
Mulberry (Morus alba) Twig and Leaf Extracts Ameliorate Obesity-Related Metabolic Disorders via Gut Microbiota Modulation in High-Fat Diet-Fed Mice
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Extract Preparation
2.2. Materials and Instruments
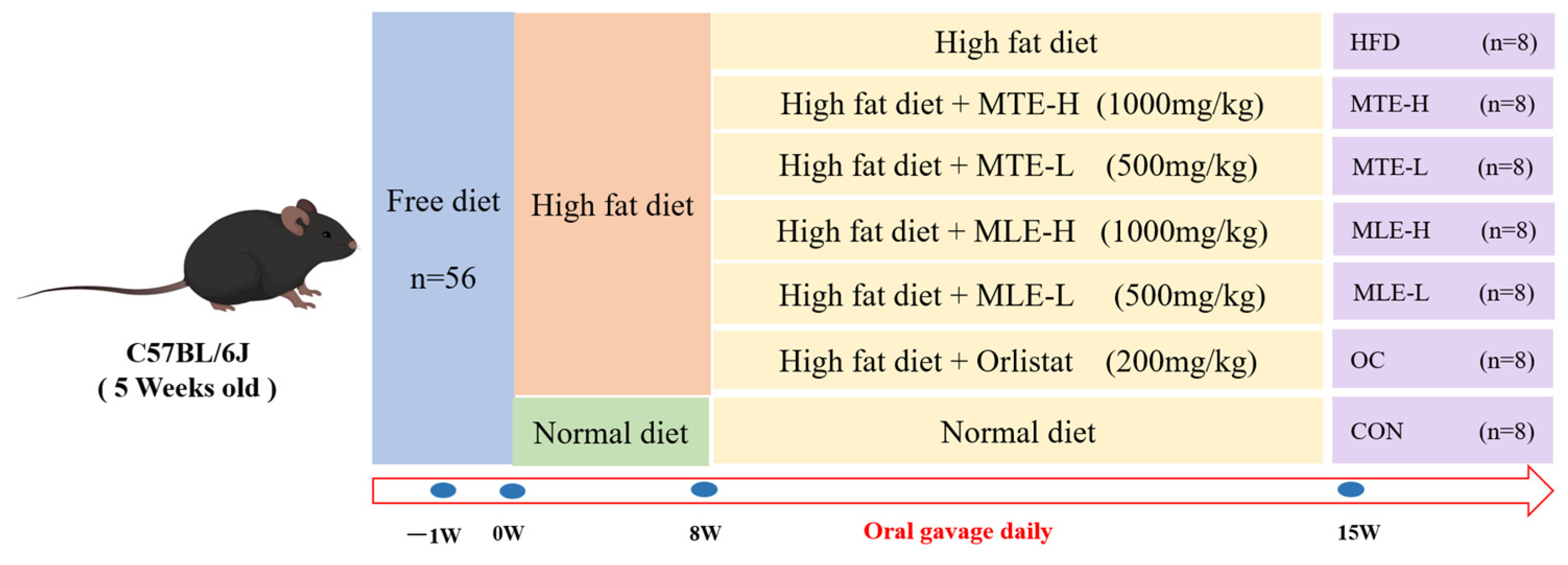
2.3. Animal Handling and Grouping
2.4. Biochemical Analysis
2.5. Histological Analysis of Liver
2.6. Intestinal Flora Analysis
2.7. Short-Chain Fatty Acid Analysis
2.8. Statistical Analysis
3. Results
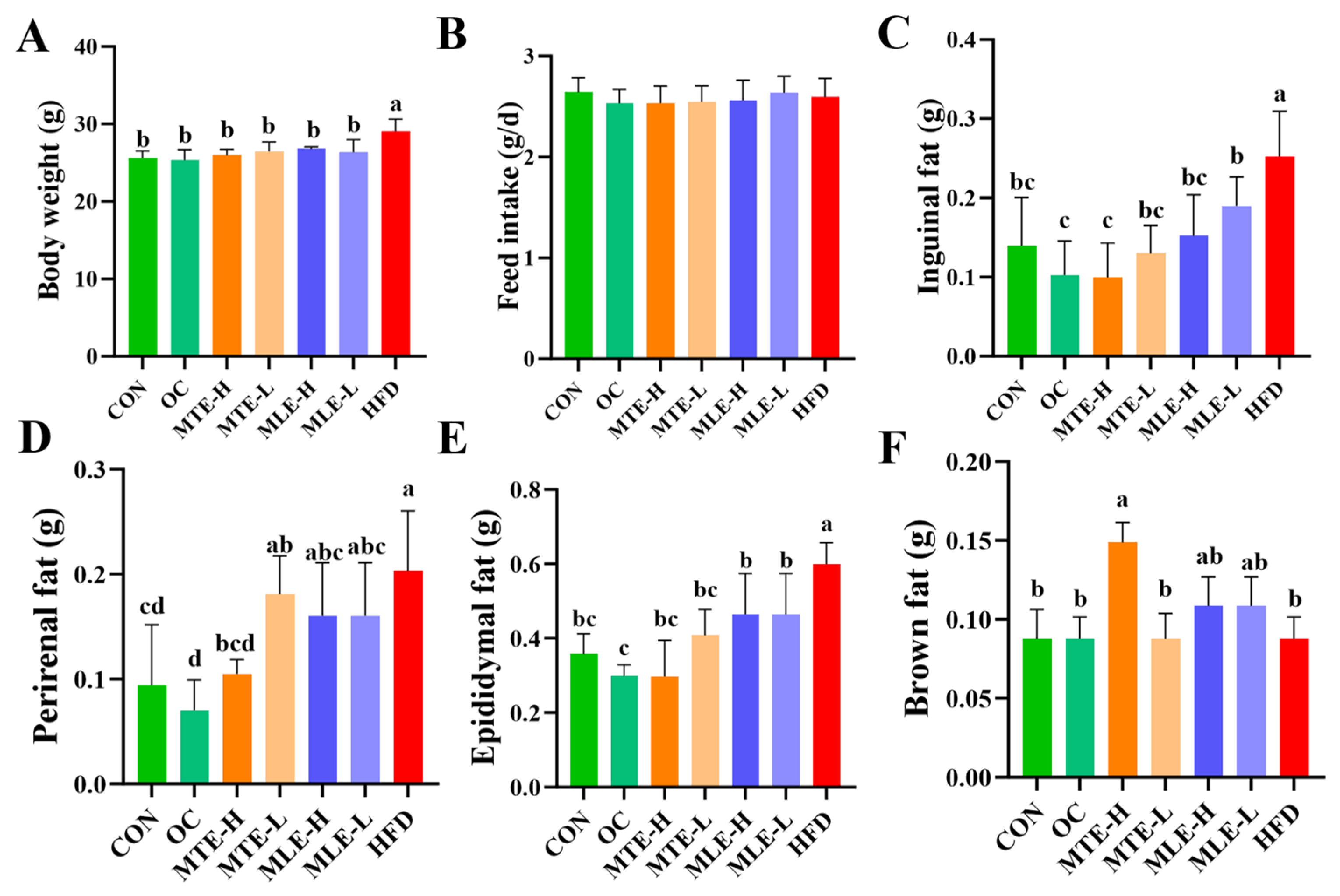
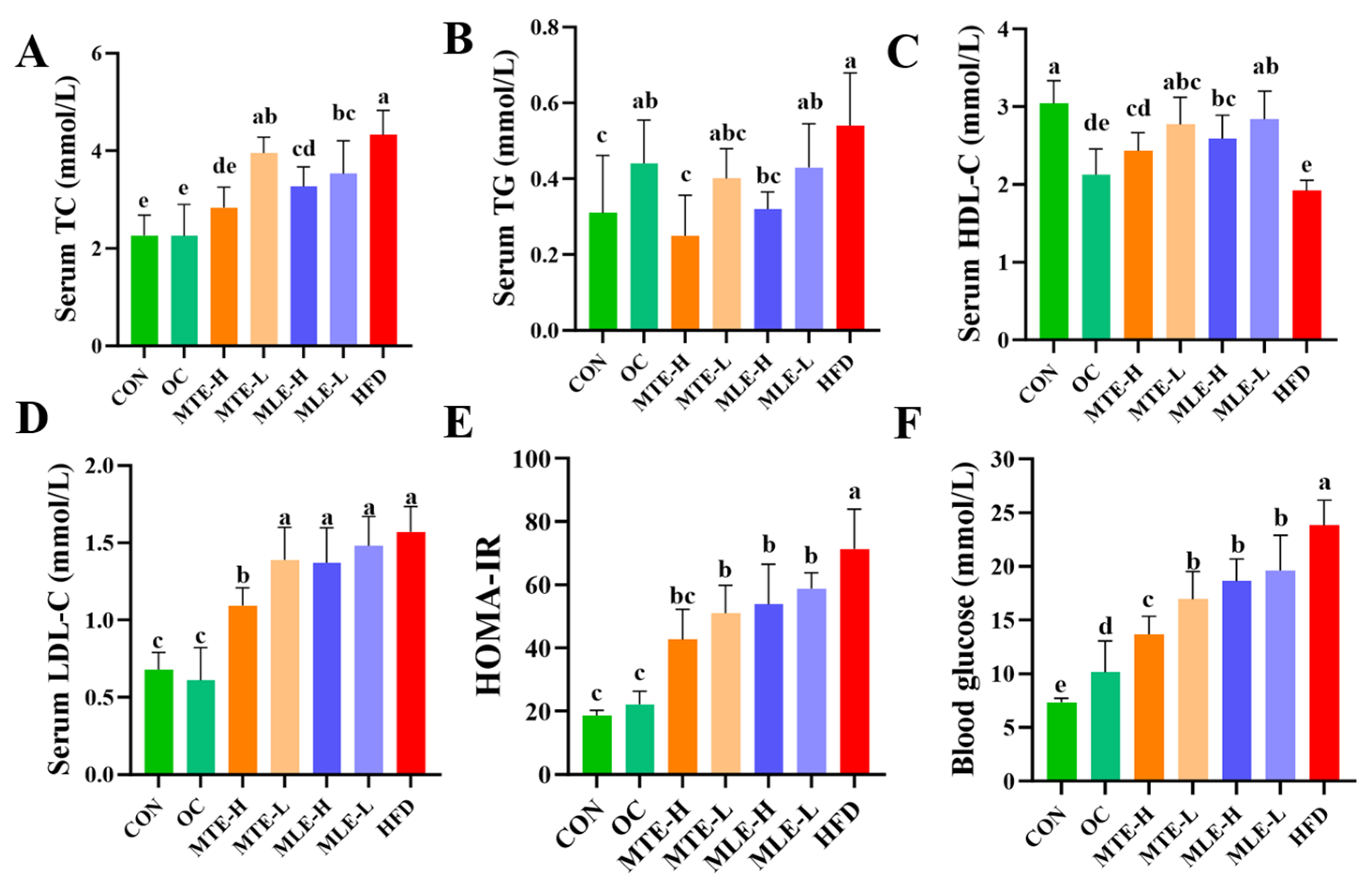
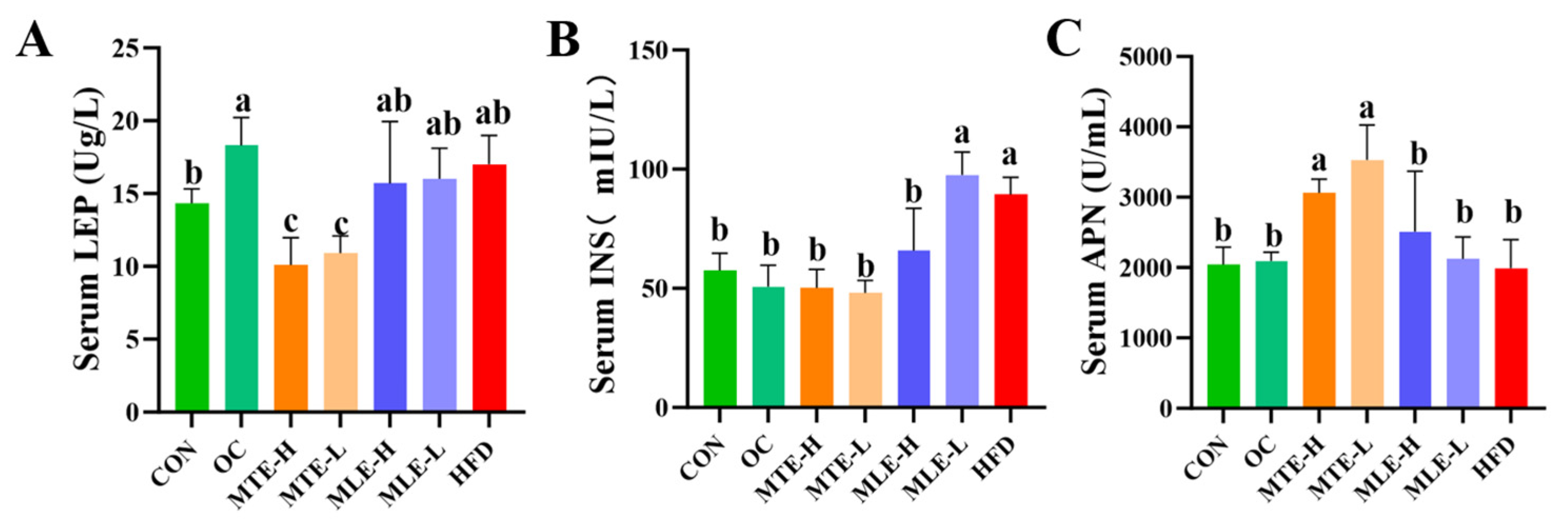
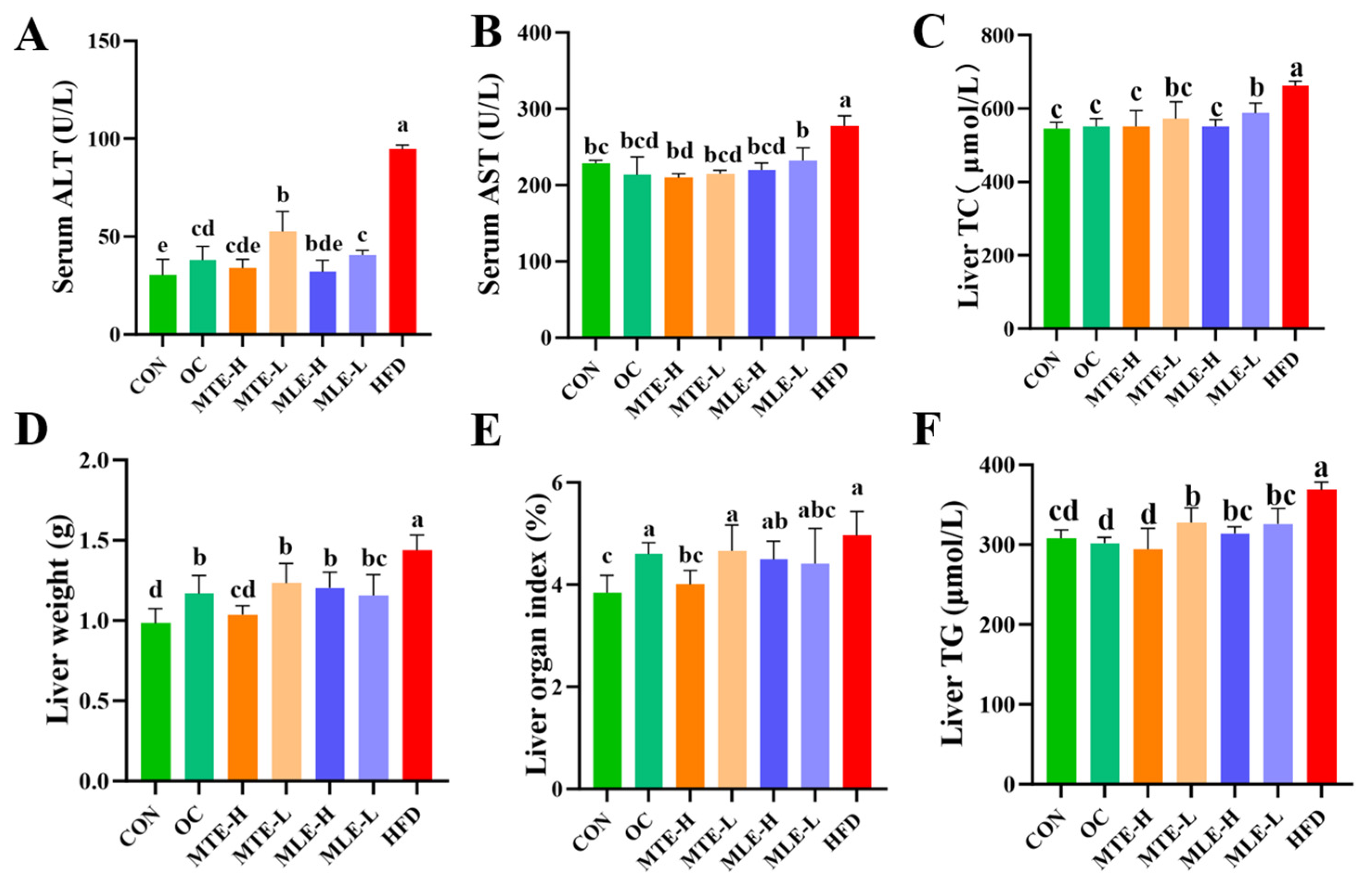
3.1. Effects of MTE and MLE Interventions on Fat Accumulation and Serum Metabolic Parameters in Obese Mice
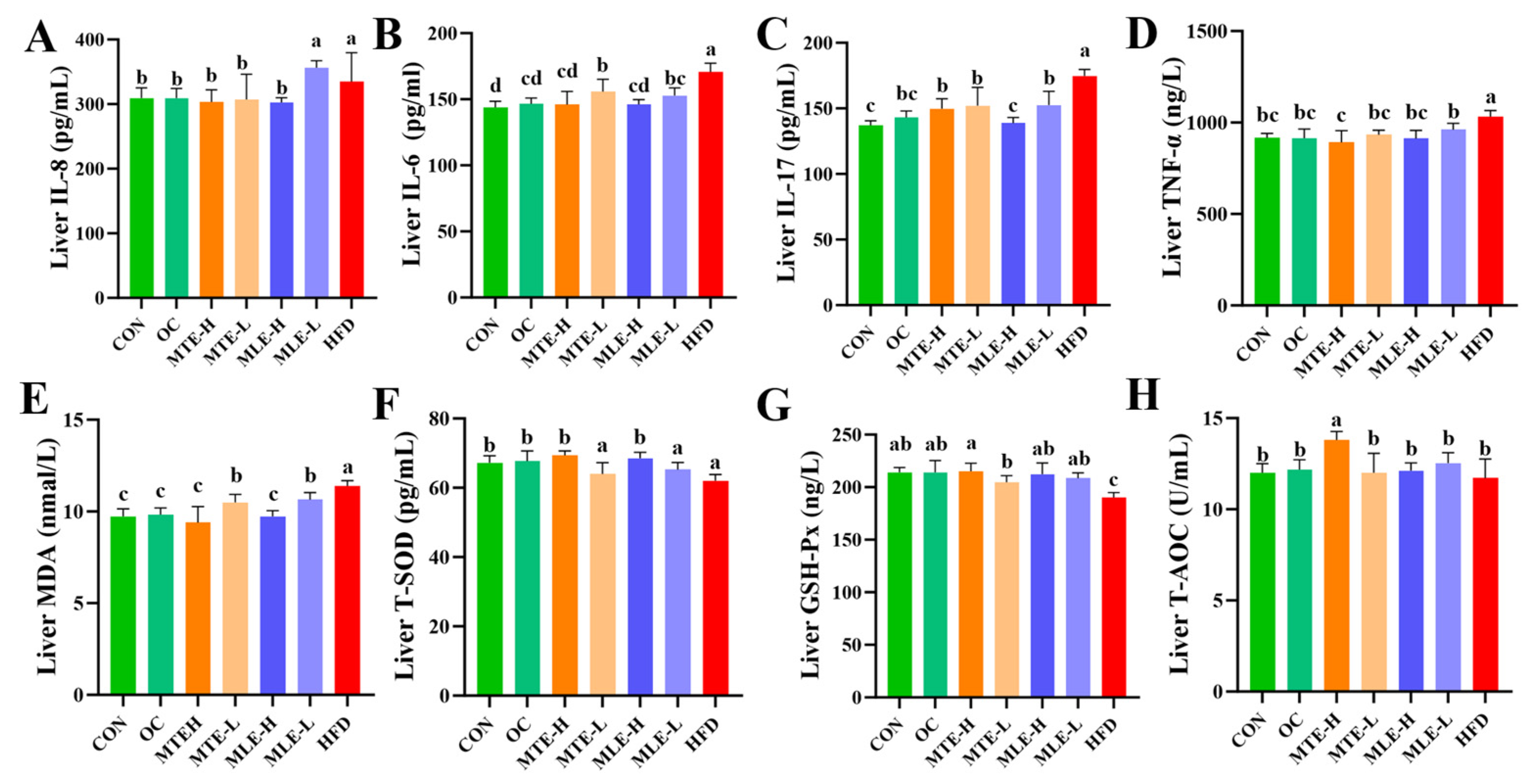
3.2. Effects of MTE and MLE Interventions on Hepatic Metabolic Parameters in Obese Mice
3.3. Pathologic Examination of Liver Tissue Sections
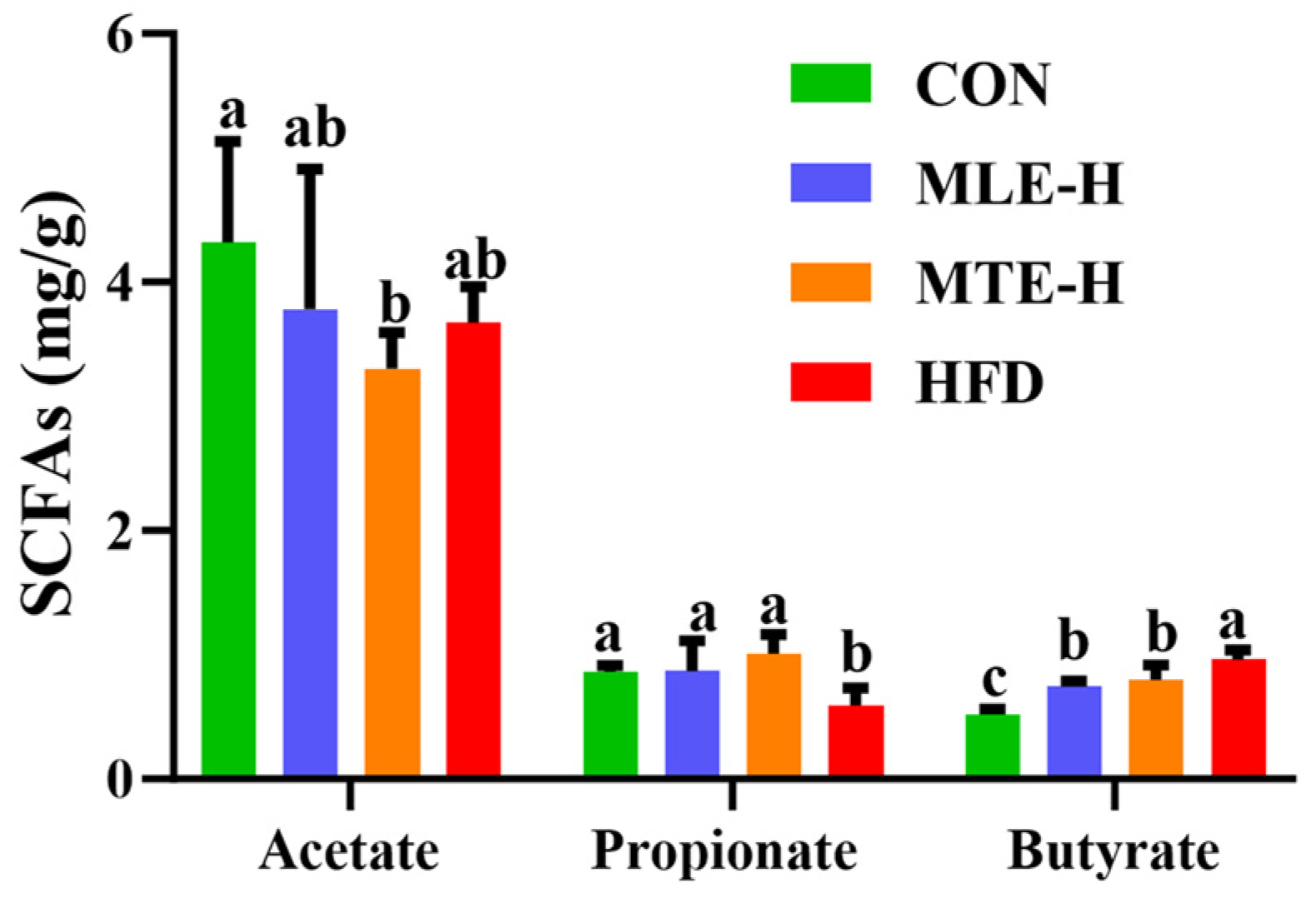
3.4. Effects of MTE and MLE Interventions on Intestinal Short-Chain Fatty Acid Metabolic Profiles in Obese Mice
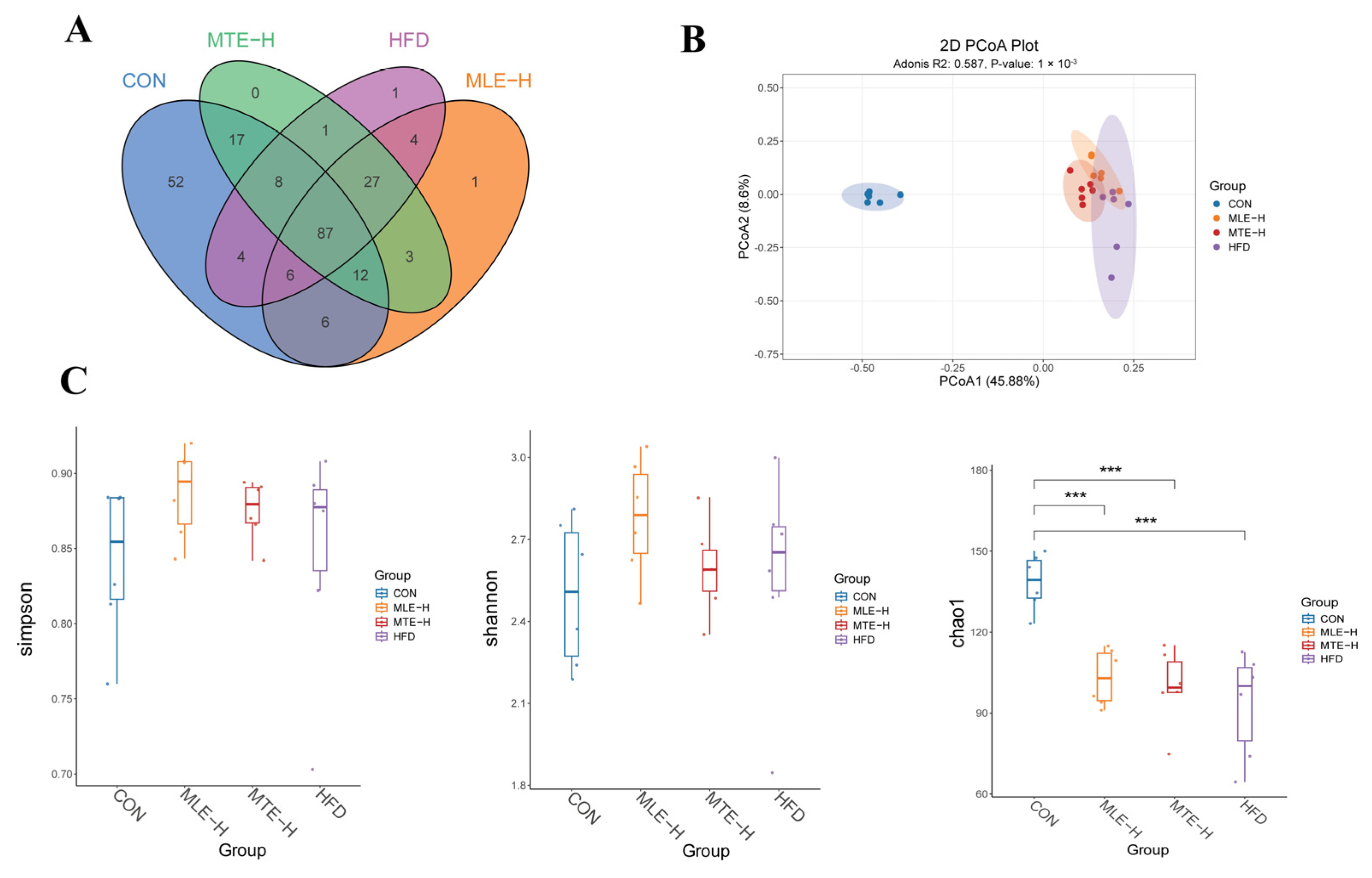
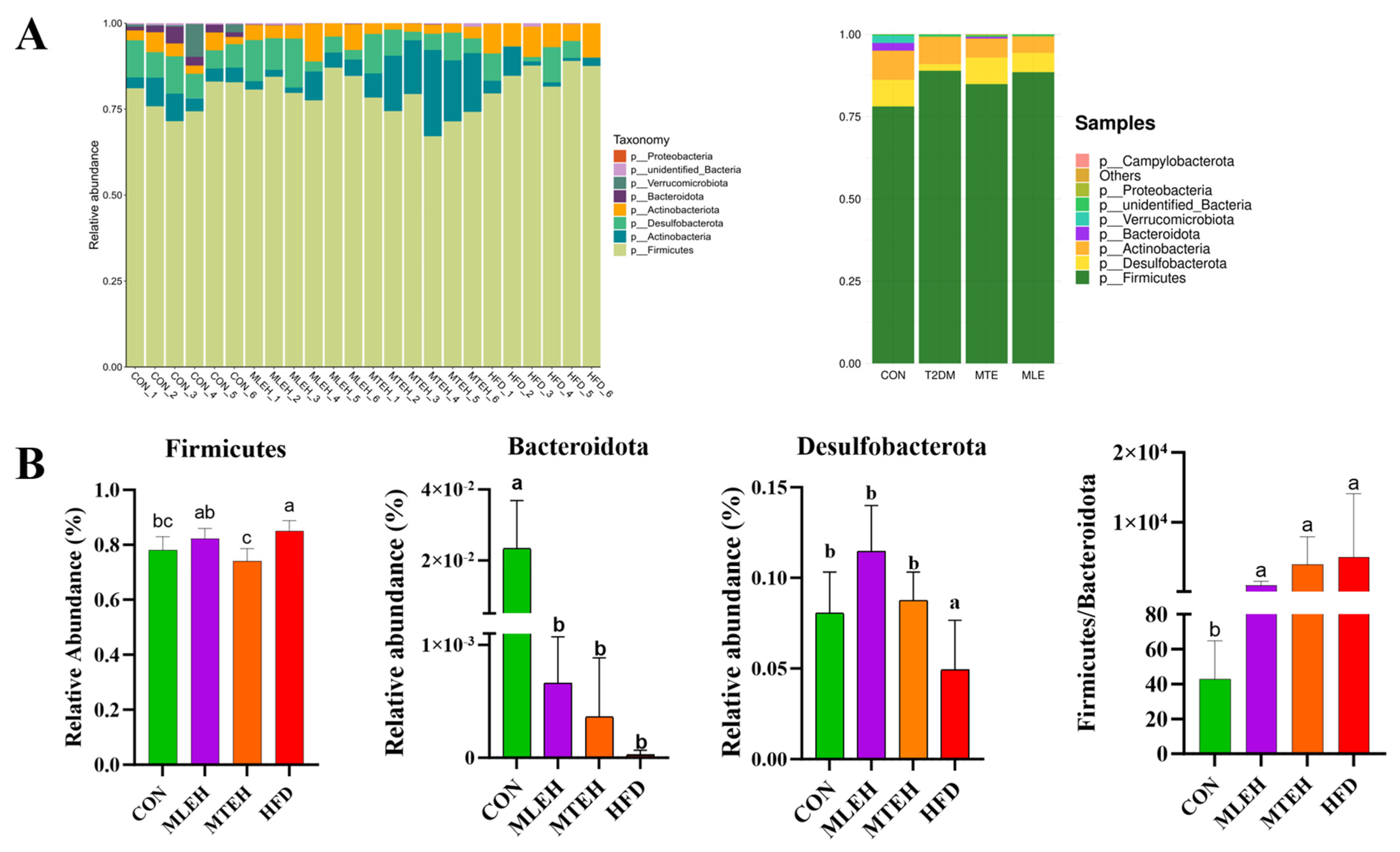
3.5. Effects of MTE and MLE Interventions on the Diversity of Intestinal Flora in Obese Mice
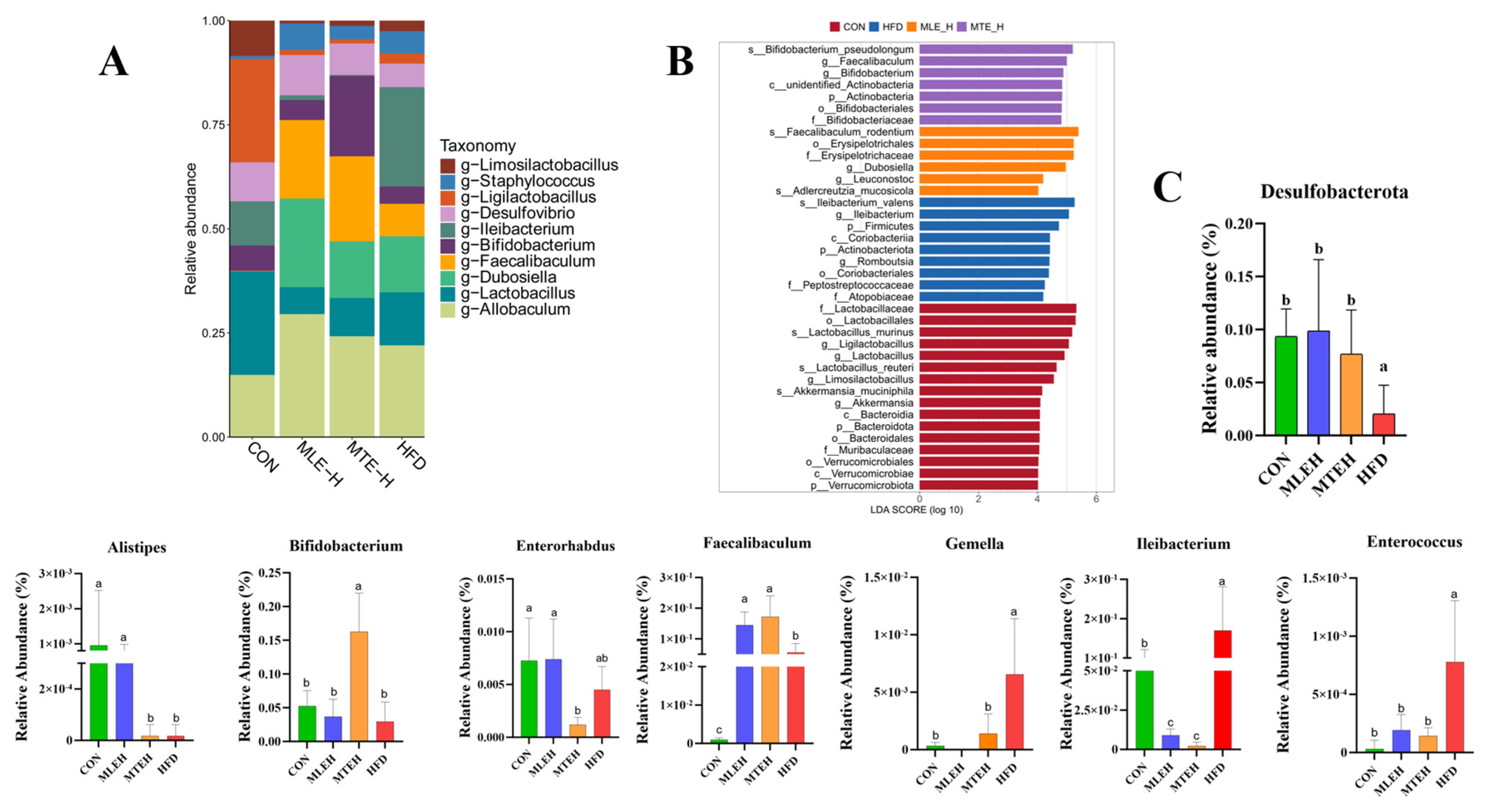
3.6. Effects of MTE and MLE on the Composition of Intestinal Flora in Obese Mice
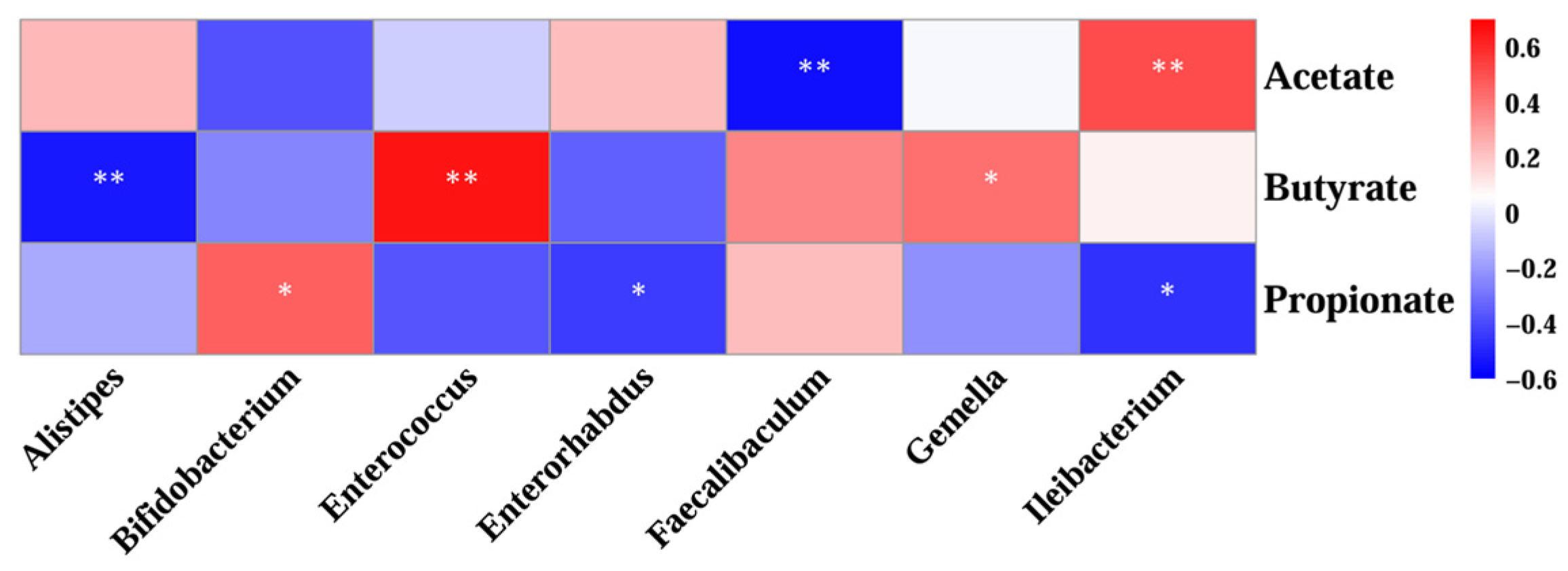
3.7. Correlation Analysis of Gut Microorganisms and Short-Chain Fatty Acid Profiles
4. Discussion
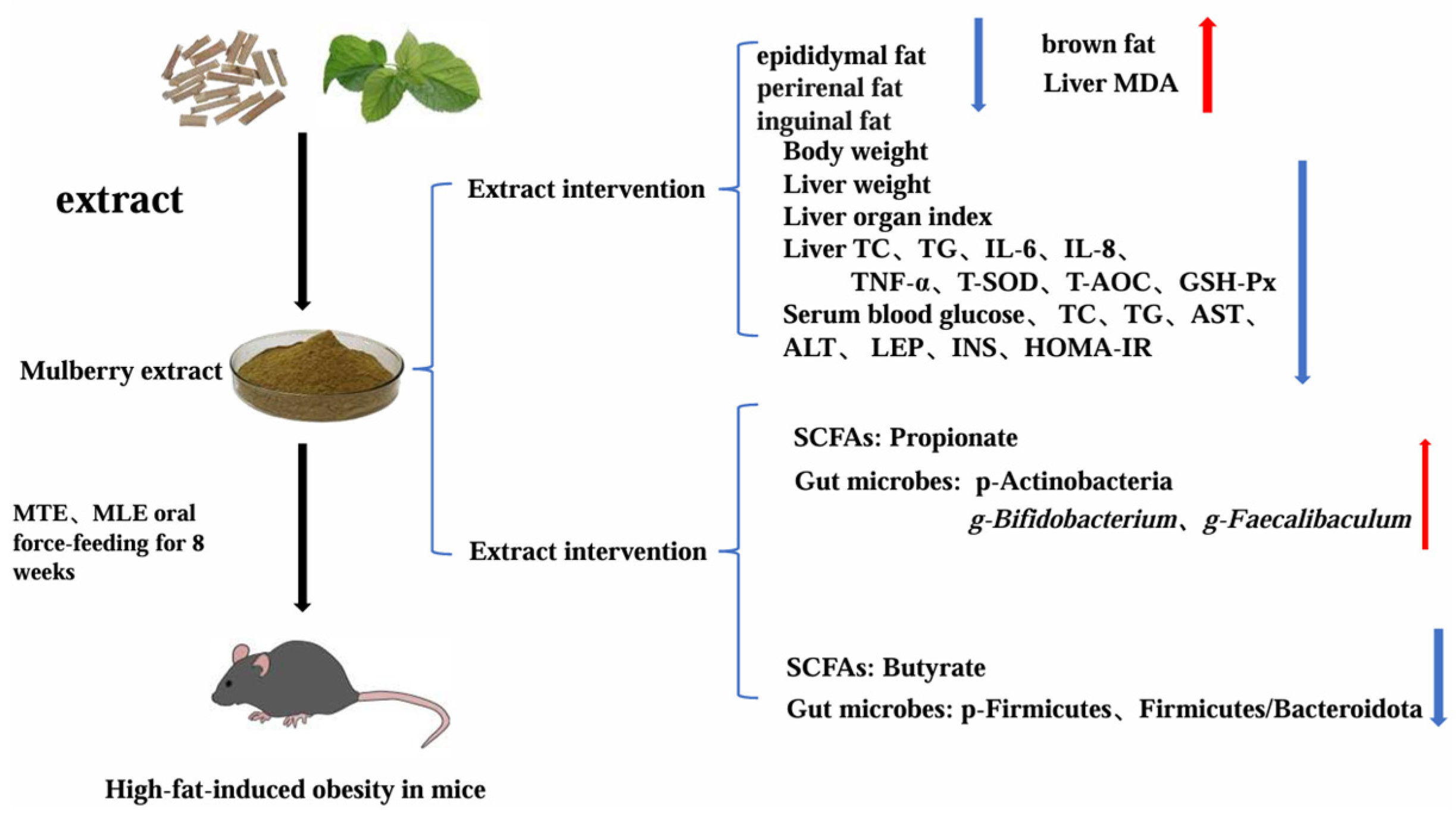
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, J.; Wu, T.; Lam, S.M.; Shui, G.; Yang, S.; Wang, Y.; Tao, C. Heterogeneity of Intramuscular, Intermuscular, and Subcutaneous Fat in Laiwu Pigs: Insights from Targeted Lipidomics and Transcriptomics. Agriculture 2024, 14, 658. [Google Scholar] [CrossRef]
- Fu, Q.; Wang, P.; Zhang, Y.; Wu, T.; Huang, J.; Song, Z. Effects of Dietary Inclusion of Asiaticoside on Growth Performance, Lipid Metabolism, and Gut Microbiota in Yellow-Feathered Chickens. Animals 2023, 13, 2653. [Google Scholar] [CrossRef]
- Velázquez, K.T.; Enos, R.T.; Bader, J.E.; Sougiannis, A.T.; Carson, M.S.; Chatzistamou, I.; Carson, J.A.; Nagarkatti, P.S.; Nagarkatti, M.; Murphy, E.A. Prolonged high-fat-diet feeding promotes non-alcoholic fatty liver disease and alters gut microbiota in mice. World J. Hepatol. 2019, 11, 619–637. [Google Scholar] [CrossRef]
- Tan, R.; Dong, H.; Chen, Z.; Jin, M.; Yin, J.; Li, H.; Shi, D.; Shao, Y.; Wang, H.; Chen, T.; et al. Intestinal Microbiota Mediates High-Fructose and High-Fat Diets to Induce Chronic Intestinal Inflammation. Front. Cell Infect. Microbiol. 2021, 11, 654074. [Google Scholar] [CrossRef]
- Guan, L.; Zhang, L.; Gong, D.; Li, P.; Zhu, S.; Tang, J.; Du, M.; Zhang, M.; Zou, Y. Genipin improves obesity through promoting bile secretion and changing bile acids composition in diet-induced obese rats. J. Pharm. Pharmacol. 2024, 76, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Gui, L.; Wang, S.; Wang, J.; Liao, W.; Chen, Z.; Pan, D.; Xia, H.; Sun, G.; Tian, S. Effects of forsythin extract in Forsythia leaves on intestinal microbiota and short-chain fatty acids in rats fed a high-fat diet. Food Sci. Hum. Wellness 2024, 13, 659–667. [Google Scholar] [CrossRef]
- Li, W.; Fu, X.; Lin, D.; Li, T.; Zhang, N.; Huo, Y.; Zhu, P.; Guo, F.; Huang, F. Conjugated linoleic acid alleviates glycolipid metabolic disorders by modulating intestinal microbiota and short-chain fatty acids in obese rats. Food Funct. 2023, 14, 1685–1698. [Google Scholar] [CrossRef]
- Luo, Y.; Jin, Y.; Wang, H.; Wang, G.; Lin, Y.; Chen, H.; Li, X.; Wang, M. Effects of Clostridium tyrobutyricum on Lipid Metabolism, Intestinal Barrier Function, and Gut Microbiota in Obese Mice Induced by High-Fat Diet. Nutrients 2024, 16, 493. [Google Scholar] [CrossRef]
- Song, B.; Zhong, Y.Z.; Zheng, C.B.; Li, F.N.; Duan, Y.H.; Deng, J.P. Propionate alleviates high-fat diet-induced lipid dysmetabolism by modulating gut microbiota in mice. J. Appl. Microbiol. 2019, 127, 1546–1555. [Google Scholar] [CrossRef]
- Jahromi, M.F.; Liang, J.B.; Ebrahimi, R.; Soleimani, A.F.; Rezaeizadeh, A.; Abdullah, N.; Shokryazdan, P. Protective potential of Lactobacillus species in lead toxicity model in broiler chickens. Animal 2017, 11, 755–761. [Google Scholar] [CrossRef]
- You, M.; Zhou, L.; Wu, F.; Zhang, L.; Zhu, S.X.; Zhang, H.X. Probiotics for the treatment of hyperlipidemia: Focus on gut-liver axis and lipid metabolism. Pharmacol. Res. 2025, 214, 107694. [Google Scholar] [CrossRef]
- Soroko, M.; Górniak, W.; Zielińska, P.; Górniak, A.; Śniegucka, K.; Nawrot, K.; Korczyński, M. Effect of Lentinula edodes on Morphological and Biochemical Blood Parameters of Horses. Animals 2022, 12, 1106. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Ma, H.; Yue, Y.; Wang, L.; Hao, K.; Zhang, Y.; Li, J.; Xiang, Y.; Min, Y. Saikosaponin a ameliorates diet-induced fatty liver via regulating intestinal microbiota and bile acid profile in laying hens. Poult. Sci. 2023, 102, 103155. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Liu, C.; Wu, J.; Cui, Y.; Zhang, M.; Bi, C.; Shan, A.; Dou, X. EGCG improve meat quality, restore lipid metabolism disorder and regulate intestinal flora in high-fat fed broilers. Poult. Sci. 2025, 104, 104875. [Google Scholar] [CrossRef]
- Xue, C. Evaluation of gut microbiota alterations following orlistat administration in obese mice. Front. Endocrinol. 2025, 15, 1337245. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Hong, Q.; Li, X.; Lu, M.; Zhou, J.; Yue, S.; Wang, Z.; Wang, L.; Peng, Q.; Xue, B. Hepatic Injury Induced by Dietary Energy Level via Lipid Accumulation and Changed Metabolites in Growing Semi-Fine Wool Sheep. Front. Vet. Sci. 2021, 8, 745078. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Q.; Abouelfetouh, M.M.; Li, H.; Zhu, H.; Zhu, C.; Kiani, F.A.; Ding, Y. The role of the hypothalamus-gut microbiota in the pathogenesis of periparturient fatty liver disease in dairy cows. Vet. J. 2025, 309, 106290. [Google Scholar] [CrossRef]
- Maqsood, M.; Anam Saeed, R.; Sahar, A.; Khan, M.I. Mulberry plant as a source of functional food with therapeutic and nutritional applications: A review. J. Food Biochem. 2022, 46, e14263. [Google Scholar] [CrossRef]
- Huang, Q.; Xin, X.; Sun, Q.; An, Z.; Gou, X.; Feng, Q. Plant-derived bioactive compounds regulate the NLRP3 inflammasome to treat NAFLD. Front. Pharmacol. 2022, 13, 896899. [Google Scholar] [CrossRef]
- Wang, Y.; Ai, Q.; Gu, M.; Guan, H.; Yang, W.; Zhang, M.; Mao, J.; Lin, Z.; Liu, Q.; Liu, J. Comprehensive overview of different medicinal parts from Morus alba L.: Chemical compositions and pharmacological activities. Front. Pharmacol. 2024, 15, 1364948. [Google Scholar] [CrossRef]
- Đurović, S.; Kojić, I.; Radić, D.; Smyatskaya, Y.A.; Bazarnova, J.G.; Filip, S.; Tosti, T. Chemical Constituents of Stinging Nettle (Urtica dioica L.): A Comprehensive Review on Phenolic and Polyphenolic Compounds and Their Bioactivity. Int. J. Mol. Sci. 2024, 25, 3430. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, S.; Gu, M.; Chen, X.; Chen, X.; Yang, J.; Zhao, F.; Ye, N. Exploration of the Effects of Different Blue LED Light Intensities on Flavonoid and Lipid Metabolism in Tea Plants via Transcriptomics and Metabolomics. Int. J. Mol. Sci. 2020, 21, 4606. [Google Scholar] [CrossRef]
- Nie, Y.; Stürzenbaum, S.R. Proanthocyanidins of Natural Origin: Molecular Mechanisms and Implications for Lipid Disorder and Aging-Associated Diseases. Adv. Nutr. 2019, 10, 464–478. [Google Scholar] [CrossRef]
- Wang, Y.; Zidichouski, J.A. Update on the Benefits and Mechanisms of Action of the Bioactive Vegetal Alkaloid Berberine on Lipid Metabolism and Homeostasis. Cholesterol 2018, 2018, 7173920. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhou, X.; Ling, N.; Yu, Q.; Wang, H.; Du, Q. The Effect of Guisangyou Tea on Abnormal Lipid Metabolism in Mice Induced by High-Fat Diet. Foods 2023, 12, 2171. [Google Scholar] [CrossRef]
- Du, Y.; Li, D.X.; Lu, D.Y.; Zhang, R.; Zhao, Y.L.; Zhong, Q.Q.; Ji, S.; Wang, L.; Tang, D.Q. Lipid metabolism disorders and lipid mediator changes of mice in response to long-term exposure to high-fat and high sucrose diets and ameliorative effects of mulberry leaves. Food Funct. 2022, 13, 4576–4591. [Google Scholar] [CrossRef]
- He, X.; Li, H.; Gao, R.; Zhang, C.; Liang, F.; Sheng, Y.; Zheng, S.; Xu, J.; Xu, W.; Huang, K. Mulberry leaf aqueous extract ameliorates blood glucose and enhances energy expenditure in obese C57BL/6J mice. J. Funct. Foods 2019, 63, 103505. [Google Scholar] [CrossRef]
- Chen, W.; Lu, Y.; Hu, D.; Mo, J.; Ni, J. Black mulberry (Morus nigra L.) polysaccharide ameliorates palmitate-induced lipotoxicity in hepatocytes by activating Nrf2 signaling pathway. Int. J. Biol. Macromol. 2021, 172, 394–407. [Google Scholar] [CrossRef]
- Lim, S.H.; Choi, C.-I. Pharmacological Properties of Morus nigra L. (Black Mulberry) as A Promising Nutraceutical Resource. Nutrients 2019, 11, 437. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.W.; Lian, C.F.; Chen, Y.M.; Ye, J.; Chen, W.; Gao, Y.; Wang, H.L.; Gao, L.L.; Liu, Y.L.; Yang, Y.F. Ramulus Mori (Sangzhi) Alkaloids Ameliorate Obesity-Linked Adipose Tissue Metabolism and Inflammation in Mice. Nutrients 2022, 14, 5050. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xu, S.; Zhang, B.; Sun, X. Ramulus Mori (Sangzhi) Alkaloids Alleviate Diabetic Nephropathy through Improving Gut Microbiota Disorder. Nutrients 2024, 16, 2346. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, M.; Elnaby Kabeel, A.; Chandra Sekhar, S.; Sathyamurthy, R.; Manokar Athikesavan, M. Experimental investigation on seaweed (sargassum wightii) derived using methanolic extracts. Alex. Eng. J. 2023, 68, 453–460. [Google Scholar] [CrossRef]
- Mohammed, E.A.; Abdalla, I.G.; Alfawaz, M.A.; Mohammed, M.A.; Al Maiman, S.A.; Osman, M.A.; Yagoub, A.E.A.; Hassan, A.B. Effects of Extraction Solvents on the Total Phenolic Content, Total Flavonoid Content, and Antioxidant Activity in the Aerial Part of Root Vegetables. Agriculture 2022, 12, 1820. [Google Scholar] [CrossRef]
- Negron, S.G.; Ercan-Sencicek, A.G.; Freed, J.; Walters, M.; Lin, Z. Both proliferation and lipogenesis of brown adipocytes contribute to postnatal brown adipose tissue growth in mice. Sci. Rep. 2020, 10, 20335. [Google Scholar] [CrossRef]
- Dong, Z.; Liu, Z.; Xu, Y.; Tan, B.; Sun, W.; Ai, Q.; Yang, Z.; Zeng, J. Potential for the development of Taraxacum mongolicum aqueous extract as a phytogenic feed additive for poultry. Front. Immunol. 2024, 15, 1354040. [Google Scholar] [CrossRef]
- Wang, R. Solvents effect on phenolics, iridoids, antioxidant activity, antibacterial activity, and pancreatic lipase inhibition activity of noni (Morinda citrifolia L.) fruit extract. Food Chem. 2022, 377, 131989. [Google Scholar] [CrossRef]
- Wang, N.; Xiong, Y.; Wang, X.; Guo, L.; Lin, Y.; Ni, K.; Yang, F. Effects of Lactobacillus plantarum on Fermentation Quality and Anti-Nutritional Factors of Paper Mulberry Silage. Fermentation 2022, 8, 144. [Google Scholar] [CrossRef]
- Available online: https://www.ema.europa.eu/en/ich-q3c-r9-residual-solvents-scientific-guideline (accessed on 12 June 2025).
- Jacob, T.; Sindhu, S.; Hasan, A.; Malik, M.Z.; Arefanian, H.; Al-Rashed, F.; Nizam, R.; Kochumon, S.; Thomas, R.; Bahman, F.; et al. Soybean oil-based HFD induces gut dysbiosis that leads to steatosis, hepatic inflammation and insulin resistance in mice. Front. Microbiol. 2024, 15, 1407258. [Google Scholar] [CrossRef]
- Filippatos, T.D.; Derdemezis, C.S.; Gazi, I.F.; Nakou, E.S.; Mikhailidis, D.P.; Elisaf, M.S. Orlistat-associated adverse effects and drug interactions: A critical review. Drug Saf. 2008, 31, 53–65. [Google Scholar] [CrossRef]
- Zhong, Y.; Song, B.; Zheng, C.; Zhang, S.; Yan, Z.; Tang, Z.; Kong, X.; Duan, Y.; Li, F. Flavonoids from Mulberry Leaves Alleviate Lipid Dysmetabolism in High Fat Diet-Fed Mice: Involvement of Gut Microbiota. Microorganisms 2020, 8, 860. [Google Scholar] [CrossRef] [PubMed]
- Suchacki, K.J.; Stimson, R.H. Nutritional Regulation of Human Brown Adipose Tissue. Nutrients 2021, 13, 1748. [Google Scholar] [CrossRef]
- Noriega, L.; Yang, C.Y.; Wang, C.H. Brown Fat and Nutrition: Implications for Nutritional Interventions. Nutrients 2023, 15, 4072. [Google Scholar] [CrossRef]
- Pei, Y.; Otieno, D.; Gu, I.; Lee, S.O.; Parks, J.S.; Schimmel, K.; Kang, H.W. Effect of quercetin on nonshivering thermogenesis of brown adipose tissue in high-fat diet-induced obese mice. J. Nutr. Biochem. 2021, 88, 108532. [Google Scholar] [CrossRef]
- Yuan, X.; Wei, G.; You, Y.; Huang, Y.; Lee, H.J.; Dong, M.; Lin, J.; Hu, T.; Zhang, H.; Zhang, C.; et al. Rutin ameliorates obesity through brown fat activation. FASEB J. 2017, 31, 333–345. [Google Scholar] [CrossRef]
- Nathalie, G.; Bonamichi, B.; Kim, J.; Jeong, J.; Kang, H.; Hartland, E.R.; Eveline, E.; Lee, J. NK cell-activating receptor NKp46 does not participate in the development of obesity-induced inflammation and insulin resistance. Mol. Cells 2024, 47, 100007. [Google Scholar] [CrossRef]
- Villanueva, E.C.; Myers, M.G., Jr. Leptin receptor signaling and the regulation of mammalian physiology. Int. J. Obes. 2008, 32 (Suppl. 7), S8–S12. [Google Scholar] [CrossRef]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Soták, M.; Clark, M.; Suur, B.E.; Börgeson, E. Inflammation and resolution in obesity. Nat. Rev. Endocrinol. 2025, 21, 45–61. [Google Scholar] [CrossRef]
- O’Riordan, K.J.; Moloney, G.M.; Keane, L.; Clarke, G.; Cryan, J.F. The gut microbiota-immune-brain axis: Therapeutic implications. Cell Rep. Med. 2025, 6, 101982. [Google Scholar] [CrossRef]
- Saoudi, M.; El Feki, A. Protective Role of Ficus carica Stem Extract against Hepatic Oxidative Damage Induced by Methanol in Male Wistar Rats. Evid. Based Complement. Altern. Med. 2012, 2012, 150458. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, Y.; Su, L.; Ren, H.; Wang, C.; Chen, J.; Fu, X. Donepezil Attenuates Obesity-Associated Oxidative Stress and Central Inflammation and Improves Memory Deficit in Mice Fed a High-Fat Diet. Dement. Geriatr. Cogn. Disord. 2019, 48, 154–163. [Google Scholar] [CrossRef]
- Wang, T.; He, C. Pro-inflammatory cytokines: The link between obesity and osteoarthritis. Cytokine Growth Factor Rev. 2018, 44, 38–50. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, C.C. Inflammatory response of macrophages in infection. Hepatobiliary Pancreat. Dis. Int. 2014, 13, 138–152. [Google Scholar] [CrossRef]
- Fernandez, M.L. Soluble fiber and nondigestible carbohydrate effects on plasma lipids and cardiovascular risk. Curr. Opin. Lipidol. 2001, 12, 35–40. [Google Scholar] [CrossRef]
- Salonen, A.; Lahti, L.; Salojärvi, J.; Holtrop, G.; Korpela, K.; Duncan, S.H.; Date, P.; Farquharson, F.; Johnstone, A.M.; Lobley, G.E.; et al. Impact of diet and individual variation on intestinal microbiota composition and fermentation products in obese men. ISME J. 2014, 8, 2218–2230. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef]
- Wen, J.J.; Li, M.Z.; Gao, H.; Hu, J.L.; Nie, Q.X.; Chen, H.H.; Zhang, Y.L.; Xie, M.Y.; Nie, S.P. Polysaccharides from fermented Momordica charantia L. with Lactobacillus plantarum NCU116 ameliorate metabolic disorders and gut microbiota change in obese rats. Food Funct. 2021, 12, 2617–2630. [Google Scholar] [CrossRef]
- Wu, L.; Park, S.H.; Kim, H. Direct and Indirect Evidence of Effects of Bacteroides spp. on Obesity and Inflammation. Int. J. Mol. Sci. 2023, 25, 438. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Chen, L.; Zhang, L.; Xu, F.; Zhang, C.; Ren, G.; Chang, K.; He, G.; Du, Z.; Le, Y.; et al. Xie Zhuo Tiao Zhi formula modulates intestinal microbiota and liver purine metabolism to suppress hepatic steatosis and pyroptosis in NAFLD therapy. Phytomedicine 2023, 121, 155111. [Google Scholar] [CrossRef] [PubMed]
- Taglialegna, A. Fat, Desulfovibrio and cancer. Nat. Rev. Microbiol. 2024, 22, 388. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, H.; Ye, X.; Chen, S. Proanthocyanidins from Chinese bayberry leaves reduce obesity and associated metabolic disorders in high-fat diet-induced obese mice through a combination of AMPK activation and an alteration in gut microbiota. Food Funct. 2022, 13, 2295–2305. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Wang, Y.; Lin, Y.; Lang, Y.; Li, E.; Zhang, X.; Zhang, Q.; Feng, Y.; Meng, X.; Li, B. Blueberry polyphenols extract as a potential prebiotic with anti-obesity effects on C57BL/6 J mice by modulating the gut microbiota. J. Nutr. Biochem. 2019, 64, 88–100. [Google Scholar] [CrossRef]
- Yue, S.; Zhao, D.; Peng, C.; Tan, C.; Wang, Q.; Gong, J. Effects of theabrownin on serum metabolites and gut microbiome in rats with a high-sugar diet. Food Funct. 2019, 10, 7063–7080. [Google Scholar] [CrossRef]
- Hong, Y.; Sheng, L.; Zhong, J.; Tao, X.; Zhu, W.; Ma, J.; Yan, J.; Zhao, A.; Zheng, X.; Wu, G.; et al. Desulfovibrio vulgaris, a potent acetic acid-producing bacterium, attenuates nonalcoholic fatty liver disease in mice. Gut Microbes 2021, 13, 1930874. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, Z. Probiotics improved hyperlipidemia in mice induced by a high cholesterol diet via downregulating FXR. Food Funct. 2020, 11, 9903–9911. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, W.; Han, J.; Shi, X.; Qin, X.; Jiao, F.; Zhang, M.; Bao, L.; Su, C. Mulberry (Morus alba) Twig and Leaf Extracts Ameliorate Obesity-Related Metabolic Disorders via Gut Microbiota Modulation in High-Fat Diet-Fed Mice. Animals 2025, 15, 1768. https://doi.org/10.3390/ani15121768
Qian W, Han J, Shi X, Qin X, Jiao F, Zhang M, Bao L, Su C. Mulberry (Morus alba) Twig and Leaf Extracts Ameliorate Obesity-Related Metabolic Disorders via Gut Microbiota Modulation in High-Fat Diet-Fed Mice. Animals. 2025; 15(12):1768. https://doi.org/10.3390/ani15121768
Chicago/Turabian StyleQian, Wei, Jinyan Han, Xiang Shi, Xiaoqing Qin, Feng Jiao, Minjuan Zhang, Lijun Bao, and Chao Su. 2025. "Mulberry (Morus alba) Twig and Leaf Extracts Ameliorate Obesity-Related Metabolic Disorders via Gut Microbiota Modulation in High-Fat Diet-Fed Mice" Animals 15, no. 12: 1768. https://doi.org/10.3390/ani15121768
APA StyleQian, W., Han, J., Shi, X., Qin, X., Jiao, F., Zhang, M., Bao, L., & Su, C. (2025). Mulberry (Morus alba) Twig and Leaf Extracts Ameliorate Obesity-Related Metabolic Disorders via Gut Microbiota Modulation in High-Fat Diet-Fed Mice. Animals, 15(12), 1768. https://doi.org/10.3390/ani15121768