
Preliminary Study of Differential circRNA Expression and Investigation of circRNA–miRNA–mRNA Competitive Endogenous Network in Rumen Acidosis of Holstein Cattle


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
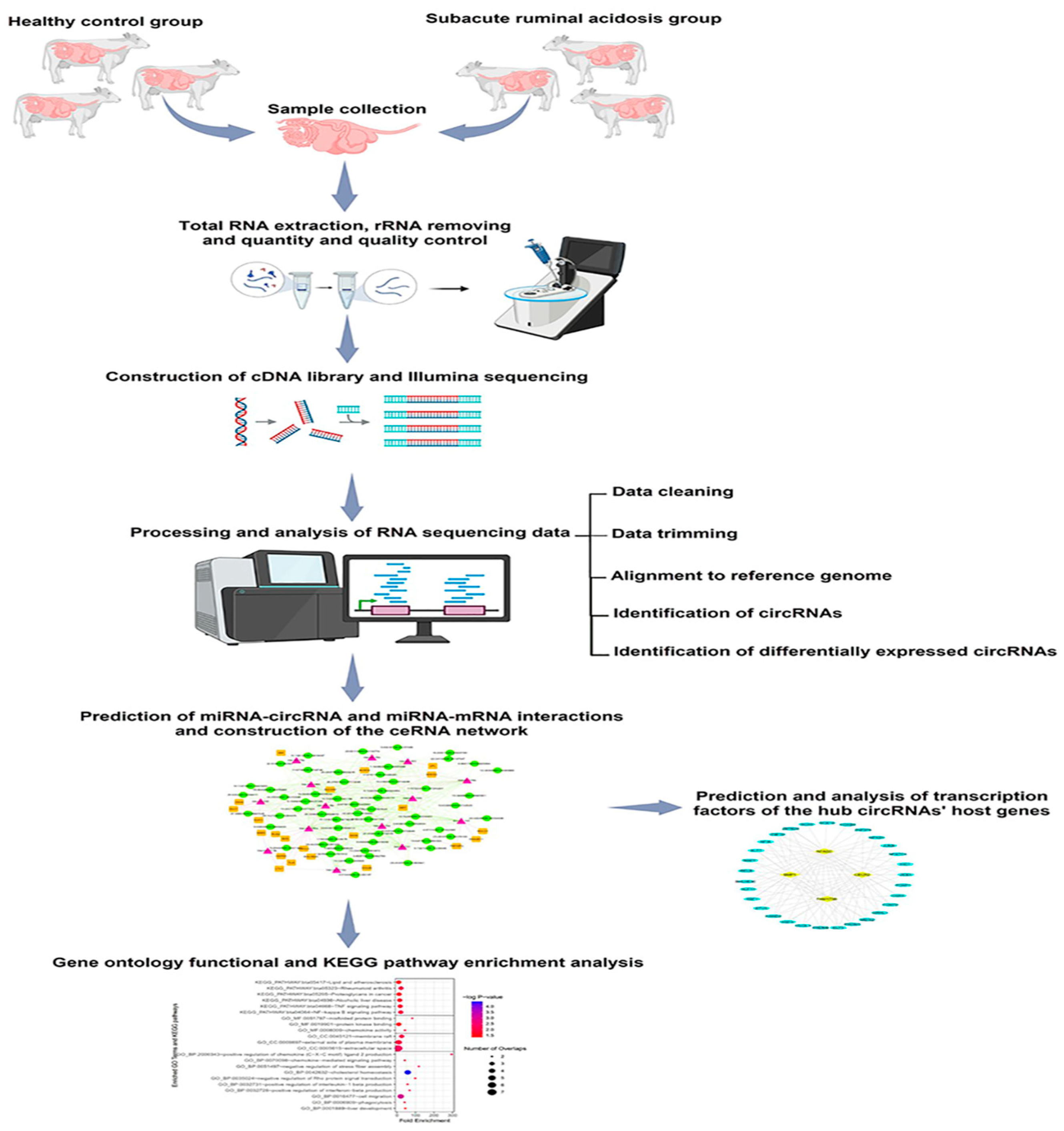
2.1. Overview of Study Workflow
2.2. Animals
2.3. Induction of Rumen Acidosis and Sample Collection
2.4. RNA Extraction
2.5. cDNA Library Construction and Sequencing
2.6. Data Preprocessing for Quality Enhancement in NGS Analysis
2.7. Alignment to Reference Genome
2.8. CircRNA Identification
2.9. Normalization and Identification of Differential Expression (DE) cicRNAs
2.10. Construction of ceRNA Network of circRNA–miRNA–mRNA Interactions
2.11. Transcription Factor Analysis for the Host Genes of Hub circRNAs
2.12. Functional and Pathway Enrichment Analyses
3. Results
3.1. Animal Health Monitoring
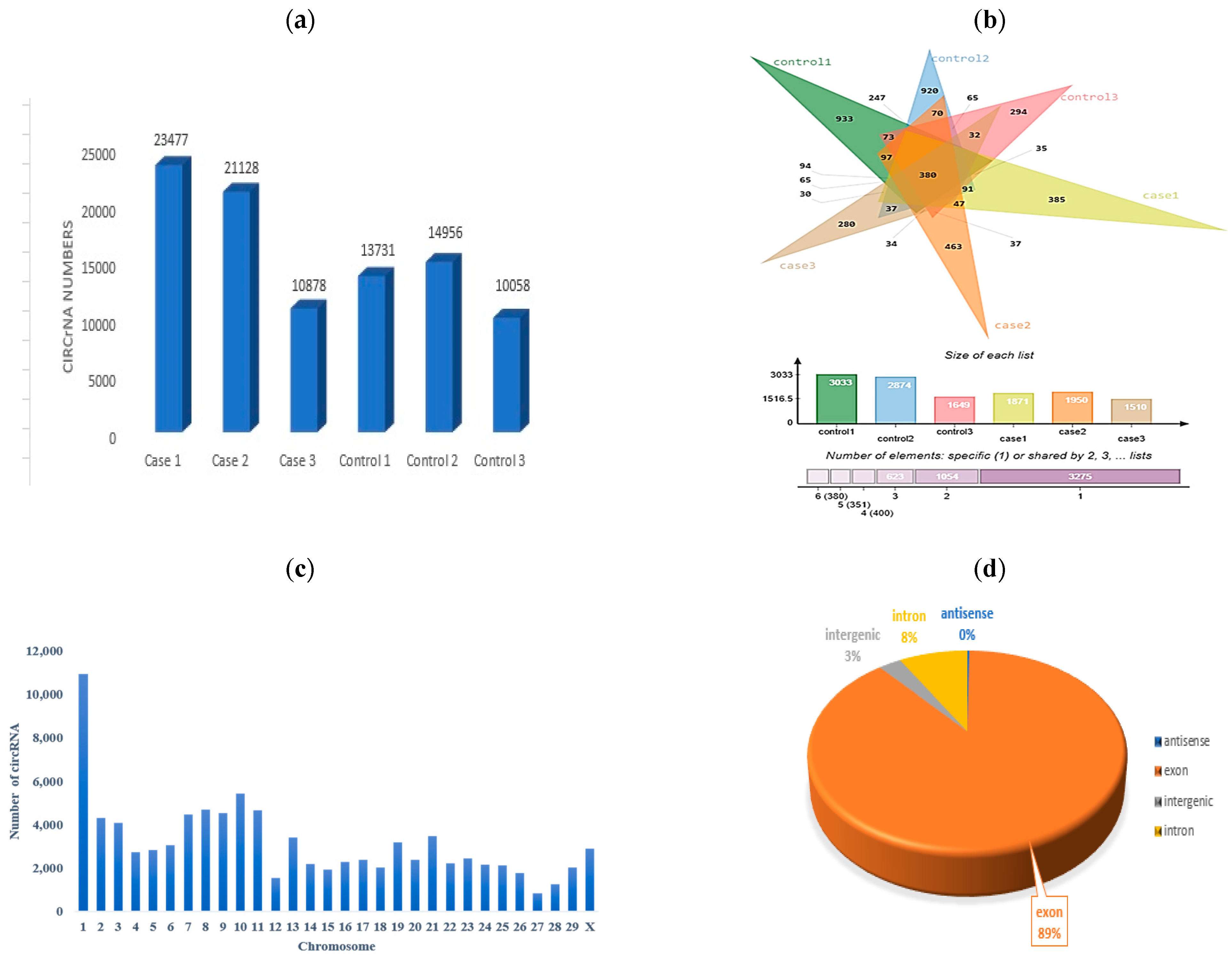
3.2. Rumen Metabolic Parameters and circRNA Quality Control
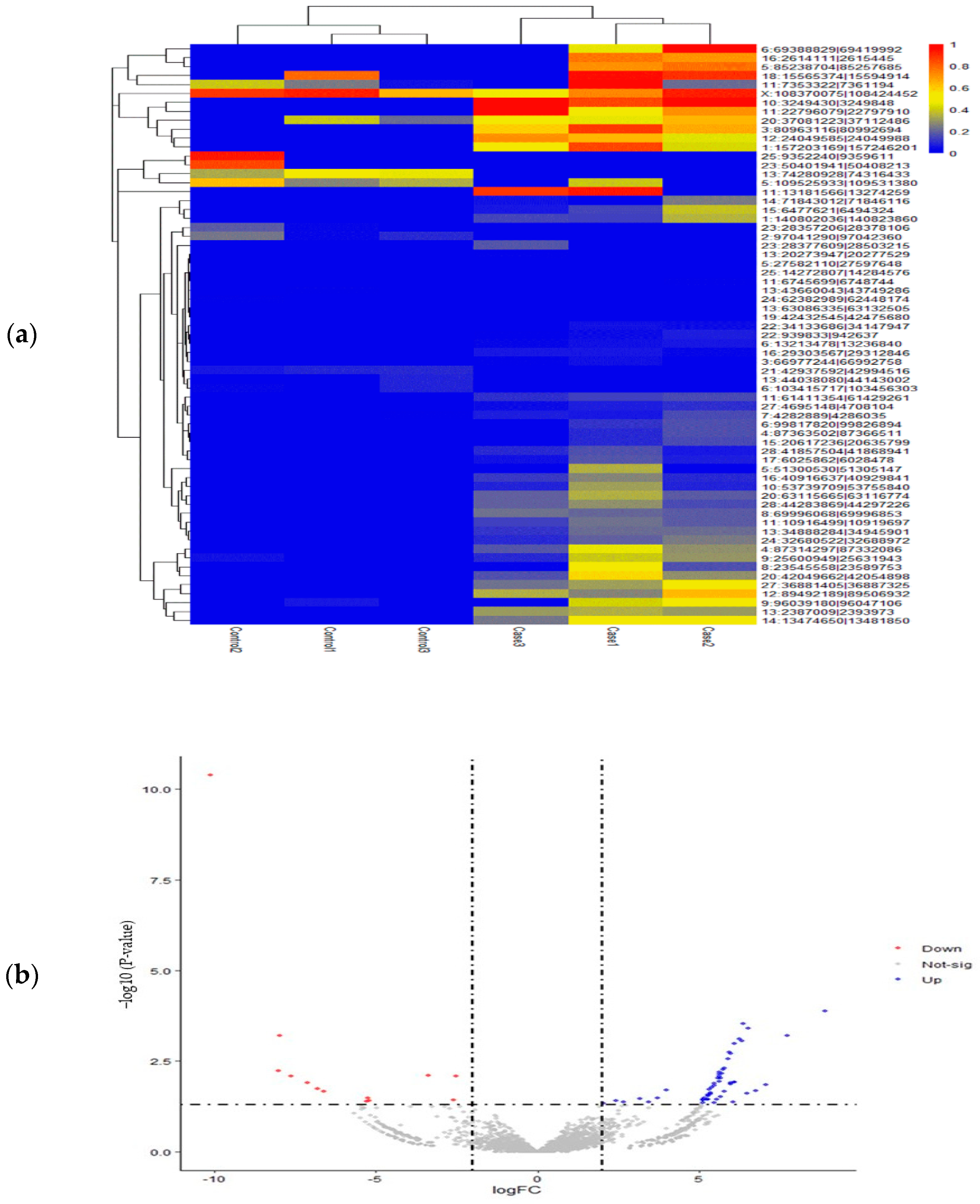
3.3. Identification of DE-circRNAs in SARA-Affected Cattle
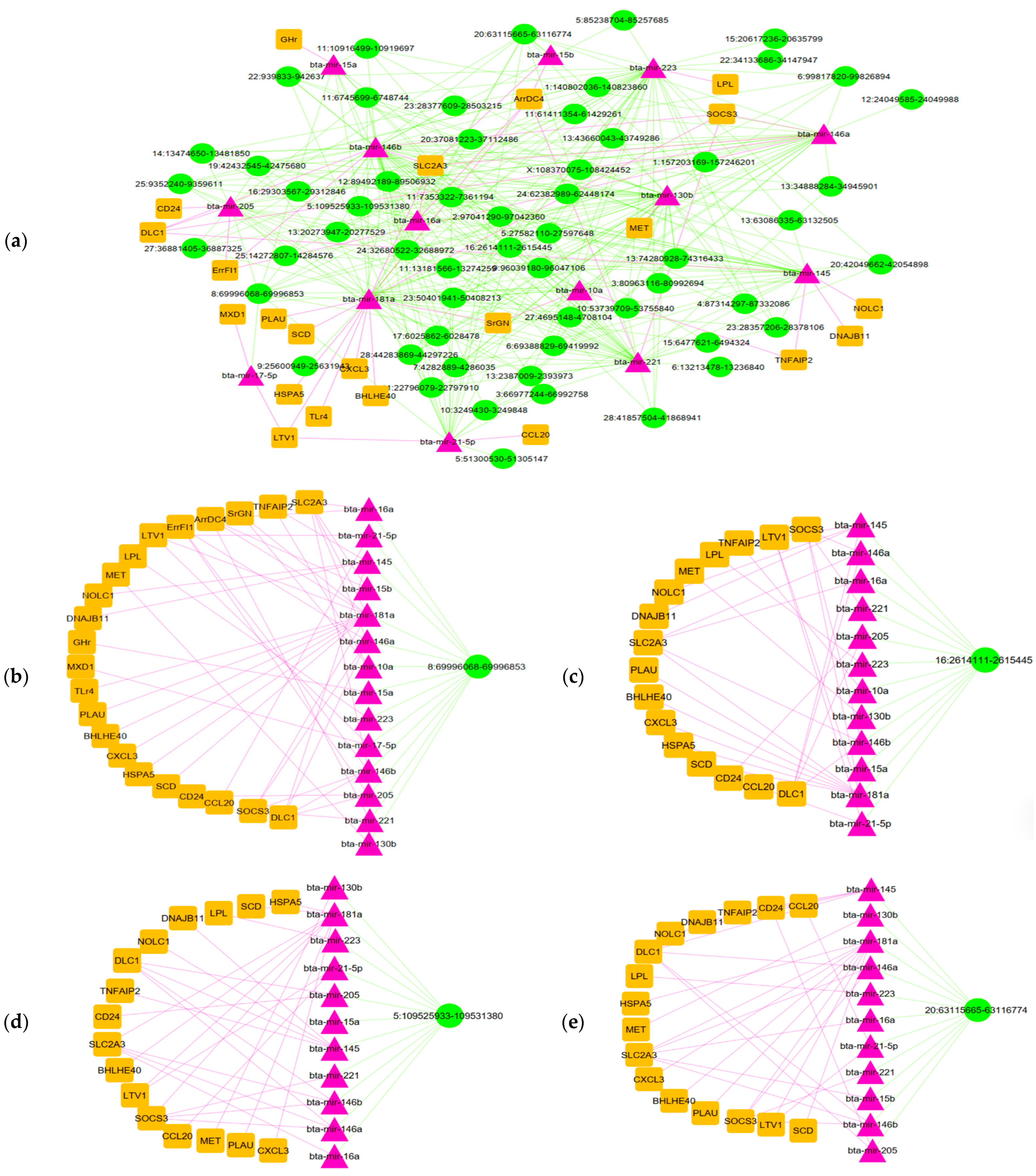
3.4. Analysis of circRNA–miRNA–mRNA Network
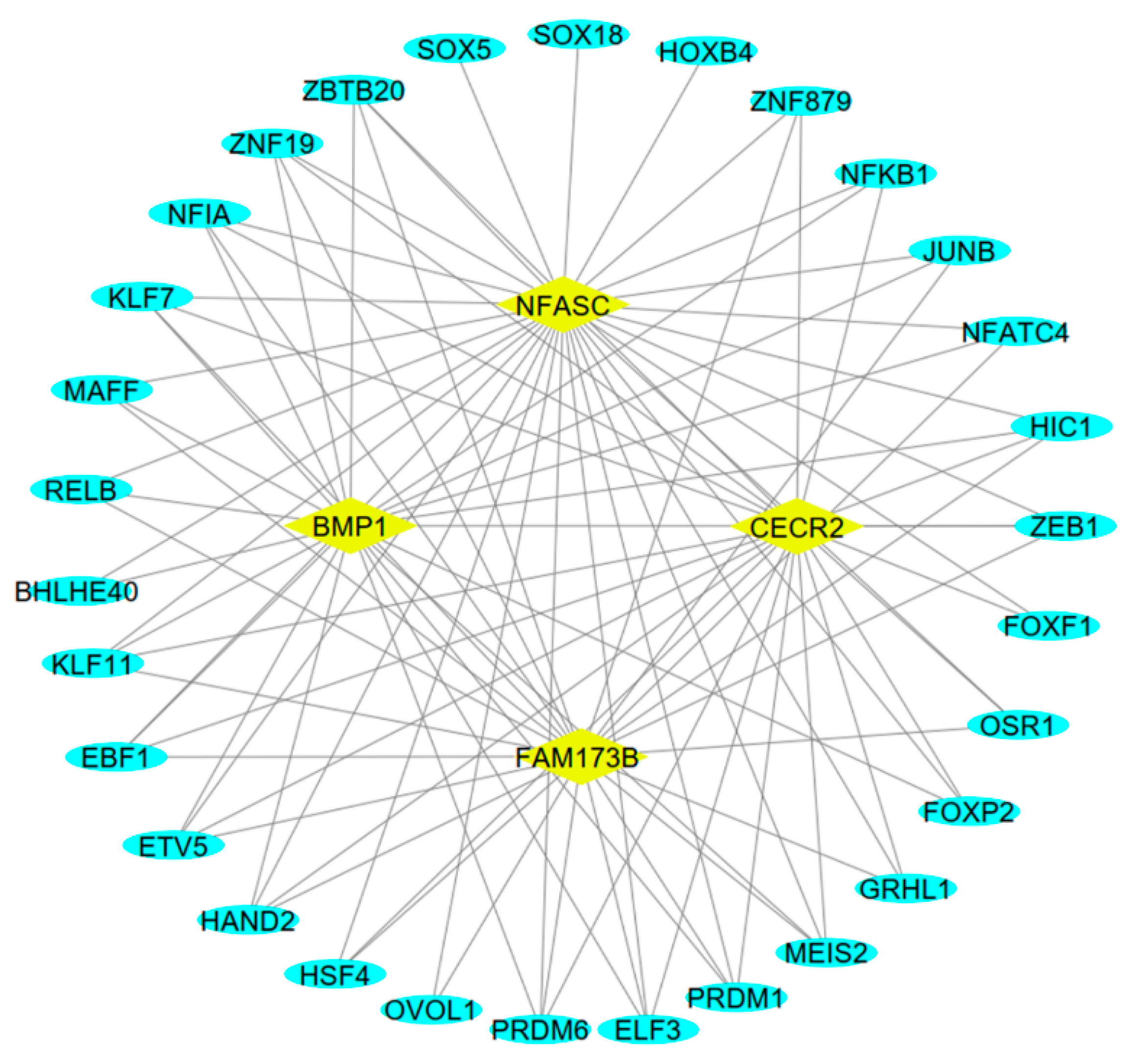
3.5. Analysis of TFs Associated with Host Genes of Hub circRNAs
3.6. Functional and Pathway Enrichment Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Shen, Y.; Wang, C.; Ding, L.; Zhao, F.; Wang, M.; Fu, J.; Wang, H. Megasphaera elsdenii lactate degradation pattern shifts in rumen acidosis models. Front. Microbiol. 2019, 10, 162. [Google Scholar] [CrossRef]
- Santos, F.A.P.; da Silva Marques, R.; Dórea, J.R.R. Grain processing for beef cattle. In Rumenology; Springer: Cham, Switzerland, 2016; pp. 213–241. [Google Scholar]
- Hernandez, J.; Benedito, J.L.; Abuelo, A.; Castillo, C. Ruminal acidosis in feedlot: From aetiology to prevention. Sci. World J. 2014, 2014, 702572. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, T.G.; Titgemeyer, E.C. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook. J. Dairy Sci. 2007, 90 (Suppl. S1), E17–E38. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo-López, E.; Itza-Ortiz, M.F.; Peraza-Mercado, G.; Carrera-Chávez, J.M. Ruminal acidosis: Strategies for its control. Austral. J. Vet. Sci. 2017, 49, 139–148. [Google Scholar] [CrossRef]
- Gozho, G.N.; Krause, D.O.; Plaizier, J.C. Ruminal lipopolysaccharide concentration and inflammatory response during grain-induced subacute ruminal acidosis in dairy cows. J. Dairy Sci. 2007, 90, 856–866. [Google Scholar] [CrossRef]
- Peek, S.F.; Divers, T.J. Rebhun’s Diseases of Dairy Cattle-E-Book; Elsevier Health Sciences: Maryland Heights, MO, USA, 2018; ISBN 978-0-323-39662-2. [Google Scholar]
- Giurgiu, O.V.; Berean, D.I.; Ionescu, A.; Ciupe, M.S.; Cimpean, C.R.; Radu, C.I.; Bitica, D.G.; Bogdan, S.; Bogdan, M.L. The effect of oral administration of zeolite on the energy metabolism and reproductive health of Romanian spotted breed in advanced gestation and post partum period. Vet. Anim. Sci. 2024, 23, 100333. [Google Scholar] [CrossRef]
- Ojo, O.E.; Hajek, L.; Johanns, S.; Pacífico, C.; Sener-Aydemir, A.; Ricci, S.; Rivera-Chacon, R.; Castillo-Lopez, E.; Reisinger, N.; Zebeli, Q. Evaluation of circulating microRNA profiles in blood as potential candidate biomarkers in a subacute ruminal acidosis cow model-a pilot study. BMC Genom. 2023, 24, 333. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Guan, L.L. Noncoding RNAs: Regulatory Molecules of Host-Microbiome Crosstalk. Trends Microbiol. 2021, 29, 713–724. [Google Scholar] [CrossRef]
- Zhou, W.Y.; Cai, Z.R.; Liu, J.; Wang, D.S.; Ju, H.Q.; Xu, R.H. Circular RNA: Metabolism, functions and interactions with proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle; National Academy of Sciences: Washington, DC, USA, 2001. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Guo, J.; Gao, J.; Liu, Z. HISAT2 Parallelization Method Based on Spark Cluster. J. Phys. Conf. Ser. 2022, 2179, 012038. [Google Scholar]
- Gao, Y.; Zhang, J.; Zhao, F. Circular RNA identification based on multiple seed matching. Brief. Bioinform. 2018, 19, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, S.; Yang, J.; Zhao, F. Accurate quantification of circular RNAs identifies extensive circular isoform switching events. Nat. Commun. 2020, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh, M.; Fayazi, J.; Asgari, Y.; Zali, H.; Kaderali, L. Reconstruction and Analysis of Cattle Metabolic Networks in Normal and Acidosis Rumen Tissue. Animals 2020, 10, 469. [Google Scholar] [CrossRef]
- Wang, J.; Chen, H.; Zhang, Y.; Jiang, S.; Zeng, X.; Shen, H. Comprehensive Analysis of Differentially Expressed CircRNAs in the Ovaries of Low- and High-Fertility Sheep. Animals 2023, 13, 236. [Google Scholar] [CrossRef]
- Wang, H.; Gao, X.; Yu, S.; Wang, W.; Liu, G.; Jiang, X.; Sun, D. Circular RNAs regulate parental gene expression: A new direction for molecular oncology research. Front. Oncol. 2022, 12, 947775. [Google Scholar] [CrossRef]
- Wang, J.; Shi, L.; Zhang, X.; Hu, R.; Yue, Z.; Zou, H.; Peng, Q.; Jiang, Y.; Wang, Z. Metabolomics and proteomics insights into subacute ruminal acidosis etiology and inhibition of proliferation of yak rumen epithelial cells in vitro. BMC Genom. 2024, 25, 394. [Google Scholar] [CrossRef]
- Lu, Z.; Yao, L.; Jiang, Z.; Aschenbach, J.R.; Martens, H.; Shen, Z. Acidic pH and short-chain fatty acids activate Na+ transport but differentially modulate expression of Na+/H+ exchanger isoforms 1, 2, and 3 in omasal epithelium. J. Dairy Sci. 2016, 99, 733–745. [Google Scholar] [CrossRef]
- Shen, Z.; Martens, H.; Schweigel-Rontgen, M. Na+ transport across rumen epithelium of hay-fed sheep is acutely stimulated by the peptide IGF-1 in vitro. Exp. Physiol. 2012, 97, 497–505. [Google Scholar] [CrossRef]
- Toker, A. Phosphoinositides and signal transduction. Cell. Mol. Life Sci. 2002, 59, 761–779. [Google Scholar] [CrossRef]
- Palhano, R.L.A.; Martins, R.A.; Lemos, G.S.; Faleiros, R.R.; da Fonseca, L.A.; Gorza, L.L.; Lopes, C.E.B.; Meneses, R.M.; de Carvalho, A.U.; Filho, E.J.F.; et al. Exploring the impact of high-energy diets on cattle: Insights into subacute rumen acidosis, insulin resistance, and hoof health. J. Dairy Sci. 2024, 107, 5054–5069. [Google Scholar] [CrossRef] [PubMed]
- Baldini, N.; Avnet, S. The Effects of Systemic and Local Acidosis on Insulin Resistance and Signaling. Int. J. Mol. Sci. 2018, 20, 126. [Google Scholar] [CrossRef]
- Gamba, M.; Pralong, F.P. Control of GnRH neuronal activity by metabolic factors: The role of leptin and insulin. Mol. Cell. Endocrinol. 2006, 254–255, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Hassanein, E.M.; Szelenyi, Z.; Szenci, O. Gonadotropin-Releasing Hormone (GnRH) and Its Agonists in Bovine Reproduction I: Structure, Biosynthesis, Physiological Effects, and Its Role in Estrous Synchronization. Animals 2024, 14, 1473. [Google Scholar] [CrossRef] [PubMed]
- Pacífico, C.; Ricci, S.; Sajovitz, F.; Castillo-Lopez, E.; Rivera-Chacon, R.; Petri, R.M.; Zebeli, Q.; Reisinger, N.; Kreuzer-Redmer, S. Bovine rumen epithelial miRNA-mRNA dynamics reveals post-transcriptional regulation of gene expression upon transition to high-grain feeding and phytogenic supplementation. Genomics 2022, 114, 110333. [Google Scholar] [CrossRef]
- Carow, B.; Rottenberg, M.E. SOCS3, a Major Regulator of Infection and Inflammation. Front. Immunol. 2014, 5, 58. [Google Scholar] [CrossRef]
- Xie, W.; Li, Z.; Li, M.; Xu, N.; Zhang, Y. miR-181a and inflammation: miRNA homeostasis response to inflammatory stimuli in vivo. Biochem. Biophys. Res. Commun. 2013, 430, 647–652. [Google Scholar] [CrossRef]
- Yao, X.; He, Z.; Qin, C.; Deng, X.; Bai, L.; Li, G.; Shi, J. SLC2A3 promotes macrophage infiltration by glycolysis reprogramming in gastric cancer. Cancer Cell Int. 2020, 20, 503. [Google Scholar] [CrossRef]
- Wang, J.; Bai, X.; Song, Q.; Fan, F.; Hu, Z.; Cheng, G.; Zhang, Y. miR-223 Inhibits Lipid Deposition and Inflammation by Suppressing Toll-Like Receptor 4 Signaling in Macrophages. Int. J. Mol. Sci. 2015, 16, 24965–24982. [Google Scholar] [CrossRef]
- Yang, Z.; Tang, Y.; Wu, X.; Wang, J.; Yao, W. MicroRNA-130b Suppresses Malignant Behaviours and Inhibits the Activation of the PI3K/Akt Signaling Pathway by Targeting MET in Pancreatic Cancer. Biochem. Genet. 2024, 63, 1660–1685. [Google Scholar] [CrossRef]
- He, X.; Li, Y.; Deng, B.; Lin, A.; Zhang, G.; Ma, M.; Wang, Y.; Yang, Y.; Kang, X. The PI3K/AKT signalling pathway in inflammation, cell death and glial scar formation after traumatic spinal cord injury: Mechanisms and therapeutic opportunities. Cell Prolif. 2022, 55, e13275. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.N.; Fan, L.; Sweet, D.R.; Jain, M.K. The Kruppel-Like Factors and Control of Energy Homeostasis. Endocr. Rev. 2019, 40, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Saito-Adachi, M.; Arai, Y.; Fujiwara, Y.; Takai, E.; Shibata, S.; Seki, M.; Rokutan, H.; Maeda, D.; Horie, M. E74-like factor 3 is a key regulator of epithelial integrity and immune response genes in biliary tract cancer. Cancer Res. 2021, 81, 489–500. [Google Scholar] [CrossRef]
- Fan, W.-J.; Li, H.-P.; Zhu, H.-S.; Sui, S.-P.; Chen, P.-G.; Deng, Y.; Sui, T.-M.; Wang, Y.-Y. NF-κB is involved in the LPS-mediated proliferation and apoptosis of MAC-T epithelial cells as part of the subacute ruminal acidosis response in cows. Biotechnol. Lett. 2016, 38, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- Somma, D.; Kok, F.O.; Kerrigan, D.; Wells, C.A.; Carmody, R.J. Defining the Role of Nuclear Factor (NF)-kappaB p105 Subunit in Human Macrophage by Transcriptomic Analysis of NFKB1 Knockout THP1 Cells. Front. Immunol. 2021, 12, 669906. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Silva, L.B.; dos Santos Neto, A.P.; Maia, S.M.; dos Santos Guimarães, C.; Quidute, I.L.; Carvalho, A.D.A.T.; Júnior, S.A.; Leão, J.C. The role of TNF-α as a proinflammatory cytokine in pathological processes. Open Dent. J. 2019, 13, 332–338. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, G.; Li, X.; Guan, Y.; Wang, Y.; Yuan, X.; Sun, G.; Wang, Z.; Li, X. Inflammatory mechanism of Rumenitis in dairy cows with subacute ruminal acidosis. BMC Vet. Res. 2018, 14, 135. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Control Diet (%) | High-Concentrate Diet (%) |
|---|---|---|
| Alfalfa hay | 10.5 | 10.5 |
| Corn silage | 14.5 | 14.5 |
| Barley grain | 34 | 34 |
| Corn grain ground | 5.3 | 23.3 |
| Soybean meal | 9 | 9 |
| Wheat bran | 10.5 | 3 |
| Rice bran | 6 | 2.5 |
| Sucrose | 5 | - |
| Fat powder | 2.5 | - |
| Supplement | 3.2 | 3.2 |
| Item | Case (Mean ± SEM) | Control (Mean ± SEM) | p-Value |
|---|---|---|---|
| pH_1 | 5.61 ± 0.21 | 5.83 ± 0.06 | 0.042 * |
| pH_30 | 5.56 ± 0.23 | 5.80 ± 0.12 | 0.038 * |
| pH_129 | 5.18 ± 0.04 | 5.81 ± 0.28 | 0.009 ** |
| Acetate (mM) | 69.70 ± 0.82 | 71.74 ± 0.78 | 0.098 ns |
| Propionate (mM) | 18.11 ± 0.49 | 14.45 ± 0.65 | 0.003 ** |
| Butyrate (mM) | 13.22 ± 0.17 | 14.21 ± 0.11 | 0.012 * |
| Valerate (mM) | 1.45 ± 0.07 | 1.25 ± 0.09 | 0.001 *** |
| NH₃-N (mg/dL) | 7.71 ± 0.21 | 6.96 ± 0.08 | 0.021 * |
| Sample | Case 1 | Case 2 | Case 3 | Control 1 | Control 2 | Control 3 |
|---|---|---|---|---|---|---|
| Count | 23,477 | 21,128 | 10,878 | 13,731 | 14,956 | 10,058 |
| Total (2×) | 46,954 | 42,256 | 21,756 | 27,462 | 29,912 | 20,116 |
| CircRNA Id | logFC | p-Value | Up/Down Change |
|---|---|---|---|
| 11:13181566|13274259 | 8.922037 | 0.000134 | up |
| 28:41857504|41868941 | 6.357273 | 0.000294 | up |
| 13:2387009|2393973 | 6.531094 | 0.000392 | up |
| 25:14272807|14284576 | 7.736988 | 0.000626 | up |
| 3:80963116|80992694 | 6.255852 | 0.000763 | up |
| 13:34888284|34945901 | 6.334839 | 0.000889 | up |
| 22:939833|942637 | 6.121243 | 0.001056 | up |
| 12:24049585|24049988 | 5.952984 | 0.001773 | up |
| 4:87314297|87332086 | 5.99522 | 0.002001 | up |
| 10:3249430|3249848 | 5.898283 | 0.002758 | up |
| 20:63115665|63116774 | 5.785773 | 0.004924 | up |
| 17:6025862|6028478 | 5.760129 | 0.005367 | up |
| 6:13213478|13236840 | 5.648043 | 0.006443 | up |
| 13:20273947|20277529 | 5.724471 | 0.006985 | up |
| 16:40916637|40929841 | 5.652667 | 0.007216 | up |
| 8:69996068|69996853 | 5.626879 | 0.008086 | up |
| 27:36881405|36887325 | 5.678958 | 0.009176 | up |
| 11:61411354|61429261 | 5.589611 | 0.009462 | up |
| 11:22796079|22797910 | 5.653773 | 0.011334 | up |
| 14:71843012|71846116 | 6.099256 | 0.012034 | up |
| 13:74280928|74316433 | −10.1139 | 4.14 × 10−11 | Down |
| 23:28357206|28378106 | −7.97577 | 0.000617 | Down |
| 24:62382989|62448174 | −8.02504 | 0.005943 | Down |
| 5:109525933|109531380 | −3.39204 | 0.00778 | Down |
| X:108370075|108424452 | −2.5376 | 0.008075 | Down |
| 25:9352240|9359611 | −7.61768 | 0.008212 | Down |
| 23:50401941|50408213 | −7.10743 | 0.0126 | Down |
| 13:44038080|44143002 | −6.81868 | 0.018544 | Down |
| 13:43660043|43749286 | −6.61989 | 0.021539 | Down |
| 21:42937592|42994516 | −5.2342 | 0.033789 | Down |
| 11:7353322|7361194 | −2.5856 | 0.037708 | Down |
| 2:97041290|97042360 | −5.21853 | 0.038482 | Down |
| 6:103415717|103456303 | −5.29858 | 0.0415 | Down |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neysi, S.; Fayazi, J.; Roshanfekr, H.; Imumorin, I.G. Preliminary Study of Differential circRNA Expression and Investigation of circRNA–miRNA–mRNA Competitive Endogenous Network in Rumen Acidosis of Holstein Cattle. Animals 2025, 15, 1472. https://doi.org/10.3390/ani15101472
Neysi S, Fayazi J, Roshanfekr H, Imumorin IG. Preliminary Study of Differential circRNA Expression and Investigation of circRNA–miRNA–mRNA Competitive Endogenous Network in Rumen Acidosis of Holstein Cattle. Animals. 2025; 15(10):1472. https://doi.org/10.3390/ani15101472
Chicago/Turabian StyleNeysi, Saeid, Jamal Fayazi, Hedayatollah Roshanfekr, and Ikhide G. Imumorin. 2025. "Preliminary Study of Differential circRNA Expression and Investigation of circRNA–miRNA–mRNA Competitive Endogenous Network in Rumen Acidosis of Holstein Cattle" Animals 15, no. 10: 1472. https://doi.org/10.3390/ani15101472
APA StyleNeysi, S., Fayazi, J., Roshanfekr, H., & Imumorin, I. G. (2025). Preliminary Study of Differential circRNA Expression and Investigation of circRNA–miRNA–mRNA Competitive Endogenous Network in Rumen Acidosis of Holstein Cattle. Animals, 15(10), 1472. https://doi.org/10.3390/ani15101472