraction and Applications. Antibiotics 2020, 9, 642. [Google Scholar] [CrossRef]
- Marmelo, I.; Lourenço-Marques, C.; Silva, I.A.L.; Soares, F.; Pousão-Ferreira, P.; Mata, L.; Marques, A.; Diniz, M.S.; Maulvault, A.L. Eco-Innovative Aquafeeds Biofortified with Asparagopsis taxiformis to Improve the Resilience of Farmed White Seabream (Diplodus sargus) to Marine Heatwave Events. Heliyon 2024, 10, e35135. [Google Scholar] [CrossRef]
- Pereira, A.; Marmelo, I.; Dias, M.; Anacleto, P.; Pires, C.; Batista, I.; Marques, A.; Maulvault, A.L. Antioxidant, Metabolic and Digestive Biomarker Responses of Farmed Sparus aurata Supplemented with Laminaria digitata. Aquaculture 2025, 598, 741984. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, Structure and Biofunctional Activities of Laminarin from Brown Algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Ramalho Ribeiro, A.; Gonçalves, A.; Colen, R.; Nunes, M.L.; Dinis, M.T.; Dias, J. Dietary Macroalgae Is a Natural and Effective Tool to Fortify Gilthead Seabream Fillets with Iodine: Effects on Growth, Sensory Quality and Nutritional Value. Aquaculture 2015, 437, 51–59. [Google Scholar] [CrossRef]
- Palstra, A.P.; Kals, J.; Blanco Garcia, A.; Dirks, R.P.; Poelman, M. Immunomodulatory Effects of Dietary Seaweeds in LPS Challenged Atlantic Salmon Salmo salar as Determined by Deep RNA Sequencing of the Head Kidney Transcriptome. Front. Physiol. 2018, 9, 625. [Google Scholar] [CrossRef]
- Anacleto, P.; Figueiredo, C.; Baptista, M.; Maulvault, A.L.; Camacho, C.; Pousão-Ferreira, P.; Valente, L.M.P.; Marques, A.; Rosa, R. Fish Energy Budget under Ocean Warming and Flame Retardant Exposure. Environ. Res. 2018, 164, 186–196. [Google Scholar] [CrossRef]
- FAO Fisheries and Aquaculture Department. Species Fact Sheets: Sparus aurata. 2023. Available online: https://www.fao.org/fishery/en/aqspecies/2384/en (accessed on 24 October 2024).
- Pereira, A.; Marmelo, I.; Dias, M.; Silva, A.C.; Grade, A.C.; Barata, M.; Pousão-Ferreira, P.; Dias, J.; Anacleto, P.; Marques, A.; et al. Asparagopsis taxiformis as a Novel Antioxidant Ingredient for Climate-Smart Aquaculture: Antioxidant, Metabolic and Digestive Modulation in Juvenile White Seabream (Diplodus sargus) Exposed to a Marine Heatwave. Antioxidants 2024, 13, 949. [Google Scholar] [CrossRef]
- Schlegel, W.R.; Smit, J.A. heatwaveR: A Central Algorithm for the Detection of Heatwaves and Cold-Spells. JOSS 2018, 3, 821. [Google Scholar] [CrossRef]
- Hobday, A.J.; Oliver, E.C.J.; Gupta, A.S.; Benthuysen, J.A.; Burrows, M.T.; Donat, M.G.; Holbrook, N.J.; Moore, P.J.; Thomsen, M.S.; Wernberg, T.; et al. Categorizing and Naming Marine Heatwaves. Oceanography 2018, 31, 162–173. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Naiel, M.A.E.; Alagawany, M.; Patra, A.K.; El-Kholy, A.I.; Amer, M.S.; Abd El-Hack, M.E. Beneficial Impacts and Health Benefits of Macroalgae Phenolic Molecules on Fish Production. Aquaculture 2021, 534, 736186. [Google Scholar] [CrossRef]
- Kamunde, C.; Sappal, R.; Melegy, T.M. Brown Seaweed (AquaArom) Supplementation Increases Food Intake and Improves Growth, Antioxidant Status and Resistance to Temperature Stress in Atlantic Salmon, Salmo salar. PLoS ONE 2019, 14, e0219792. [Google Scholar] [CrossRef]
- Vizcaíno, A.J.; Mendes, S.I.; Varela, J.L.; Ruiz-Jarabo, I.; Rico, R.; Figueroa, F.L.; Abdala, R.; Moriñigo, M.Á.; Mancera, J.M.; Alarcón, F.J. Growth, Tissue Metabolites and Digestive Functionality in Sparus aurata Juveniles Fed Different Levels of Macroalgae, Gracilaria cornea and Ulva rigida. Aquac. Res. 2016, 47, 3224–3238. [Google Scholar] [CrossRef]
- Maulvault, A.L.; Barbosa, V.; Alves, R.; Anacleto, P.; Camacho, C.; Cunha, S.; Fernandes, J.O.; Ferreira, P.P.; Rosa, R.; Marques, A.; et al. Integrated Multi-Biomarker Responses of Juvenile Seabass to Diclofenac, Warming and Acidification Co-Exposure. Aquat. Toxicol. 2018, 202, 65–79. [Google Scholar] [CrossRef]
- Yuan, D.; Wang, H.; Liu, X.; Wang, S.; Shi, J.; Cheng, X.; Gu, H.; Xiao, S.; Wang, Z. High Temperature Induced Metabolic Reprogramming and Lipid Remodeling in a High-Altitude Fish Species, Triplophysa Bleekeri. Front. Mar. Sci. 2022, 9, 1017142. [Google Scholar] [CrossRef]
- Peres, H.; Santos, S.; Oliva-Teles, A. Blood Chemistry Profile as Indicator of Nutritional Status in European Seabass (Dicentrarchus labrax). Fish. Physiol. Biochem. 2014, 40, 1339–1347. [Google Scholar] [CrossRef]
- Shahjahan, M.; Islam, M.J.; Hossain, M.T.; Mishu, M.A.; Hasan, J.; Brown, C. Blood Biomarkers as Diagnostic Tools: An Overview of Climate-Driven Stress Responses in Fish. Sci. Total Environ. 2022, 843, 156910. [Google Scholar] [CrossRef]
- Ramesh, M.; Thilagavathi, T.; Rathika, R.; Poopal, R.K. Antioxidant Status, Biochemical, and Hematological Responses in a Cultivable Fish Cirrhinus mrigala Exposed to an Aquaculture Antibiotic Sulfamethazine. Aquaculture 2018, 491, 10–19. [Google Scholar] [CrossRef]
- Islam, M.J.; Puebla, O.; Kunzmann, A. Nutritional Mitigation of Heatwave Stress in European Seabass, Dicentrarchus labrax: Metabolic, Cellular, and Molecular Responses. Aquac. Rep. 2024, 36, 102168. [Google Scholar] [CrossRef]
- Nemova, N.N.; Lysenko, L.A.; Kantserova, N.P. Degradation of Skeletal Muscle Protein during Growth and Development of Salmonid Fish. Russ. J. Dev. Biol. 2016, 47, 161–172. [Google Scholar] [CrossRef]
- Chadwick, J.G.; McCormick, S.D. Upper Thermal Limits of Growth in Brook Trout and Their Relationship to Stress Physiology. J. Exp. Biol. 2017, 220, 3976–3987. [Google Scholar] [CrossRef]
- Pérez-Sánchez, J.; Terova, G.; Simó-Mirabet, P.; Rimoldi, S.; Folkedal, O.; Calduch-Giner, J.A.; Olsen, R.E.; Sitjà-Bobadilla, A. Skin Mucus of Gilthead Sea Bream (Sparus aurata L.). Protein Mapping and Regulation in Chronically Stressed Fish. Front. Physiol. 2017, 8, 34. [Google Scholar] [CrossRef]
- Tan, X.; Lin, H.; Huang, Z.; Zhou, C.; Wang, A.; Qi, C.; Zhao, S. Effects of Dietary Leucine on Growth Performance, Feed Utilization, Non-Specific Immune Responses and Gut Morphology of Juvenile Golden Pompano Trachinotus ovatus. Aquaculture 2016, 465, 100–107. [Google Scholar] [CrossRef]
- Tao, Y.-F.; Qiang, J.; Bao, J.-W.; Chen, D.-J.; Yin, G.-J.; Xu, P.; Zhu, H.-J. Changes in Physiological Parameters, Lipid Metabolism, and Expression of MicroRNAs in Genetically Improved Farmed Tilapia (Oreochromis niloticus) With Fatty Liver Induced by a High-Fat Diet. Front. Physiol. 2018, 9, 1521. [Google Scholar] [CrossRef]
- Panigrahi, A.; Kiron, V.; Satoh, S.; Watanabe, T. Probiotic Bacteria Lactobacillus Rhamnosus Influences the Blood Profile in Rainbow Trout Oncorhynchus mykiss (Walbaum). Fish. Physiol. Biochem. 2010, 36, 969–977. [Google Scholar] [CrossRef]
- Harikrishnan, R.; Kim, M.-C.; Kim, J.-S.; Balasundaram, C.; Heo, M.-S. Probiotics and Herbal Mixtures Enhance the Growth, Blood Constituents, and Nonspecific Immune Response in Paralichthys olivaceus against Streptococcus parauberis. Fish. Shellfish. Immunol. 2011, 31, 310–317. [Google Scholar] [CrossRef]
- Barton, B.A. Stress in Fishes: A Diversity of Responses with Particular Reference to Changes in Circulating Corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef]
- Refaey, M.M.; Li, D.; Tian, X.; Zhang, Z.; Zhang, X.; Li, L.; Tang, R. High Stocking Density Alters Growth Performance, Blood Biochemistry, Intestinal Histology, and Muscle Quality of Channel Catfish Ictalurus punctatus. Aquaculture 2018, 492, 73–81. [Google Scholar] [CrossRef]
- Alfons, M.S.; Ibrahim, A.T.A.; Harabawy, A.S.A.; Al-salahy, M.B.; Badr, G. Cytoprotective Effect of Propolis on Heat Stress Induces Alteration to Histological, Ultrastructural, and Oxidative Stress in Catfish (Clarias gariepinus). Environ. Sci. Pollut. Res. 2023, 30, 114152–114165. [Google Scholar] [CrossRef]
- Gomez Isaza, D.F.; Cramp, R.L.; Smullen, R.; Glencross, B.D.; Franklin, C.E. Coping with Climatic Extremes: Dietary Fat Content Decreased the Thermal Resilience of Barramundi (Lates calcarifer). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 230, 64–70. [Google Scholar] [CrossRef]
- Generalić Mekinić, I.; Šimat, V.; Botić, V.; Crnjac, A.; Smoljo, M.; Soldo, B.; Ljubenkov, I.; Čagalj, M.; Skroza, D. Bioactive Phenolic Metabolites from Adriatic Brown Algae Dictyota Dichotoma and Padina Pavonica (Dictyotaceae). Foods 2021, 10, 1187. [Google Scholar] [CrossRef]
- Galindo, A.; Pérez, J.A.; Martín, V.; Acosta, N.G.; Reis, D.B.; Jiménez, I.A.; Rosa, G.; Venuleo, M.; Marrero, M.; Rodríguez, C. Effect of Feed Supplementation with Seaweed Wracks on Performance, Muscle Lipid Composition, Antioxidant Status, Digestive Enzyme Activities, and Plasma Biochemistry of Gilthead Seabream (Sparus aurata) Juveniles. Aquac. Rep. 2023, 31, 101673. [Google Scholar] [CrossRef]
- De Mello, P.H.; Divanach, P.; Papadakis, I.E. Temperature Influences Growth, Digestive System Ontogeny and Lipids Deposition in the Liver in Gilthead Seabream (Sparus aurata) Larvae and Juveniles. Aquac. Res. 2022, 53, 3416–3429. [Google Scholar] [CrossRef]
- Ziskowski, J.; Mercaldo-Allen, R.; Pereira, J.J.; Kuropat, C.; Goldberg, R. The Effects of Fin Rot Disease and Sampling Method on Blood Chemistry and Hematocrit Measurements of Winter Flounder, Pseudopleuronectes Americanus from New Haven Harbor (1987–1990). Mar. Pollut. Bull. 2008, 56, 740–750. [Google Scholar] [CrossRef]
- Castro, R.; Zarra, I.; Lamas, J. Water-Soluble Seaweed Extracts Modulate the Respiratory Burst Activity of Turbot Phagocytes. Aquaculture 2004, 229, 67–78. [Google Scholar] [CrossRef]
- Marcogliese, D.J.; Brambilla, L.G.; Gagné, F.; Gendron, A.D. Joint Effects of Parasitism and Pollution on Oxidative Stress Biomarkers in Yellow Perch Perca flavescens. Dis. Aquat. Org. 2005, 63, 77–84. [Google Scholar] [CrossRef]
- Peixoto, M.J.; Ferraz, R.; Magnoni, L.J.; Pereira, R.; Gonçalves, J.F.; Calduch-Giner, J.; Pérez-Sánchez, J.; Ozório, R.O.A. Protective Effects of Seaweed Supplemented Diet on Antioxidant and Immune Responses in European Seabass (Dicentrarchus labrax) Subjected to Bacterial Infection. Sci. Rep. 2019, 9, 16134. [Google Scholar] [CrossRef]
- Thanigaivel, S.; Chandrasekaran, N.; Mukherjee, A.; Thomas, J. Protective Efficacy of Microencapsulated Seaweed Extracts for Preventing Aeromonas Infections in Oreochromis mossambicus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 218, 36–45. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Experimental Diets | |||
|---|---|---|---|---|
| CTR | 0.3P | 1.5P | 0.3E | |
| Fishmeal Super Prime a | 17.0 | 17.0 | 17.0 | 17.0 |
| Fish protein hydrolysate b | 2.0 | 2.0 | 2.0 | 2.0 |
| Poultry meal c | 7.0 | 7.0 | 7.0 | 7.0 |
| Soy protein concentrate d | 6.0 | 6.0 | 6.0 | 6.0 |
| Wheat gluten e | 10.0 | 10.0 | 10.0 | 10.0 |
| Corn gluten meal f | 11.0 | 11.0 | 11.0 | 11.0 |
| Soybean meal (Hipro) g | 12.8 | 12.8 | 12.8 | 12.8 |
| Wheat meal h | 12.0 | 11.7 | 10.5 | 11.7 |
| Whole peas i | 6.0 | 6.0 | 6.0 | 6.0 |
| Vitamin and mineral premix j | 1.0 | 1.0 | 1.0 | 1.0 |
| Choline chloride 50% k | 0.2 | 0.2 | 0.2 | 0.2 |
| Monoammonium phosphate l | 1.0 | 1.0 | 1.0 | 1.0 |
| Fish oil m | 6.0 | 6.0 | 6.0 | 6.0 |
| Salmon oil n | 8.0 | 8.0 | 8.0 | 8.0 |
| Laminaria digitata powder o | - | 0.3 | 1.5 | - |
| Laminaria digitata extract p | - | - | - | 0.3 |
| Proximate composition (%) | ||||
| Crude protein | 46.0 | 46.0 | 45.9 | 46.0 |
| Crude fat | 16.0 | 16.0 | 16.0 | 16.0 |
| Fiber | 1.4 | 1.4 | 1.4 | 1.4 |
| Starch | 13.5 | 13.3 | 12.5 | 13.3 |
| Ash | 7.4 | 7.4 | 7.7 | 7.4 |
| Gross energy (MJ/kg feed) | 20.5 | 20.5 | 20.4 | 20.5 |
| Time | TL (cm) | W (g) | K | HSI (%) | SGR (% day −1) | FCR | ||
|---|---|---|---|---|---|---|---|---|
| Supplementation + MHW conditions | CTR | T1 | 14.21 ± 0.64 | 47.21 ± 3.80 | 1.61 ± 0.10 | 1.11 ± 0.16 a | 1.42 ± 0.14 b | 0.38 ± 0.03 * |
| 0.3P | T1 | 14.38 ± 1.05 | 52.83 ± 6.82 | 1.61 ± 0.13 | 0.84 ± 0.08 c | 2.07 ± 0.31 a* | 0.34 ± 0.05 * | |
| 1.5P | T1 | 14.42 ± 0.86 | 48.13 ± 2.74 * | 1.61 ± 0.15 | 0.86 ± 0.10 bc | 1.59 ± 0.13 b | 0.37 ± 0.02 * | |
| 0.3E | T1 | 14.64 ± 0.75 | 53.70 ± 6.97 | 1.66 ± 0.22 | 1.01 ± 0.12 ab* | 2.10 ± 0.36 a* | 0.34 ± 0.04 * | |
| CTR | T3 | 14.73 ± 0.61 | 53.78 ± 4.73 | 1.68 ± 0.32 | 1.10 ± 0.18 a | 1.13 ± 0.13 b | 0.51 ± 0.04 ab# | |
| CTRHW | T3 | 15.38 ± 0.81 | 57.35 ± 5.41 | 1.50 ± 0.11 | 0.72 ± 0.05 b | 1.09 ± 0.16 b | 0.49 ± 0.05 ab# | |
| 0.3P | T3 | 15.26 ± 0.63 | 52.22 ± 2.90 | 1.53 ± 0.10 | 0.62 ± 0.04 b | 1.12 ± 0.09 b# | 0.55 ± 0.03 a# | |
| 1.5P | T3 | 15.72 ± 0.50 | 58.61 ± 3.87 # | 1.49 ± 0.17 | 0.74 ± 0.05 b | 1.45 ± 0.06 a | 0.46 ± 0.02 b# | |
| 0.3E | T3 | 15.29 ± 0.58 | 54.21 ± 4.61 | 1.50 ± 0.16 | 0.63 ± 0.08 b# | 1.12 ± 0.14 b# | 0.52 ± 0.04 a# | |
| MHW conditions + V. harveyi challenge | CTRHW | T2 | 14.96 ± 0.68 | 48.52 ± 2.62 b$ | 1.48 ± 0.09 | 0.78 ± 0.06 b | 1.09 ± 0.09 b | 0.52 ± 0.03 a |
| CTRHW-PAT | T2 | 15.56 ± 0.73 | 58.24 ± 7.37 ab | 1.58 ± 0.12 | 1.15 ± 0.08 a | 1.91 ± 0.23 a$ | 0.41 ± 0.15 ab | |
| 0.3P | T2 | 15.98 ± 0.73 | 58.29 ± 4.43 a | 1.52 ± 0.07 $ | 1.11 ± 0.11 a | 1.66 ± 0.15 a$ | 0.41 ± 0.04 b$ | |
| 1.5P | T2 | 15.48 ± 0.81 | 55.92 ± 4.68 a | 1.61 ± 0.15 | 1.14 ± 0.08 a$ | 1.65 ± 0.18 a | 0.43 ± 0.04 b | |
| 0.3E | T2 | 15.98 ± 0.71 | 61.42 ± 3.14 a | 1.51 ± 0.10 | 1.08 ± 0.10 a | 1.78 ± 0.09 a$ | 0.41 ± 0.02 b | |
| CTRHW | T3.1 | 15.38 ± 0.81 | 57.35 ± 5.41 / | 1.50 ± 0.11 ab | 0.72 ± 0.05 c | 1.09 ± 0.16 c | 0.49 ± 0.05 ab | |
| CTRHW-PAT | T3.1 | 15.57 ± 0.59 | 56.28 ± 4.95 | 1.57 ± 0.12 a | 1.30 ± 0.18 a | 1.27 ± 0.09 bc/ | 0.50 ± 0.04 ab | |
| 0.3P | T3.1 | 16.02 ± 0.55 | 54.37 ± 4.74 | 1.35 ± 0.06 b/ | 0.93 ± 0.09 b | 1.15 ± 0.14 c/ | 0.53 ± 0.04 a/ | |
| 1.5P | T3.1 | 16.04 ± 0.48 | 61.39 ± 3.13 | 1.52 ± 0.13 ab | 0.92 ± 0.08 b/ | 1.54 ± 0.08 a | 0.46 ± 0.02 b | |
| 0.3E | T3.1 | 15.59 ± 0.51 | 57.46 ± 4.19 | 1.55 ± 0.11 a | 0.98 ± 0.08 b | 1.39 ± 0.09 ab/ | 0.47 ± 0.04 ab |
| Time | GLU (mg dL−1) | TP (g dL−1) | BUN (mg dL−1) | PHOS (mg dL−1) | ALT (U L−1) | CA (mg dL−1) | GLOB (g dL−1) | ALP (U L−1) | CHOL (mg dL−1) | AMY (U L−1) | Cortisol (ng mL−1) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Supplementation + MHW conditions | CTR | T1 | 80.33 ± 4.04 b | 3.15 ± 0.21 a | 7.73 ± 1.70 | 7.73 ± 0.64 | 22.00 ± 2.65 a* | 11.93 ± 0.35 b | 2.62 ± 0.24 | 233.67 ± 12.70 a | 164.33 ± 7.57 | 36.00 ± 9.42 | 46.84 ± 1.36 |
| 0.3P | T1 | 110.67 ± 5.69 a* | 3.13 ± 0.34 a | 9.08 ± 1.72 | 9.06 ± 0.59 | 15.33 ± 0.58 b | 15.35 ± 0.72 a* | 2.48 ± 0.33 | 234.33 ± 6.81 a | 170.75 ± 39.85 | 25.75 ± 2.75 | 43.54 ± 5.21 * | |
| 1.5P | T1 | 121.33 ± 7.23 a | 2.53 ± 0.05 b | 8.13 ± 1.65 | 7.92 ± 0.60 | 14.33 ± 1.53 b | 12.33 ± 0.28 b* | 2.20 ± 0.10 | 205.00 ± 9.54 b* | 148.50 ± 8.10 | 27.00 ± 4.08 | 52.40 ± 8.18 | |
| 0.3E | T1 | 121.00 ± 12.29 a | 2.55 ± 0.29 ab* | 8.13 ± 0.92 | 7.46 ± 0.77 | 19.00 ± 3.46 ab* | 11.38 ± 0.34 b | 2.24 ± 0.34 | 191.00 ± 7.00 b | 136.25 ± 32.57 | 25.00 ± 3.37 | 43.78 ± 5.48 * | |
| CTR | T3 | 94.67 ± 4.62 b | 2.85 ± 0.37 ab | 6.55 ± 1.06 ab | 6.74 ± 0.89 | 11.00 ± 1.00 ab# | 12.65 ± 0.87 ab | 2.26 ± 0.23 | 197.33 ± 21.83 b | 176.75 ± 18.82 ab | 32.25 ± 7.37 | 56.38 ± 2.60 b | |
| CTRHW | T3 | 78.67 ± 9.87 b | 2.60 ± 0.24 b | 5.88 ± 0.57 b | 5.90 ± 0.45 | 10.33 ± 0.58 b | 11.10 ± 0.74 b | 2.34 ± 0.26 | 197.00 ± 6.24 b | 150.00 ± 13.29 b | 24.25 ± 5.56 | 61.56 ± 15.36 ab | |
| 0.3P | T3 | 130.67 ± 2.52 a# | 3.28 ± 0.05 a | 7.20 ± 0.29 a | 7.30 ± 0.70 | 14.00 ± 1.73 a | 13.85 ± 0.33 a# | 2.50 ± 0.14 | 222.00 ± 13.89 ab | 180.50 ± 21.46 ab | 19.75 ± 6.50 | 76.88 ± 6.77 a# | |
| 1.5P | T3 | 176.33 ± 41.19 a | 2.98 ± 0.22 ab | 8.33 ± 1.43 a | 7.44 ± 1.05 | 14.67 ± 3.21 a | 13.48 ± 0.71 a# | 2.32 ± 0.23 | 255.00 ± 16.09 a# | 175.75 ± 17.37 ab | 22.25 ± 6.65 | 62.62 ± 6.28 ab | |
| 0.3E | T3 | 133.33 ± 9.02 a | 3.23 ± 0.34 ab# | 6.28 ± 1.63 ab | 7.18 ± 1.46 | 10.17 ± 0.12 b# | 13.08 ± 0.97 a | 2.34 ± 0.13 | 225.67 ± 5.03 b | 201.00 ± 9.83 a | 20.50 ± 3.87 | 64.81 ± 6.71 ab# | |
| MHW conditions + V. harveyi challenge | CTRHW | T2 | 103.33 ± 13.01 | 2.75 ± 0.13 | 9.78 ± 1.60 a$ | 7.28 ± 0.70 b | 22.33 ± 3.06 c | 11.13 ± 0.84 b | 2.44 ± 0.32 | 250.00 ± 18.19 b$ | 128.50 ± 21.76 | 34.25 ± 9.32 | 55.06 ± 5.21 c |
| CTRHW-PAT | T2 | 84.33 ± 12.86 $ | 3.05 ± 0.13 | 5.53 ± 0.64 bc | 11.68 ± 2.35 a$ | 288.00 ± 30.81 a$ | 11.13 ± 0.79 b | 2.24 ± 0.18 | 368.00 ± 44.00 a$ | 120.25 ± 10.50 | 28.50 ± 4.80 | 92.68 ± 2.87 a | |
| 0.3P | T2 | 89.00 ± 3.46 | 3.28 ± 0.10 | 5.48 ± 0.56 bc | 10.78 ± 2.76 a | 367.00 ± 50.74 a$ | 12.45 ± 0.66 ab | 2.20 ± 0.16 | 330.67 ± 28.54 a | 141.25 ± 10.11 | 22.25 ± 5.91 | 91.00 ± 6.50 a$ | |
| 1.5P | T2 | 96.67 ± 4.04 | 2.98 ± 0.26 | 5.03 ± 0.05 c$ | 10.06 ± 1.71 a | 172.67 ± 23.18 b$ | 12.30 ± 0.42 b | 1.88 ± 0.16 | 338.00 ± 31.58 a$ | 138.50 ± 15.78 | 31.25 ± 1.26 | 79.51 ± 2.57 b$ | |
| 0.3E | T2 | 96.67 ± 7.02 $ | 3.30 ± 0.42 | 6.30 ± 0.73 b | 11.80 ± 2.16 a$ | 198.33 ± 45.80 b$ | 14.28 ± 0.85 a | 1.94 ± 0.27 | 306.00 ± 41.07 a | 110.50 ± 23.59 | 25.00 ± 4.55 | 53.95 ± 1.43 c | |
| CTRHW | T3.1 | 78.67 ± 9.87 c | 2.60 ± 0.24 | 5.88 ± 0.57 ab/ | 5.90 ± 0.45 b | 10.33 ± 0.58 b | 11.10 ± 0.74 b | 2.34 ± 0.26 | 197.00 ± 6.24 c/ | 150.00 ± 13.29 a | 24.25 ± 5.56 ab | 61.56 ± 15.36 ab | |
| CTRHW-PAT | T3.1 | 133.33 ± 3.79 a/ | 2.58 ± 0.44 | 5.95 ± 0.91 ab | 7.54 ± 0.88 a/ | 10.03 ± 0.05 b/ | 12.43 ± 0.95 ab | 1.80 ± 0.24 | 222.33 ± 4.16 b/ | 119.25 ± 20.25 ab | 18.75 ± 4.86 b | 80.60 ± 10.91 a | |
| 0.3P | T3.1 | 105.33 ± 4.51 b | 2.93 ± 0.54 | 5.65 ± 0.79 ab | 7.48 ± 0.91 a | 40.33 ± 14.01 a/ | 12.65 ± 1.37 ab | 1.88 ± 0.30 | 282.67 ± 18.50 a | 121.75 ± 20.66 ab | 39.25 ± 13.89 a | 50.14 ± 1.20 b/ | |
| 1.5P | T3.1 | 98.67 ± 7.37 b | 2.85 ± 0.44 | 6.20 ± 0.45 a/ | 7.22 ± 0.86 a | 10.09 ± 0.08 b/ | 12.50 ± 0.73 b | 1.72 ± 0.40 | 206.67 ± 13.43 bc/ | 117.00 ± 5.89 b | 17.75 ± 7.23 ab | 55.38 ± 4.04 b/ | |
| 0.3E | T3.1 | 130.00 ± 13.53 a/ | 3.43 ± 0.54 | 5.35 ± 0.26 b | 8.46 ± 0.94 a/ | 10.07 ± 0.06 b/ | 14.40 ± 0.67 a | 1.90 ± 0.35 | 234.33 ± 13.80 b | 137.75 ± 2.87 a | 19.00 ± 2.45 b | 51.04 ± 7.94 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, A.; Marmelo, I.; Chainho, T.; Bolotas, D.; Dias, M.; Cereja, R.; Barata, M.; Pousão-Ferreira, P.; Vieira, E.F.; Delerue-Matos, C.; et al. Improving Farmed Juvenile Gilthead Seabream (Sparus aurata) Stress Response to Marine Heatwaves and Vibriosis Through Seaweed-Based Dietary Modulation. Animals 2025, 15, 1970. https://doi.org/10.3390/ani15131970
Pereira A, Marmelo I, Chainho T, Bolotas D, Dias M, Cereja R, Barata M, Pousão-Ferreira P, Vieira EF, Delerue-Matos C, et al. Improving Farmed Juvenile Gilthead Seabream (Sparus aurata) Stress Response to Marine Heatwaves and Vibriosis Through Seaweed-Based Dietary Modulation. Animals. 2025; 15(13):1970. https://doi.org/10.3390/ani15131970
Chicago/Turabian StylePereira, Alícia, Isa Marmelo, Tomás Chainho, Daniel Bolotas, Marta Dias, Rui Cereja, Marisa Barata, Pedro Pousão-Ferreira, Elsa F. Vieira, Cristina Delerue-Matos, and et al. 2025. "Improving Farmed Juvenile Gilthead Seabream (Sparus aurata) Stress Response to Marine Heatwaves and Vibriosis Through Seaweed-Based Dietary Modulation" Animals 15, no. 13: 1970. https://doi.org/10.3390/ani15131970
APA StylePereira, A., Marmelo, I., Chainho, T., Bolotas, D., Dias, M., Cereja, R., Barata, M., Pousão-Ferreira, P., Vieira, E. F., Delerue-Matos, C., Diniz, M. S., Marques, A., & Maulvault, A. L. (2025). Improving Farmed Juvenile Gilthead Seabream (Sparus aurata) Stress Response to Marine Heatwaves and Vibriosis Through Seaweed-Based Dietary Modulation. Animals, 15(13), 1970. https://doi.org/10.3390/ani15131970