
Cutaneous Allodynia of the Withers in Cattle: An Experimental In Vivo Neuroanatomical Preliminary Investigation of the Dichotomizing Sensory Neurons Projecting into the Reticulum and Skin of the Withers—A Case Study on Two Calves
 ,
,  ,
,  and
and {kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Study Design—Neuroanatomical Experimental Study with a Descriptive Observational Design
- -
- Age between 30 and 120 days old.
- -
- Body weight ranging from 40 to 90 kg.
- -
- Both sexes (male and female).
- -
- Weaned and unweaned animals.
- -
- Clinically healthy at the time of enrollment.
- -
- Presence of any comorbidities with clinical signs at the time of evaluation.
- -
- History of disease within 30 days prior to the study.
- -
- Participation in other clinical or experimental studies concurrently.
- -
- Administration of any medication within 15 days prior to enrollment.
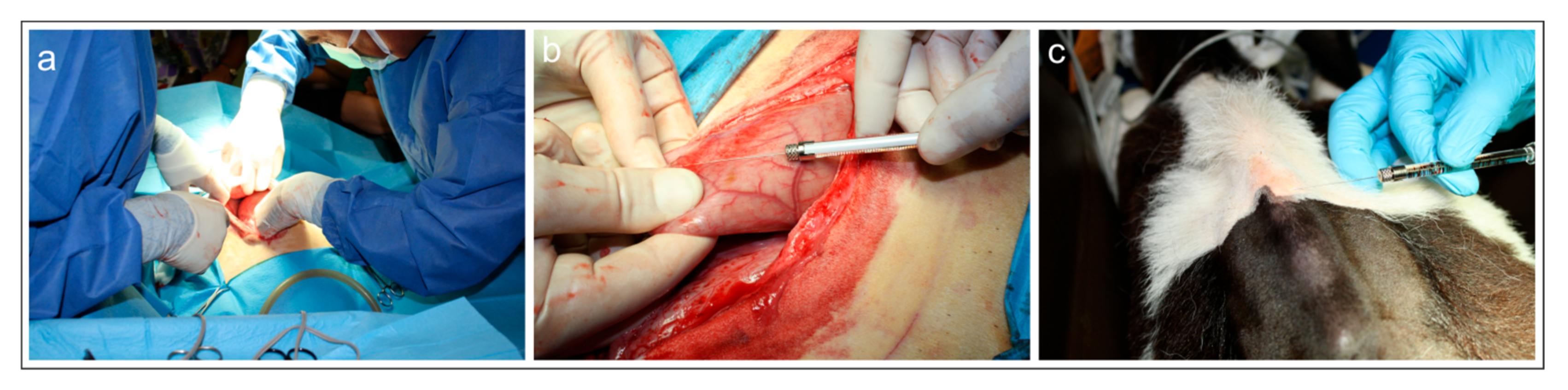
2.2. Injection of the Fluorescent Tracers
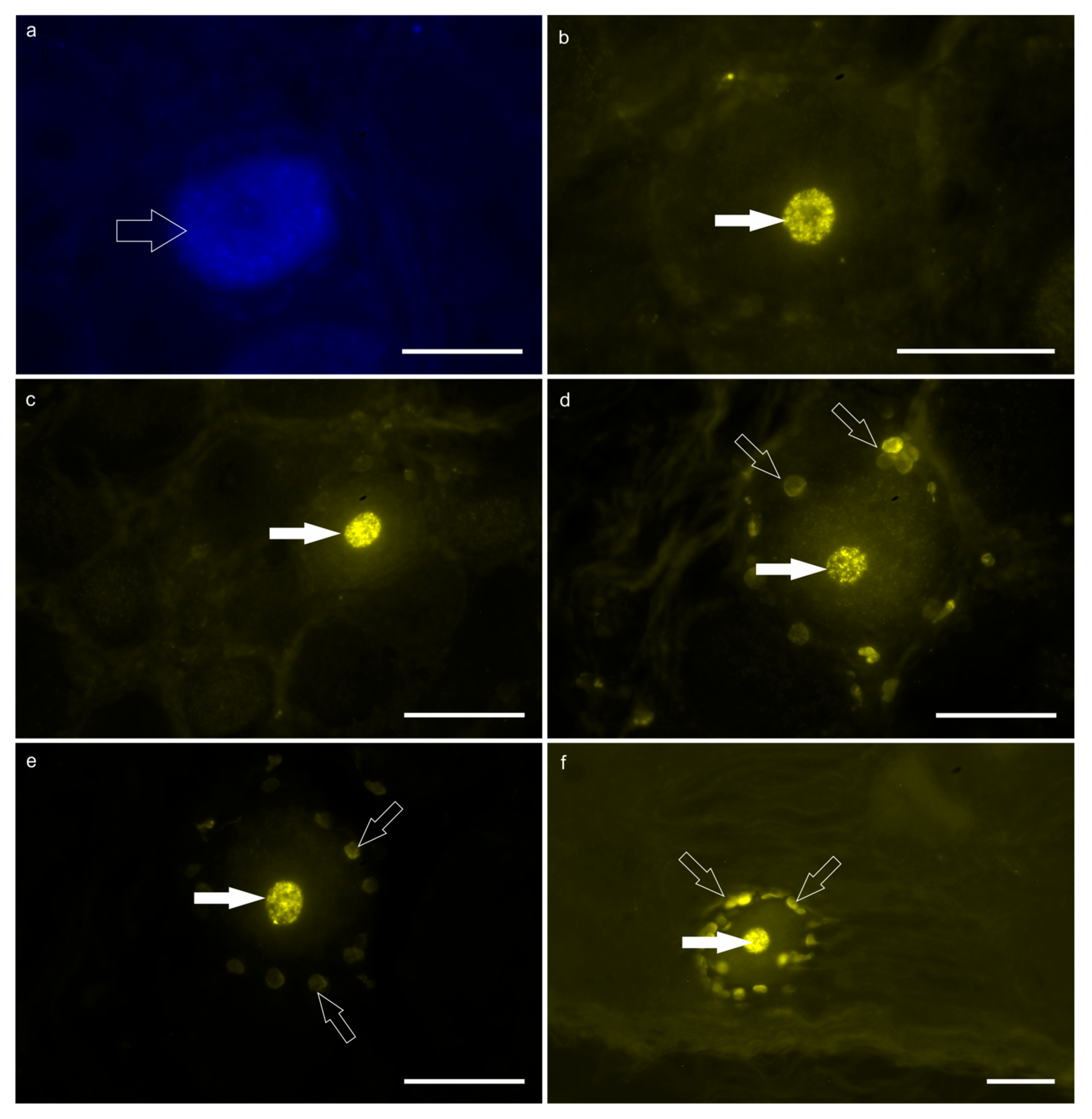
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Data from Cardiorespiratory Monitoring During Calf Anesthesia
- -
- HR and rhythm from 70–100 bpm (considering that younger calves tend to have higher HR);
- -
- ECG normal sinus rhythm, no bradycardia, tachycardia or arrhythmias (avoid deep planes of anaesthesia);
- -
- Pulse intensity (systolic 90–130 mmHg, diastolic 60–90 mmHg) MAP from 70–90, but always >60 mmHg—monitored with non-invasive oscillometric system; important to maintain perfusion;
- -
- Capillary refill time < 2 s, clinical estimate of perfusion;
- -
- Mucous membranes colo—pink;
- -
- RR 15–25 breaths/min set by controlled ventilation (respiratory depression is a common complication in anesthetized calves and is often a result of anesthetic-induced respiratory depression and the abnormal position, e.g., dorsal recumbency, required for these surgical procedures);
- -
- SpO2 > 95%—lower values suggest hypoxemia;
- -
- ETCO2 35–49 mmHg—monitor for hypoventilation;
- -
- Body temperature 37.00–36.2 °C—risk of hypothermia.
References
- Kalchschmidt, H.G. Die “Fremdkoerperzone” eine Headsche Zone beim Rind. Schweiz. Arch. Tierheilk 1948, 90, 433–443. [Google Scholar]
- Kalchschmidt, H.G. Eine Headsche Zone als diagnostisches Hilfsmittel bei der Fremdkoerpererkrankung des Rindes. Wien. Tierzaerztl Monatsschr 1953, 41, 531–550. [Google Scholar]
- Head, H. On disturbances of sensation with especial reference to the pain of visceral disease. Brain 1893, 16, 1–133. [Google Scholar] [CrossRef]
- Cervero. Neurophysiology of gastrointestinal pain. Bailliers Clin. Gastroenterol. 1998, 2, 183–199. [Google Scholar]
- Robinson, D.R.; Gebhart, G.F. Inside information: The unique features of visceral sensation. Mol. Interv. 2008, 8, 242–253. [Google Scholar] [CrossRef]
- Christianson, J.A.; Liang, R.; Ustinova, E.E.; Davis, B.M.; Fraser, M.O.; Pezzone, M.A. Convergence of bladder and colon sensory innervation occurs at the primary afferent level. Pain 2007, 128, 235–243. [Google Scholar] [CrossRef]
- Vermeulen, W.; De Man, J.G.; Pelckmans, P.A.; De Winter, B.Y. Neuroanatomy of lower gastrointestinal pain disorders. World J. Gastroenterol. 2014, 20, 1005–1020. [Google Scholar] [CrossRef]
- De Winter, B.Y.; Deiteren, A.; De Man, J.G. Novel nervous system mechanisms in visceral pain. Neurogastroenterol. Motil. 2016, 28, 309–315. [Google Scholar] [CrossRef]
- Calvino, B.; Grilo, R.M. Central pain control. Jt. Bone Spine 2006, 73, 10–16. [Google Scholar] [CrossRef]
- Luz, L.L.; Fernandes, E.C.; Sivado, M.; Kokai, E.; Szucs, P.; Safronov, B.V. Monosynaptic convergence of somatic and visceral C-fiber afferents on projection and local circuit neurons in lamina I: A substrate for referred pain. Pain 2015, 156, 2042–2051. [Google Scholar] [CrossRef]
- Mayer, E.A.; Gupta, A.; Kilpatrick, L.A.; Hong, J.-Y. Imaging brain mechanisms in chronic visceral pain. Pain 2015, 156, S50–S63. [Google Scholar] [CrossRef] [PubMed]
- Shibata, Y.; Ugawa, S.; Imura, M.; Kubota, Y.; Ueda, T.; Kojima, Y.; Ishida, Y.; Sasaki, S.; Hayashi, Y.; Kohri, K.; et al. TRPM8-expressing dorsal root ganglion neurons project dichotomizing axons to both skin and bladder in rats. Neuroreport 2011, 22, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Pierau, F.K.; Fellmer, G.; Taylor, D.C. Somato-visceral convergence in cat dorsal root ganglion neurones demonstrated by double-labelling with fluorescent tracers. Brain Res. 1984, 321, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, X.; Liu, J.; Tang, W.; Zhao, T.; Zhang, J. Distribution of convergent afferents innervating bladder and prostate at dorsal root Ganglia in rats. Urology 2010, 76, 764.e1-6. [Google Scholar] [CrossRef]
- Li, C.; Zhu, Y.; Shenoy, M.; Pai, R.; Liu, L.; Pasricha, P.J. Anatomical and functional characterization of a duodeno-pancreatic neural reflex that can induce acute pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G490–G500. [Google Scholar] [CrossRef]
- Sameda, H.; Takahashi, Y.; Takahashi, K.; Chiba, T.; Ohtori, S.; Moriya, H. Primary sensory neurons with dichotomizing axons projecting to the facet joint and the sciatic nerve in rats. Spine 2001, 26, 1105–1109. [Google Scholar] [CrossRef]
- Wakai, K.; Ohtori, S.; Yamashita, M.; Yamauchi, K.; Inoue, G.; Suzuki, M.; Orita, S.; Eguchi, Y.; Ochiai, N.; Kishida, S.; et al. Primary sensory neurons with dichotomizing axons projecting to the facet joint and the low back muscle in rats. J. Orthop. Sci. 2010, 15, 402–406. [Google Scholar] [CrossRef]
- Umimura, T.; Miyagi, M.; Ishikawa, T.; Kamoda, H.; Wakai, K.; Sakuma, T.; Sakai, R.; Kuniyoshi, K.; Ochiai, N.; Kishida, S.; et al. Investigation of dichotomizing sensory nerve fibers projecting to the lumbar multifidus muscles and intervertebral disc or facet joint or sacroiliac joint in rats. Spine 2012, 37, 557–562. [Google Scholar] [CrossRef]
- Costa, G.L.; Leonardi, F.; Interlandi, C.; Licata, P.; Lizarraga, I.; Macrì, F.; Macrì, D.; Ferrantelli, V.; Spadola, F. Tramadol administered intravenously either as a bolus or a slow injection in pain management of romifidine- sedated calves undergoing umbilical hernia repair. Animals 2023, 13, 1145. [Google Scholar] [CrossRef]
- Chiocchetti, R.; Clavenzani, P.; Barazzoni, A.M.; Grandis, A.; Bombardi, C.; Costerbosa, G.L.; Petrosino, G.; Avoni, G.B.; Bortolami, R. Viscerotopic representation of the subdiaphragmatic tracts of the digestive apparatus within the vagus complex in the sheep. Brain Res. 2003, 961, 32–44. [Google Scholar] [CrossRef]
- Chiocchetti, R.; Bombardi, C.; Grandis, A.; Mazzuoli, G.; Gentile, A.; Pisoni, L.; Joechler, M.; Lucchi, M.L. Cytoarchitecture, morphology, and lumbosacral spinal cord projections of the red nucleus in cattle. Am. J. Vet. Res. 2006, 67, 1662–1669. [Google Scholar] [CrossRef] [PubMed]
- Russo, D.; Bombardi, C.; Grandis, A.; Furness, J.; Spadari, A.; Bernardini, C.; Chiocchetti, R. Sympathetic innervation of the ileocecal junction in horses. J. Comp. Neurol. 2010, 518, 4046–4066. [Google Scholar] [CrossRef] [PubMed]
- Russo, D.; Clavenzani, P.; Sorteni, C.; Minelli, L.B.; Botti, M.; Gazza, F.; Panu, R.; Ragionieri, L.; Chiocchetti, R. Neurochemical features of boar lumbosacral dorsal root ganglion neurons and characterization of sensory neurons innervating the urinary bladder trigone. J. Comp. Neurol. 2013, 521, 342–366. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, Y.; Atoji, Y.; Saito, S.; Ueno, H.; Inoshima, Y.; Ishiguro, N. Differences in extrinsic innervation patterns of the small intestine in the cattle and sheep. Auton. Neurosci. 2012, 167, 39–44. [Google Scholar] [CrossRef]
- Cottrell, D.F.; Greenhorn, J.G. The vagal and spinal innervation of the gastro-duodenal junction of sheep. Q. J. Exp. Physiol. 1987, 72, 513–524. [Google Scholar] [CrossRef]
- Harding, R.; Leek, B.F. Central projections of gastric afferent vagal inputs. J. Physiol. 1973, 228, 73–90. [Google Scholar] [CrossRef]
- Torigoe, Y.; Cernucan, R.D.; Nishimoto, J.A.; Blanks, R.H. Sympathetic preganglionic efferent and afferent neurons mediated by the greater splanchnic nerve in rabbit. Exp. Neurol. 1985, 87, 334–348. [Google Scholar] [CrossRef]
- Kuo, D.C.; de Groat, W.C. 1985. Primary afferent projections of the major splanchnic nerve to the spinal cord and gracile nucleus of the cat. J. Comp. Neurol. 1985, 231, 421–434. [Google Scholar] [CrossRef]
- Clerc, N. Afferent innervation of the lower oesophageal sphincter of the cat. An HRP study. J. Auton. Nerv. Syst. 1983, 9, 623–636. [Google Scholar] [CrossRef]
- Collman, P.I.; Tremblay, L.; Diamant, N.E. The distribution of spinal and vagal sensory neurons that innervate the esophagus of the cat. Gastroenterology 1992, 103, 817–822. [Google Scholar] [CrossRef]
- Giancola, F.; Gentilini, F.; Romagnoli, N.; Spadari, A.; Turba, M.E.; Giunta, M.; Sadeghinezhad, J.; Sorteni, C.; Chiocchetti, R. Extrinsic innervation of ileum and pelvic flexure of foals with ileocolonic aganglionosis. Cell Tissue Res. 2016, 366, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Brookes, S.J.; Spencer, N.J.; Costa, M.; Zagorodnyuk, V.P. Extrinsic primary afferent signalling in the gut. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Blackshaw, L.A.; Gebhart, G.F. The pharmacology of gastrointestinal nociceptive pathways. Curr. Opin. Pharmacol. 2002, 2, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.F.; Wedel, T.; Krammer, H.J.; Kühnel, W. Structural differences of the enteric nervous system in the cattle forestomach revealed by whole mount immunohistochemistry. Ann. Anat. 1998, 180, 393–400. [Google Scholar] [CrossRef]
- Furness, J.B.; Rivera, L.R.; Cho, H.J.; Bravo, D.M.; Callaghan, B. The gut as a sensory organ. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 729–740. [Google Scholar] [CrossRef]
- Jenkinson, D.M.; Sengupta, B.P.; Blackburn, P.S. The distribution of nerves, monoamine oxidase and cholinesterase in the skin of cattle. J. Anat. 1966, 100, 593–613. [Google Scholar]
- Young, R.L.; Page, A.J.; Cooper, N.J.; Frisby, C.L.; Blackshaw, L.A. Sensory and motor innervation of the crural diaphragm by the vagus nerves. Gastroenterology 2010, 138, 1091–1101.e1-5. [Google Scholar] [CrossRef]
- Person, R.J. Somatic and vagal afferent convergence on solitary tract neurons in cat: Electrophysiological characteristics. Neuroscience 1989, 30, 283–295. [Google Scholar] [CrossRef]
- Retamal, M.A.; Riquelme, M.A.; Stehberg, J.; Alcayaga, J. Connexin43 hemichannels in satellite glial cells, can they influence sensory neuron activity? Front. Mol. Neurosci. 2017, 10, 374. [Google Scholar] [CrossRef]
- Thalakoti, S.; Patil, V.V.; Damodaram, S.; Vause, C.V.; Langford, L.E.; Freeman, S.E.; Durham, P.L. Neuron-glia signaling in trigeminal ganglion: Implications for migraine pathology. Headache 2007, 7, 1008–1023. [Google Scholar] [CrossRef]
- Bráz, J.M.; Ackerman, L.; Basbaum, A.I. Sciatic nerve transection triggers release and intercellular transfer of a genetically expressed macromolecular tracer in dorsal root ganglia. J. Comp. Neurol. 2011, 519, 2648–2657. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.Y.; Cherkas, P.S.; Rosenthal, D.W.; Hanani, M. Dye coupling among satellite glial cells in mammalian dorsal root ganglia. Brain Res. 2005, 1036, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Chen, Y.; Zhang, X.; Li, G.-W.; Wang, C.; Huang, L.-Y.M. Neuronal soma-satellite glial cell interactions in sensory ganglia and the participation of purinergic receptors. Neuron Glia Biol. 2010, 6, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.Y.; Gu, Y.; Chen, Y. Communication between neuronal somata and satellite glial cells in sensory ganglia. Glia 2013, 61, 1571–1581. [Google Scholar] [CrossRef]
- Kung, L.H.; Gong, K.; Adedoyin, M.; Ng, J.; Bhargava, A.; Ohara, P.T.; Jasmin, L. Evidence for glutamate as a neuroglial transmitter within sensory ganglia. PLoS ONE 2013, 8, e68312. [Google Scholar] [CrossRef]
- Zhang, F.C.; Weng, R.X.; Li, D.; Li, Y.C.; Dai, X.X.; Hu, S.; Sun, Q.; Li, R.; Xu, G.Y. A vagus nerve dominant tetra-synaptic as-cending pathway for gastric pain processing. Nat. Commun. 2024, 15, 9824. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiocchetti, R.; Pisoni, L.; Joechler, M.; Cancellieri, A.; Giancola, F.; Galiazzo, G.; Salamanca, G.; Cunha, R.Z.; Gentile, A. Cutaneous Allodynia of the Withers in Cattle: An Experimental In Vivo Neuroanatomical Preliminary Investigation of the Dichotomizing Sensory Neurons Projecting into the Reticulum and Skin of the Withers—A Case Study on Two Calves. Animals 2025, 15, 1689. https://doi.org/10.3390/ani15121689
Chiocchetti R, Pisoni L, Joechler M, Cancellieri A, Giancola F, Galiazzo G, Salamanca G, Cunha RZ, Gentile A. Cutaneous Allodynia of the Withers in Cattle: An Experimental In Vivo Neuroanatomical Preliminary Investigation of the Dichotomizing Sensory Neurons Projecting into the Reticulum and Skin of the Withers—A Case Study on Two Calves. Animals. 2025; 15(12):1689. https://doi.org/10.3390/ani15121689
Chicago/Turabian StyleChiocchetti, Roberto, Luciano Pisoni, Monika Joechler, Adele Cancellieri, Fiorella Giancola, Giorgia Galiazzo, Giulia Salamanca, Rodrigo Zamith Cunha, and Arcangelo Gentile. 2025. "Cutaneous Allodynia of the Withers in Cattle: An Experimental In Vivo Neuroanatomical Preliminary Investigation of the Dichotomizing Sensory Neurons Projecting into the Reticulum and Skin of the Withers—A Case Study on Two Calves" Animals 15, no. 12: 1689. https://doi.org/10.3390/ani15121689
APA StyleChiocchetti, R., Pisoni, L., Joechler, M., Cancellieri, A., Giancola, F., Galiazzo, G., Salamanca, G., Cunha, R. Z., & Gentile, A. (2025). Cutaneous Allodynia of the Withers in Cattle: An Experimental In Vivo Neuroanatomical Preliminary Investigation of the Dichotomizing Sensory Neurons Projecting into the Reticulum and Skin of the Withers—A Case Study on Two Calves. Animals, 15(12), 1689. https://doi.org/10.3390/ani15121689