

Acute Effect of Short-Term Benzocaine Anesthesia on the Skin Mucus Microbiome of Atlantic salmon (Salmo salar)



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site, Animals and Feeding
2.2. Salmon Skin Mucus Sampling and DNA Extraction
2.3. Water Sampling and DNA Extraction
2.4. Next-Generation Sequencing Analysis
2.5. Data Analyses
3. Results and Discussion
3.1. Effect of Anesthesia in the Skin Mucus Microbiome of Atlantic salmon
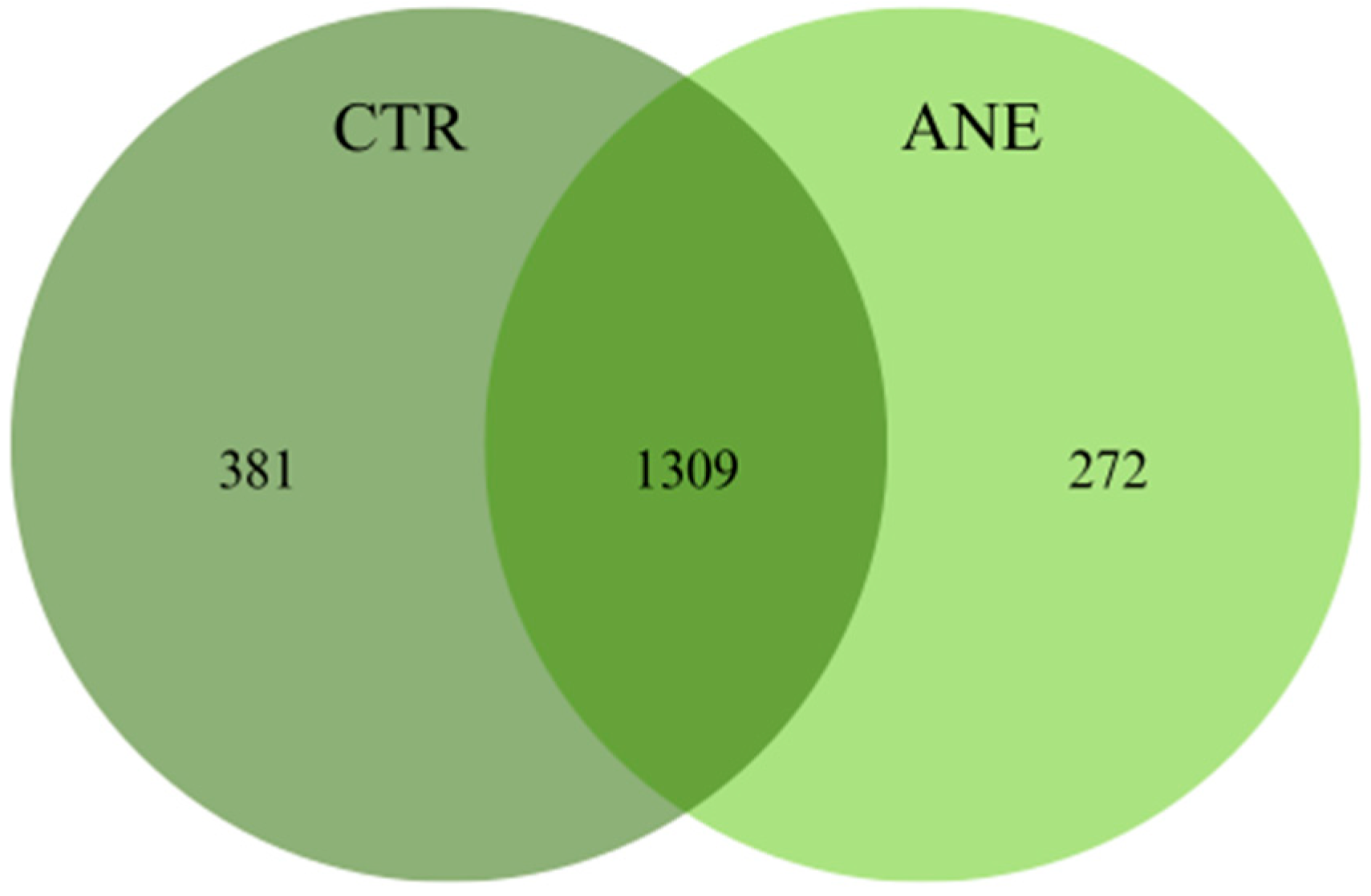
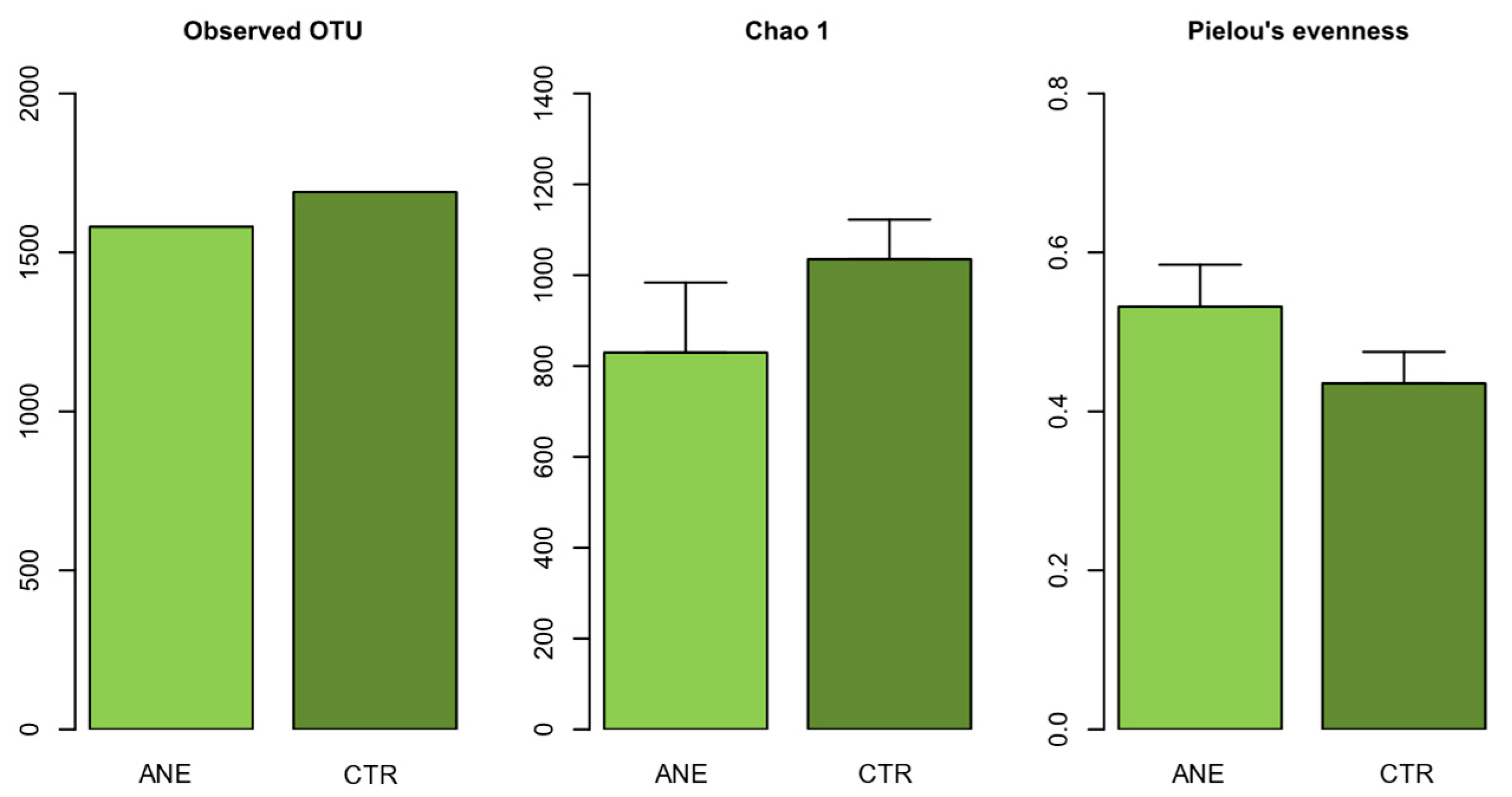
3.1.1. Diversity and Community Structure of Bacterial Composition
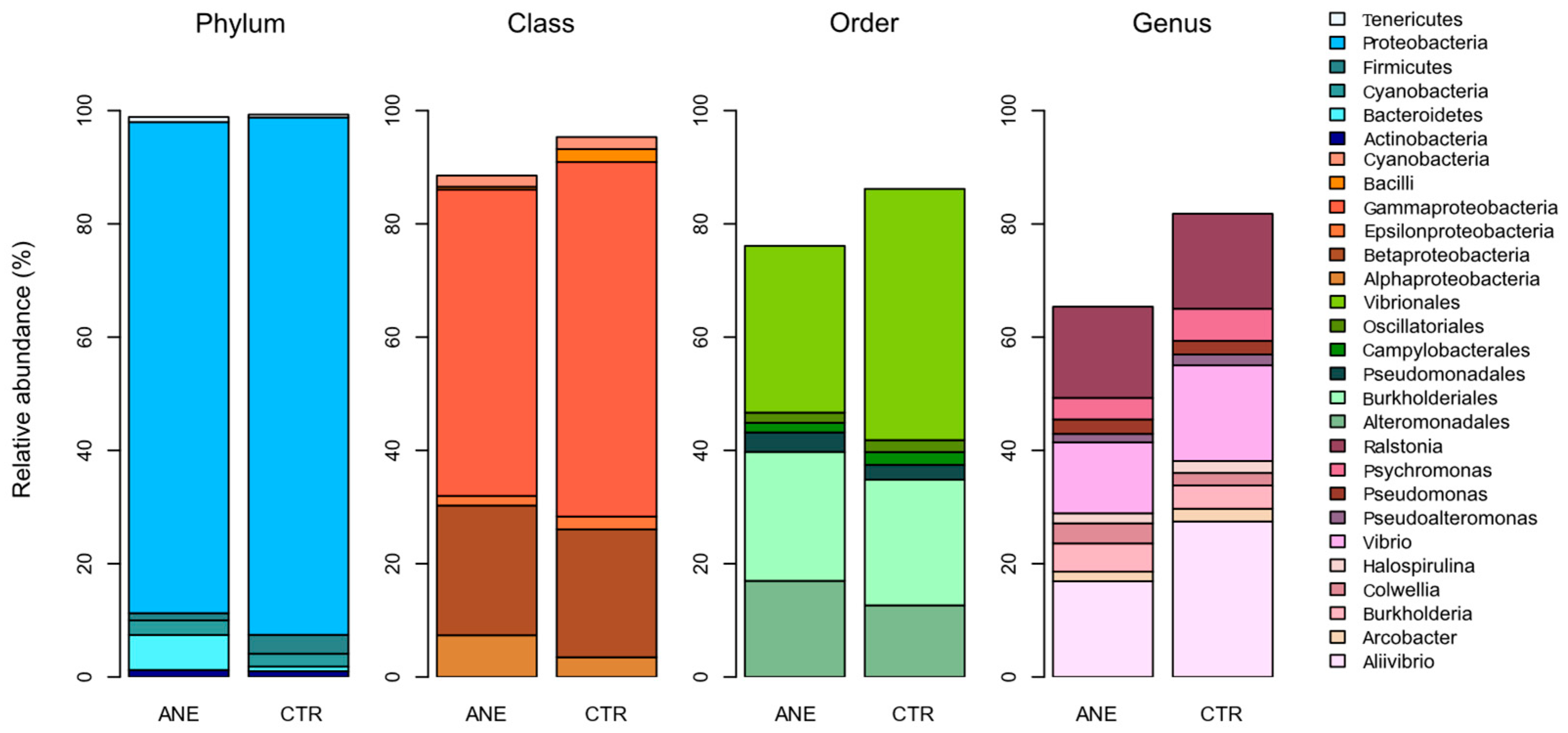
3.1.2. Taxonomic Profiling of Bacterial Composition
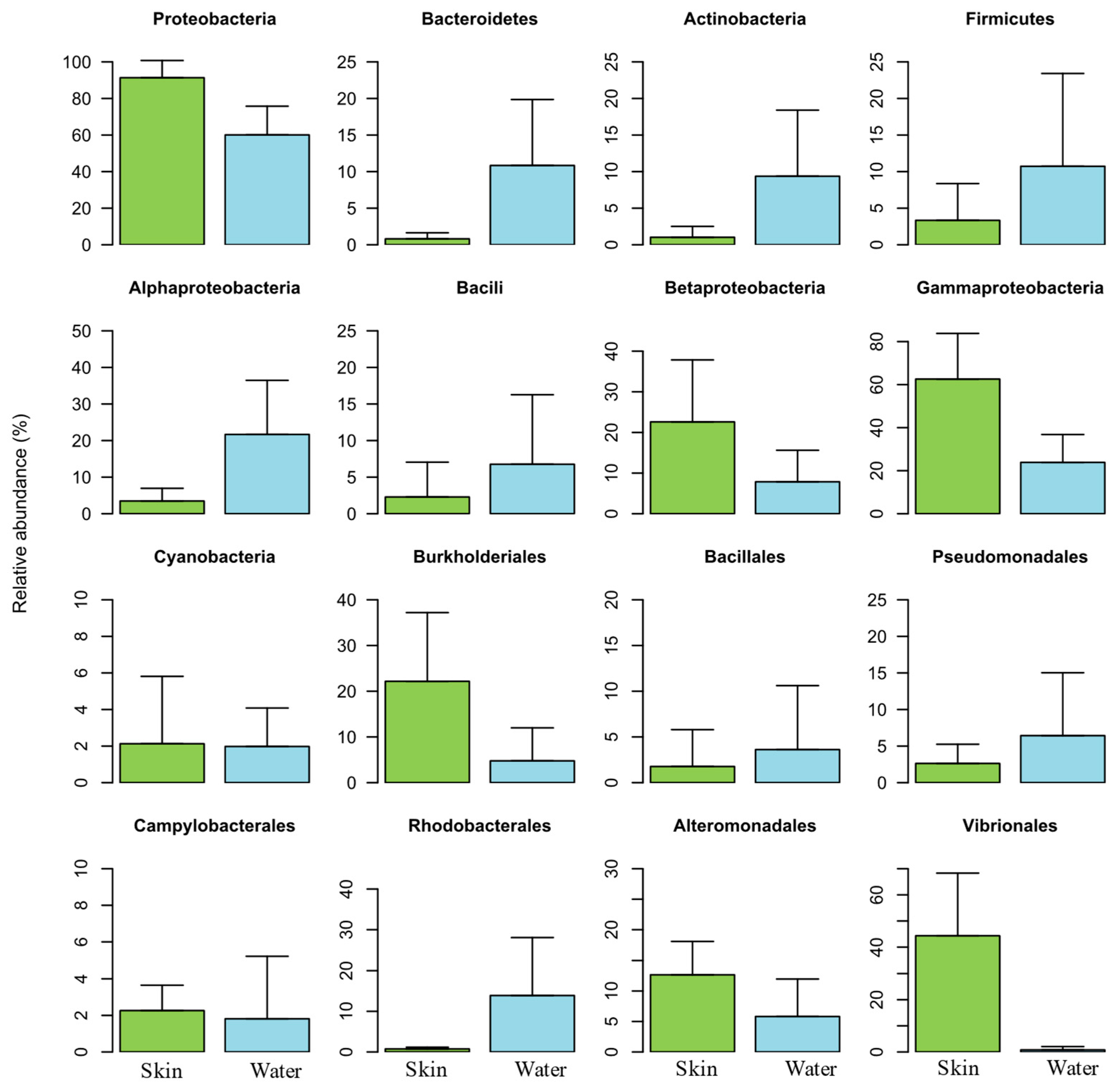
3.2. Comparison of Skin Mucus Microbiome of Atlantic salmon and Rearing Water
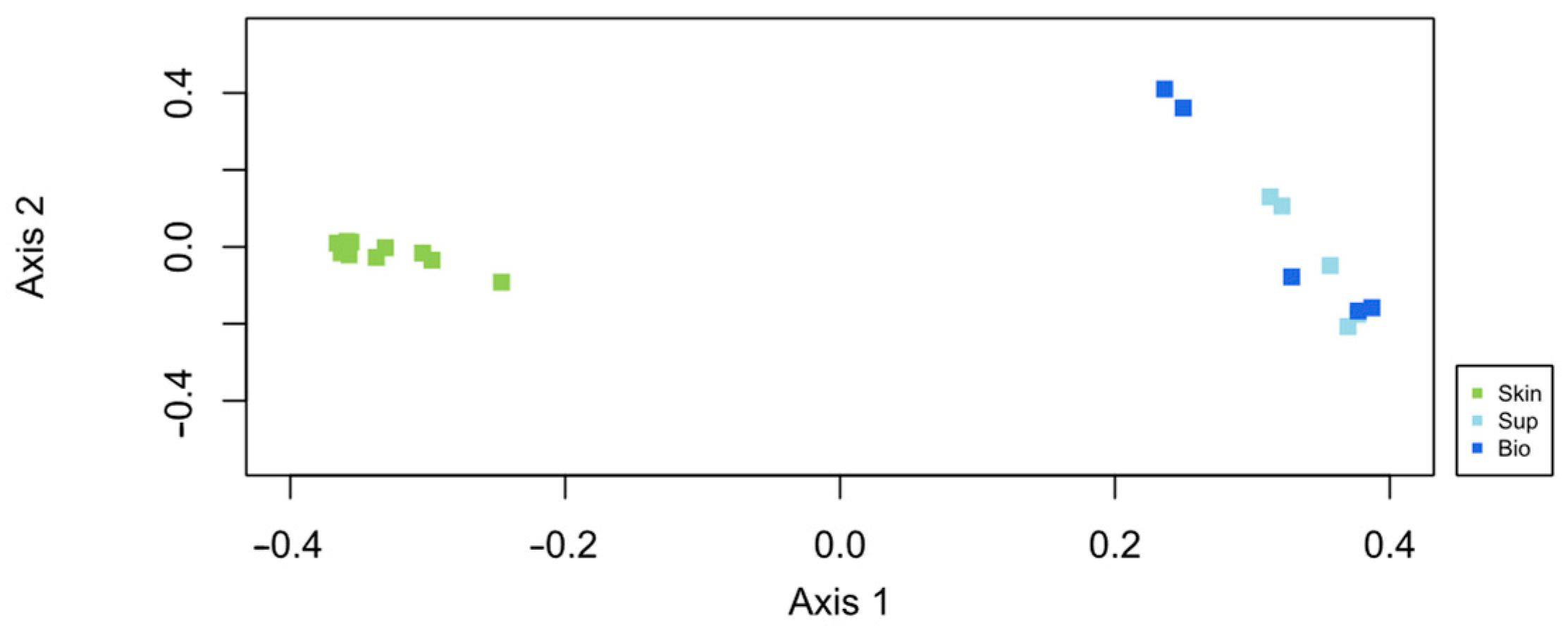
3.2.1. Overall Assessment of Bacterial Composition
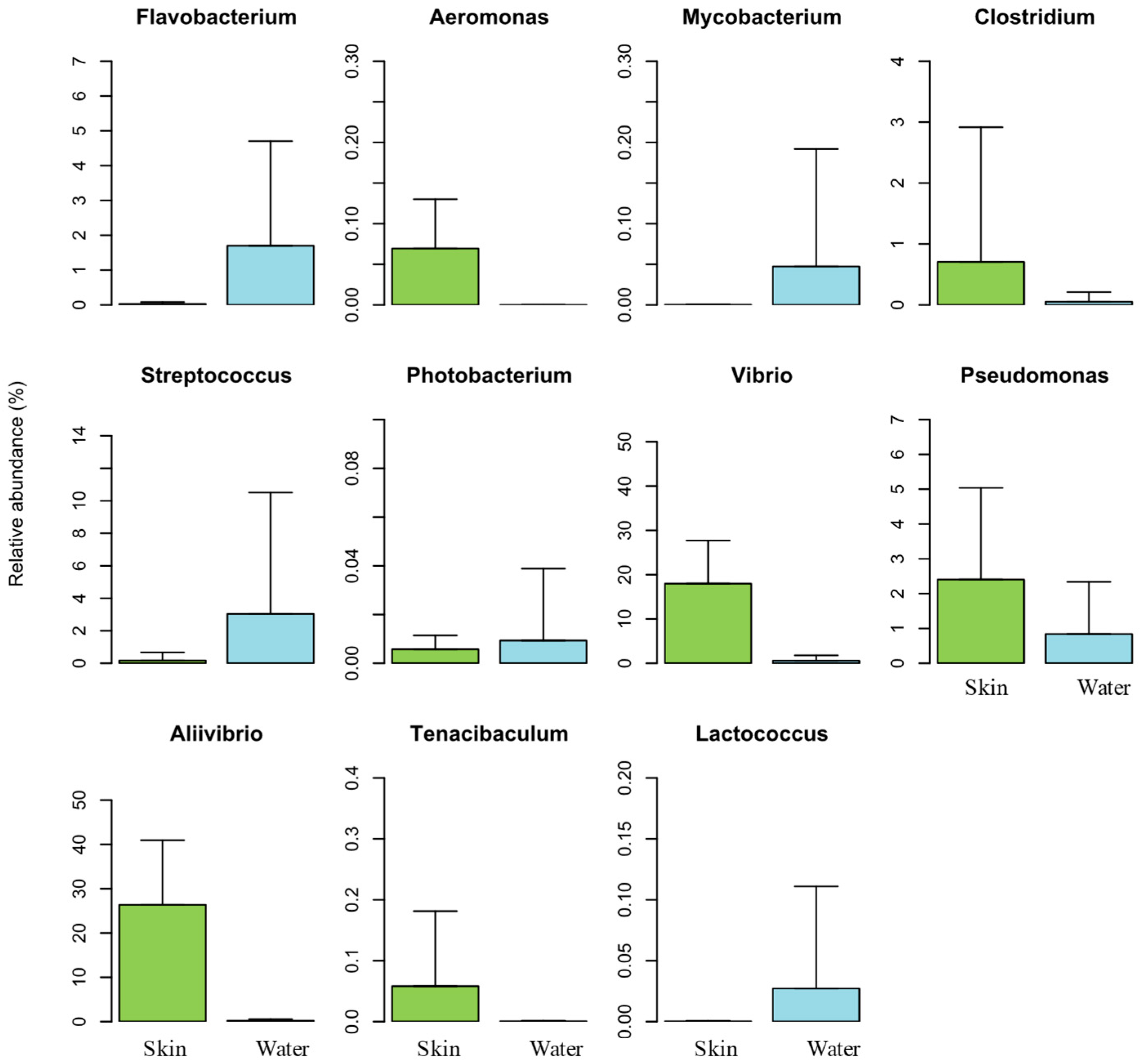
3.2.2. Taxonomic Bacterial Composition
3.2.3. Composition Analysis of Dominant OTUs in Salmon Skin Mucus
3.2.4. Core Microbiome Composition of Atlantic salmon
3.2.5. Bacterial Community Profile of Rearing Water
3.2.6. Potential Pathogens Detected in the Skin Mucus Microbiome of Atlantic salmon and in Rearing Water
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Esteban, M.Á. An Overview of the Immunological Defenses in Fish Skin. ISRN Immunol. 2012, 2012, 853470. [Google Scholar] [CrossRef]
- Benhamed, S.; Guardiola, F.A.; Mars, M.; Esteban, M.Á. Pathogen Bacteria Adhesion to Skin Mucus of Fishes. Vet. Microbiol. 2014, 171, 1–12. [Google Scholar] [CrossRef]
- Tiralongo, F.; Messina, G.; Lombardo, B.M.; Longhitano, L.; Volti, G.L.; Tibullo, D. Skin Mucus of Marine Fish as a Source for the Development of Antimicrobial Agents. Front. Mar. Sci. 2020, 7, 541853. [Google Scholar] [CrossRef]
- Dash, S.; Das, S.K.; Samal, J.; Thatoi, H.N. Epidermal Mucus, a Major Determinant in Fish Health: A Review. Iran. J. Vet. Res. 2018, 19, 72–81. [Google Scholar]
- Wang, L.-C.; Chen, L.-H.; Chiu, Y.-C.; Liou, C.-Y.; Chen, H.-C.; Lu, C.-Y.; Chen, J.-L. Teleost Skin Microbiome: An Intimate Interplay between the Environment and the Host Immunity. Fish Shellfish Immunol. 2023, 139, 108869. [Google Scholar] [CrossRef]
- de Bruijn, I.; Liu, Y.; Wiegertjes, G.F.; Raaijmakers, J.M. Exploring Fish Microbial Communities to Mitigate Emerging Diseases in Aquaculture. FEMS Microbiol. Ecol. 2018, 94, fix161. [Google Scholar] [CrossRef]
- Xavier, R.; Severino, R.; Silva, S.M. Signatures of Dysbiosis in Fish Microbiomes in the Context of Aquaculture. Rev. Aquac. 2024, 16, 706–731. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost Microbiomes: The State of the Art in Their Characterization, Manipulation and Importance in Aquaculture and Fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar] [CrossRef]
- Lorgen-Ritchie, M.; Clarkson, M.; Chalmers, L.; Taylor, J.F.; Migaud, H.; Martin, S.A.M. Temporal Changes in Skin and Gill Microbiomes of Atlantic Salmon in a Recirculating Aquaculture System—Why Do They Matter? Aquaculture 2022, 558, 738352. [Google Scholar] [CrossRef]
- Barata, M.; Soares, F.; Aragão, C.; Almeida, A.C.; Pousão-Ferreira, P.; Ribeiro, L. Efficiency of 2-Phenoxyethanol and Clove Oil for Reducing Handling Stress in Reared Meagre, Argyrosomus regius (Pisces: Sciaenidae). J. World Aquac. Soc. 2016, 47, 82–92. [Google Scholar] [CrossRef]
- Bowker, J.D.; Trushenski, J.T.; Bowman, M. Efficacy of Eugenol to Lightly Sedate Freshwater Salmonids for an Extended Time Period. N. Am. J. Aquac. 2019, 81, 40–46. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, X.-W.; Ding, H.-T.; Dong, X.-J.; Zhang, J.-J.; Zheng, Y.-C.; Chen, X.-N.; Cheng, H.-L.; Ding, Z.-J.; Xu, J.-H. Effects of Tricaine Methanesulfonate (MS-222) on Sedation and Responses of Yellow Catfish (Pelteobagrus fulvidraco) Subjected to Simulated Transportation Stress. Aquaculture 2022, 549, 737789. [Google Scholar] [CrossRef]
- Sadoul, B.; Geffroy, B. Measuring Cortisol, the Major Stress Hormone in Fishes. J. Fish Biol. 2019, 94, 540–555. [Google Scholar] [CrossRef]
- Cao, X.; Wang, Y.; Yu, N.; Le, Q.; Hu, J.; Yang, Y.; Kuang, S.; Zhang, M.; Sun, Y.; Gu, W.; et al. Transcriptome Analysis Reveals the Influence of Anaesthetic Stress on the Immune System of Crucian Carp (Carassius auratus) under the Process of Treatment and Low Concentration Transport by MS-222 and Eugenol. Aquac. Res. 2019, 50, 3138–3153. [Google Scholar] [CrossRef]
- Cho, G.K.; Heath, D.D. Comparison of Tricaine Methanesulphonate (MS222) and Clove Oil Anaesthesia Effects on the Physiology of Juvenile Chinook Salmon Oncorhynchus tshawytscha (Walbaum). Aquac. Res. 2000, 31, 537–546. [Google Scholar] [CrossRef]
- Zahl, I.H.; Samuelsen, O.; Kiessling, A. Anaesthesia of Farmed Fish: Implications for Welfare. Fish Physiol. Biochem. 2012, 38, 201–218. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2022: Towards Blue Transformation; FAO: Rome, Italy, 2022; p. 223. [Google Scholar]
- Føre, H.M.; Thorvaldsen, T.; Osmundsen, T.C.; Asche, F.; Tveterås, R.; Fagertun, J.T.; Bjelland, H. V Technological Innovations Promoting Sustainable Salmon (Salmo salar) Aquaculture in Norway. Aquac. Rep. 2022, 24, 101115. [Google Scholar] [CrossRef]
- Martins, P.; Cleary, D.F.R.; Pires, A.C.C.; Rodrigues, A.M.; Quintino, V.; Calado, R.; Gomes, N.C.M. Molecular Analysis of Bacterial Communities and Detection of Potential Pathogens in a Recirculating Aquaculture System for Scophthalmus maximus and Solea senegalensis. PLoS ONE 2013, 8, e80847. [Google Scholar] [CrossRef]
- Rosado, D.; Canada, P.; Marques Silva, S.; Ribeiro, N.; Diniz, P.; Xavier, R. Disruption of the Skin, Gill, and Gut Mucosae Microbiome of Gilthead Seabream Fingerlings after Bacterial Infection and Antibiotic Treatment. FEMS Microbes 2023, 4, xtad011. [Google Scholar] [CrossRef]
- Yue, G.H.; Wang, L. Current Status of Genome Sequencing and Its Applications in Aquaculture. Aquaculture 2017, 468, 337–347. [Google Scholar] [CrossRef]
- Pimentel, T.; Marcelino, J.; Ricardo, F.; Soares, A.M.V.M.; Calado, R. Bacterial Communities 16S rDNA Fingerprinting as a Potential Tracing Tool for Cultured Seabass Dicentrarchus labrax. Sci. Rep. 2017, 7, 11862. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global Patterns of 16S RRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4516–4522. [Google Scholar] [CrossRef]
- Kakakhel, M.A.; Bibi, N.; Mahboub, H.H.; Wu, F.; Sajjad, W.; Din, S.Z.U.; Hefny, A.A.; Wang, W. Influence of Biosynthesized Nanoparticles Exposure on Mortality, Residual Deposition, and Intestinal Bacterial Dysbiosis in Cyprinus carpio. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2023, 263, 109473. [Google Scholar] [CrossRef]
- Wang, Q.; Cole, J.R. Updated RDP Taxonomy and RDP Classifier for More Accurate Taxonomic Classification. Microbiol. Resour. Announc. 2024, 13, e0106323. [Google Scholar] [CrossRef]
- Noble, C.; Gismervik, S.; Iversen, M.; Kolarevic, J.; Nilsson, J.; Stien, L.; Turnbull, J. Welfare Indicators for Farmed Atlantic Salmon: Tools for Assessing Fish Welfare; Nofima: Tromsø, Norway, 2018; ISBN 978-82-8296-556-9. [Google Scholar]
- Lokesh, J.; Kiron, V. Transition from Freshwater to Seawater Reshapes the Skin-Associated Microbiota of Atlantic Salmon. Sci. Rep. 2016, 6, 19707. [Google Scholar] [CrossRef]
- Minniti, G.; Hagen, L.H.; Porcellato, D.; Jørgensen, S.M.; Pope, P.B.; Vaaje-Kolstad, G. The Skin-Mucus Microbial Community of Farmed Atlantic Salmon (Salmo salar). Front. Microbiol. 2017, 8, 2043. [Google Scholar] [CrossRef]
- Rud, I.; Kolarevic, J.; Holan, A.B.; Berget, I.; Calabrese, S.; Terjesen, B.F. Deep-Sequencing of the Bacterial Microbiota in Commercial-Scale Recirculating and Semi-Closed Aquaculture Systems for Atlantic Salmon Post-Smolt Production. Aquac. Eng. 2017, 78, 50–62. [Google Scholar] [CrossRef]
- Kearns, P.J.; Bowen, J.L.; Tlusty, M.F. The Skin Microbiome of Cow-Nose Rays (Rhinoptera bonasus) in an Aquarium Touch-Tank Exhibit. Zoo Biol. 2017, 36, 226–230. [Google Scholar] [CrossRef]
- Lowrey, L.; Woodhams, D.C.; Tacchi, L.; Salinas, I. Topographical Mapping of the Rainbow Trout (Oncorhynchus mykiss) Microbiome Reveals a Diverse Bacterial Community with Antifungal Properties in the Skin. Appl. Environ. Microbiol. 2015, 81, 6915–6925. [Google Scholar] [CrossRef]
- Flemer, B.; Gulati, S.; Bergna, A.; Rändler, M.; Cernava, T.; Witzel, K.; Berg, G.; Grosch, R. Biotic and Abiotic Stress Factors Induce Microbiome Shifts and Enrichment of Distinct Beneficial Bacteria in Tomato Roots. Phytobiomes J. 2022, 6, 276–289. [Google Scholar] [CrossRef]
- Rubin, M.A.; Leff, L.G. Nutrients and Other Abiotic Factors Affecting Bacterial Communities in an Ohio River (USA). Microb. Ecol. 2007, 54, 374–383. [Google Scholar] [CrossRef]
- Karlsen, C.; Vanberg, C.; Mikkelsen, H.; Sørum, H. Co-Infection of Atlantic Salmon (Salmo salar), by Moritella viscosa and Aliivibrio wodanis, Development of Disease and Host Colonization. Vet. Microbiol. 2014, 171, 112–121. [Google Scholar] [CrossRef]
- Lunder, T.; Sørum, H.; Holstad, G.; Steigerwalt, A.G.; Mowinckel, P.; Brenner, D.J. Phenotypic and Genotypic Characterization of Vibrio viscosus Sp. Nov. and Vibrio wodanis sp. Nov. Isolated from Atlantic Salmon (Salmo salar) with “Winter Ulcer”. Int. J. Syst. Evol. Microbiol. 2000, 50, 427–450. [Google Scholar] [CrossRef]
- Klakegg, Ø.; Myhren, S.; Juell, R.A.; Aase, M.; Salonius, K.; Sørum, H. Improved Health and Better Survival of Farmed Lumpfish (Cyclopterus lumpus) after a Probiotic Bath with Two Probiotic Strains of Aliivibrio. Aquaculture 2020, 518, 734810. [Google Scholar] [CrossRef]
- Godoy, M.; Coca, Y.; Suárez, R.; de Oca, M.M.; Bledsoe, J.W.; Burbulis, I.; Caro, D.; Pontigo, J.P.; Maracaja-Coutinho, V.; Arias-Carrasco, R.; et al. Salmo Salar Skin and Gill Microbiome during Piscirickettsia salmonis Infection. Animals 2023, 14, 97. [Google Scholar] [CrossRef]
- Viver, T.; Ruiz, A.; Bertomeu, E.; Martorell-Barceló, M.; Urdiain, M.; Grau, A.; Signaroli, M.; Barcelo-Serra, M.; Aspillaga, E.; Pons, A.; et al. Food Determines Ephemerous and Non-Stable Gut Microbiome Communities in Juvenile Wild and Farmed Mediterranean Fish. Sci. Total Environ. 2023, 889, 164080. [Google Scholar] [CrossRef]
- Cerezo-Ortega, I.M.; Di Zeo-Sánchez, D.E.; García-Márquez, J.; Ruiz-Jarabo, I.; Sáez-Casado, M.I.; Balebona, M.C.; Moriñigo, M.A.; Tapia-Paniagua, S.T. Microbiota Composition and Intestinal Integrity Remain Unaltered after the Inclusion of Hydrolysed Nannochloropsis gaditana in Sparus aurata Diet. Sci. Rep. 2021, 11, 18779. [Google Scholar] [CrossRef]
- Keating, C.; Bolton-Warberg, M.; Hinchcliffe, J.; Davies, R.; Whelan, S.; Wan, A.H.L.; Fitzgerald, R.D.; Davies, S.J.; Ijaz, U.Z.; Smith, C.J. Temporal Changes in the Gut Microbiota in Farmed Atlantic Cod (Gadus Morhua) Outweigh the Response to Diet Supplementation with Macroalgae. Anim. Microbiome 2021, 3, 7. [Google Scholar] [CrossRef]
- Zrihan, S.; Itay, P.; Kroin, Y.; Davidovich, N.; Wosnick, N.; Tchernov, D.; Koh, X.P.; Lau, S.C.K.; Morick, D. Monitoring Fish Bacterial Pathogens of Wild Fish Species From the South China Sea by Applying Next-Generation Sequencing on Gill Tissue. J. Fish. Dis. 2025, 48, e14050. [Google Scholar] [CrossRef]
- Razavi, B.M.; Fazly Bazzaz, B.S. A Review and New Insights to Antimicrobial Action of Local Anesthetics. Eur. J. Clin. Microbiol. Infect Dis. 2019, 38, 991–1002. [Google Scholar] [CrossRef]
- Morrow, M.E.; Berry, C.W. Antimicrobial Properties of Topical Anesthetic Liquids Containing Lidocaine or Benzocaine. Anesth. Prog. 1988, 35, 9–13. [Google Scholar] [PubMed]
- Shephard, K.L. Functions for Fish Mucus. Rev. Fish Biol. Fish. 1994, 4, 401–429. [Google Scholar] [CrossRef]
- Gomez, D.; Sunyer, J.O.; Salinas, I. The Mucosal Immune System of Fish: The Evolution of Tolerating Commensals While Fighting Pathogens. Fish Shellfish Immunol. 2013, 35, 1729–1739. [Google Scholar] [CrossRef]
- Dehler, C.E.; Secombes, C.J.; Martin, S.A.M. Environmental and Physiological Factors Shape the Gut Microbiota of Atlantic Salmon Parr (Salmo salar L.). Aquaculture 2017, 467, 149–157. [Google Scholar] [CrossRef]
- Liu, H.; Guo, X.; Gooneratne, R.; Lai, R.; Zeng, C.; Zhan, F.; Wang, W. The Gut Microbiome and Degradation Enzyme Activity of Wild Freshwater Fishes Influenced by Their Trophic Levels. Sci. Rep. 2016, 6, 24340. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; McGinnity, P.; Dionne, M.; Letourneau, J.; Thonier, F.; Carvalho, G.R.; Creer, S.; Derome, N. The Biogeography of the Atlantic Salmon (Salmo salar) Gut Microbiome. ISME J. 2016, 10, 1280–1284. [Google Scholar] [CrossRef]
- Webster, T.M.U.; Consuegra, S.; Hitchings, M.; de Leaniz, C.G. Interpopulation Variation in the Atlantic Salmon Microbiome Reflects Environmental and Genetic Diversity. Appl. Environ. Microbiol. 2018, 84, e00691-18. [Google Scholar] [CrossRef]
- Malick, R.C.; Bera, A.K.; Chowdhury, H.; Bhattacharya, M.; Abdulla, T.; Swain, H.S.; Baitha, R.; Kumar, V.; Das, B.K. Identification and Pathogenicity Study of Emerging Fish Pathogens Acinetobacter junii and Acinetobacter pittii Recovered from a Disease Outbreak in Labeo catla (Hamilton, 1822) and Hypophthalmichthys molitrix (Valenciennes, 1844) of Freshwater Wetland in West Bengal, India. Aquac. Res. 2020, 51, 2410–2420. [Google Scholar] [CrossRef]
- Minniti, G.; Sandve, S.R.; Padra, J.T.; Hagen, L.H.; Lindén, S.; Pope, P.B.; Arntzen, M.Ø.; Vaaje-Kolstad, G. The Farmed Atlantic Salmon (Salmo salar) Skin–Mucus Proteome and Its Nutrient Potential for the Resident Bacterial Community. Genes 2019, 10, 515. [Google Scholar] [CrossRef]
- Benediktsdo, E.; Verdonck, L.; Spro, C.; Helgason, S.; Swings, J. Characterization of Vibrio viscosus and Vibrio wodanis Isolated at Different Geographical Locations: A Proposal for Reclassification of Vibrio viscosus as Moritella viscosa comb. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 479–488. [Google Scholar] [CrossRef]
- Aranda, C.; Valenzuela, C.; Barrientos, J.; Paredes, J.; Leal, P.; Maldonado, M.; Godoy, F.; Osorio, C. Bacteriostatic Anti-Vibrio parahaemolyticus Activity of Pseudoalteromonas sp. Strains DIT09, DIT44 and DIT46 Isolated from Southern Chilean Intertidal Perumytilus purpuratus. World J. Microbiol. Biotechnol. 2012, 28, 2365–2374. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Gupta, P.; Dhawan, A. Paenibacillus Polymyxa as a Water Additive Improved Immune Response of Cyprinus carpio and Disease Resistance against Aeromonas hydrophila. Aquac. Rep. 2016, 4, 86–92. [Google Scholar] [CrossRef]
- Tarnecki, A.M.; Brennan, N.P.; Schloesser, R.W.; Rhody, N.R. Shifts in the Skin-Associated Microbiota of Hatchery-Reared Common Snook Centropomus undecimalis During Acclimation to the Wild. Microb. Ecol. 2019, 77, 770–781. [Google Scholar] [CrossRef]
- Urbanczyk, H.; Ast, J.C.; Higgins, M.J.; Carson, J.; Dunlap, P.V. Reclassification of Vibrio fischeri, Vibrio logei, Vibrio salmonicida and Vibrio wodanis as Aliivibrio fischeri gen. nov., comb. nov., Aliivibrio logei comb. nov., Aliivibrio salmonicida comb. nov. and Aliivibrio wodanis comb. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 2823–2829. [Google Scholar] [CrossRef]
- Gauger, E.; Smolowitz, R.; Uhlinger, K.; Casey, J.; Gómez-Chiarri, M. Vibrio harveyi and Other Bacterial Pathogens in Cultured Summer Flounder, Paralichthys dentatus. Aquaculture 2006, 260, 10–20. [Google Scholar] [CrossRef]
- Zhang, X.H.; Austin, B. Pathogenicity of Vibrio harveyi to Salmonids. J. Fish. Dis. 2000, 23, 93–102. [Google Scholar] [CrossRef]
- Zorrilla, I.; Arijo, S.; Chabrillon, M.; Diaz, P.; Martinez-Manzanares, E.; Balebona, M.; Morinigo, M. Vibrio Species Isolated from Diseased Farmed Sole, Solea senegalensis (Kaup), and Evaluation of the Potential Virulence Role of Their Extracellular Products. J. Fish Dis. 2003, 26, 103–108. [Google Scholar] [CrossRef]
- Balebona, M.C.; Moriñigo, M.A.; Faris, A.; Krovacek, K.; Månsson, I.; Bordas, M.A.; Borrego, J.J. Influence of Salinity and pH on the Adhesion of Pathogenic Vibrio Strains to Sparus aurata Skin Mucus. Aquaculture 1995, 132, 113–120. [Google Scholar] [CrossRef]
- Bordas, M.A.; Balebona, M.C.; Zorrilla, I.; Borrego, J.J.; Moriñigo, M.A.; Moriñigo, M. Kinetics of Adhesion of Selected Fish-Pathogenic Vibrio Strains to Skin Mucus of Gilt-Head Sea Bream (Sparus aurata L.). Appl. Environ. Microbiol. 1996, 62, 3650–3654. [Google Scholar] [CrossRef]
- Joly-Guillou, M. Clinical Impact and Pathogenicity of Acinetobacter. Clin. Microbiol. Infect. 2005, 11, 868–873. [Google Scholar] [CrossRef]
- Buchan, A.; González, J.M.; Moran, M.A. Overview of the Marine Roseobacter Lineage. Appl. Environ. Microbiol. 2005, 71, 5665–5677. [Google Scholar] [CrossRef] [PubMed]
- D’Alvise, P.; Melchiorsen, J.; Porsby, C.; Nielsen, K.; Gram, L. Inactivation of Vibrio anguillarum by Attached and Planktonic Roseobacter Cells. Appl. Environ. Microbiol. 2010, 76, 2366–2370. [Google Scholar] [CrossRef]
- Hjelm, M.; Bergh, Ø.; Riaza, A.; Nielsen, J.; Melchiorsen, J.; Jensen, S.; Duncan, H.; Ahrens, P.; Birkbeck, H.; Gram, L. Selection and Identification of Autochthonous Potential Probiotic Bacteria from Turbot Larvae (Scophthalmus maximus) Rearing Units. Syst. Appl. Microbiol. 2004, 27, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Martins, P.; Coelho, F.J.R.C.; Cleary, D.F.R.; Pires, A.C.C.; Marques, B.; Rodrigues, A.M.; Quintino, V.; Gomes, N.C.M. Seasonal Patterns of Bacterioplankton Composition in a Semi-Intensive European Seabass (Dicentrarchus labrax) Aquaculture System. Aquaculture 2018, 490, 240–250. [Google Scholar] [CrossRef]
- Salati, F. Fish Diseases and Disorders. In Viral, Bacterial and Fungal Infections; Woo, P.T.K., Bruno, D.W., Eds.; CABI: Oxfordshire, UK, 2011; Volume 3, ISBN 978-1-84593-554-2. [Google Scholar]
- Ayyat, M.S.; Labib, H.M.; Mahmoud, H.K. A Probiotic Cocktail as a Growth Promoter in Nile Tilapia (Oreochromis niloticus). J. Appl. Aquac. 2014, 26, 208–215. [Google Scholar] [CrossRef]
- Haenen, O. Major Bacterial Diseases Affecting Aquaculture. In Proceedings of the Aquatic AMR Workshop 1, Mangalore, India, 10–11 April 2017. [Google Scholar]
- O’Brien, D.; Mooney, J.; Ryan, D.; Powell, E.; Hiney, M.; Smith, P.R.; Powell, R. Detection of Aeromonas salmonicida, Causal Agent of Furunculosis in Salmonid Fish, from the Tank Effluent of Hatchery-Reared Atlantic Salmon Smolts. Appl. Environ. Microbiol. 1994, 60, 3874–3877. [Google Scholar] [CrossRef]
- Bruno, D.W.; Hastings, T.S.; Wootten, R. Outbreak of a Cold Water Vibriosis in Atlantic Salmon in Scotland. Bull. Eur. Assoc. Fish Pathol. 1985, 5, 62–63. [Google Scholar]
- Kashulin, A.; Seredkina, N.; Sørum, H. Cold-Water Vibriosis. The Current Status of Knowledge. J. Fish Dis. 2017, 40, 119–126. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OTU | Genus Identification | Average Abundance ANE | Average Abundance CTR | Ind. % | Cum.% |
|---|---|---|---|---|---|
| OTU 3097 | Aliivibrio | 16,909.20 | 55,259.30 | 23.68 | 23.68 |
| OTU 3110 | Vibrio | 10,091.20 | 24,032.70 | 10.54 | 34.22 |
| OTU 2 | Ralstonia | 9092.10 | 21,759.30 | 9.64 | 43.86 |
| OTU 427 | Psychromonas | 3147.50 | 9258.50 | 4.18 | 48.04 |
| OTU 1 | Vibrio | 2505.50 | 8468.00 | 3.70 | 51.73 |
| OTU 4 | Burkholderia | 2840.30 | 4570.80 | 1.93 | 53.66 |
| OTU 14 | Bacillus | 2.10 | 3257.40 | 1.68 | 55.34 |
| OTU 8 | Pseudomonas | 2106.20 | 2732.90 | 1.67 | 57.01 |
| OTU 7 | Pseudoalteromonas | 1265.70 | 3465.60 | 1.62 | 58.63 |
| OTU 30 | Tenacibaculum | 2054.60 | 135.90 | 1.44 | 60.07 |
| OTU 16 | Altererythrobacter | 1069.20 | 1768.30 | 1.21 | 61.28 |
| OTU 32 | Colwellia | 2604.90 | 1908.80 | 1.20 | 62.48 |
| OTU 19 | Massilia | 1599.10 | 984.50 | 1.08 | 63.57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, P.; Pimentel, T.; Ribeiro, N.; Calado, R. Acute Effect of Short-Term Benzocaine Anesthesia on the Skin Mucus Microbiome of Atlantic salmon (Salmo salar). Animals 2025, 15, 1566. https://doi.org/10.3390/ani15111566
Martins P, Pimentel T, Ribeiro N, Calado R. Acute Effect of Short-Term Benzocaine Anesthesia on the Skin Mucus Microbiome of Atlantic salmon (Salmo salar). Animals. 2025; 15(11):1566. https://doi.org/10.3390/ani15111566
Chicago/Turabian StyleMartins, Patrícia, Tânia Pimentel, Nuno Ribeiro, and Ricardo Calado. 2025. "Acute Effect of Short-Term Benzocaine Anesthesia on the Skin Mucus Microbiome of Atlantic salmon (Salmo salar)" Animals 15, no. 11: 1566. https://doi.org/10.3390/ani15111566
APA StyleMartins, P., Pimentel, T., Ribeiro, N., & Calado, R. (2025). Acute Effect of Short-Term Benzocaine Anesthesia on the Skin Mucus Microbiome of Atlantic salmon (Salmo salar). Animals, 15(11), 1566. https://doi.org/10.3390/ani15111566