
Changes in Muscle Quality and Gut Microbiota of Whiteleg Shrimp (Penaeus vannamei) Within a Live Supply Chain
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
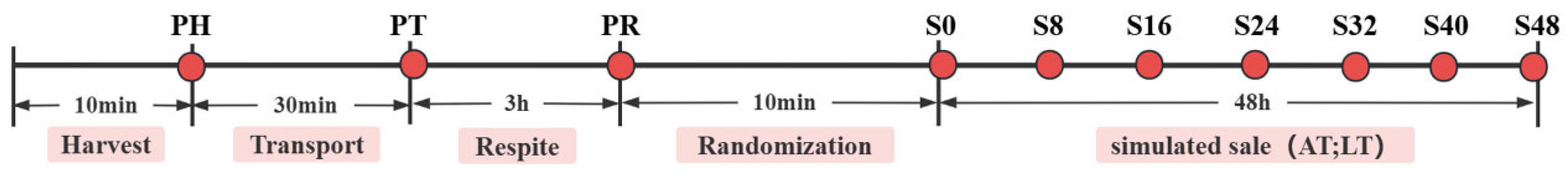
2.1. Experimental Design and Supply Chain Procedure
2.2. Sample Collection
2.3. Cumulative Survival Rate Analysis
2.4. Muscle Quality Analysis
2.4.1. Color Characteristic Analysis
2.4.2. Texture Profile Analysis of Fresh Muscle
2.4.3. TBARS Content in Muscle
2.4.4. pH and Lactate Content
2.5. Gut Microbiota
2.6. Data Evaluation and Analysis
3. Results
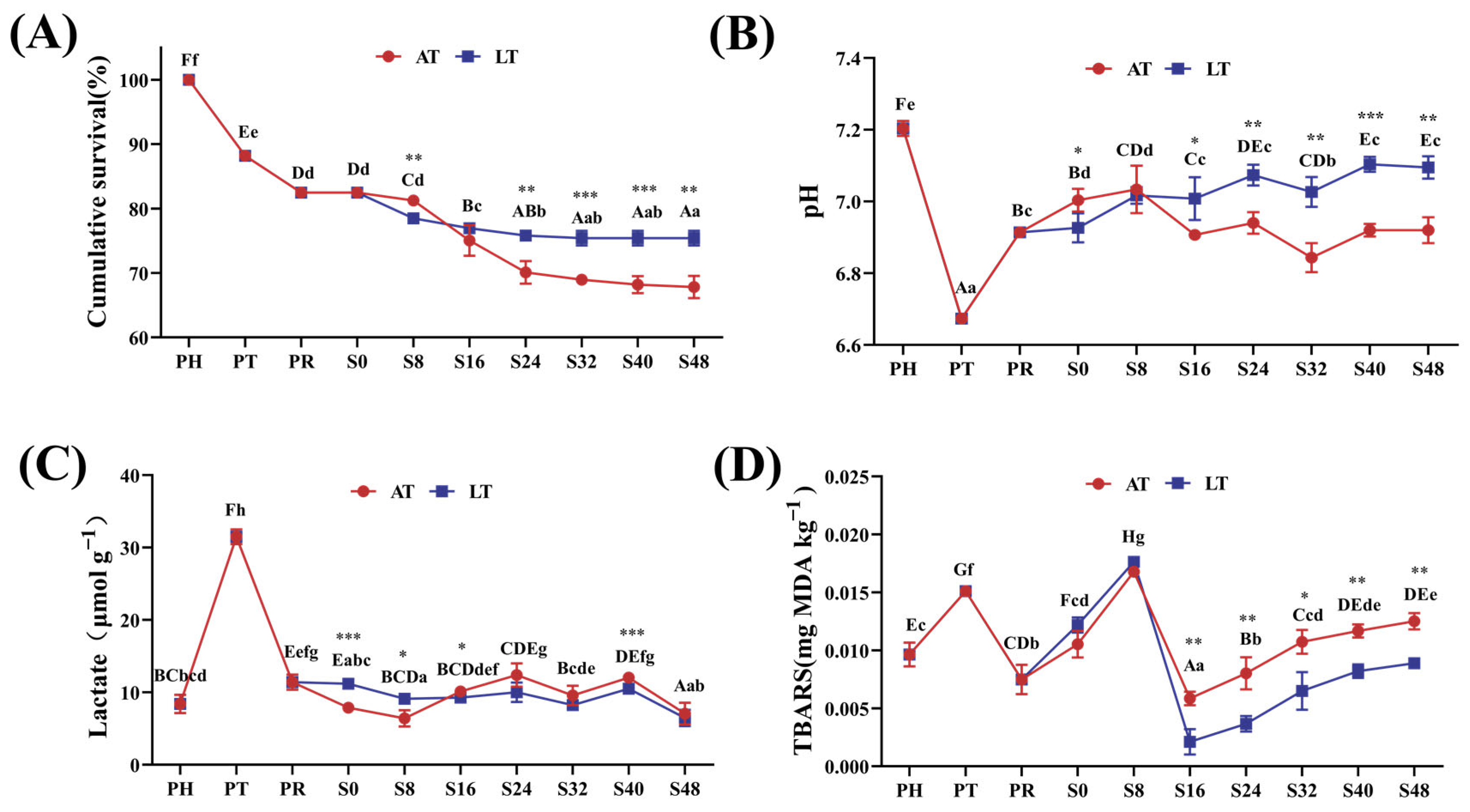
3.1. Cumulative Survival Rates
3.2. Changes in Muscle Quality
3.2.1. pH and Lactate Content
3.2.2. Muscle TBARS Content
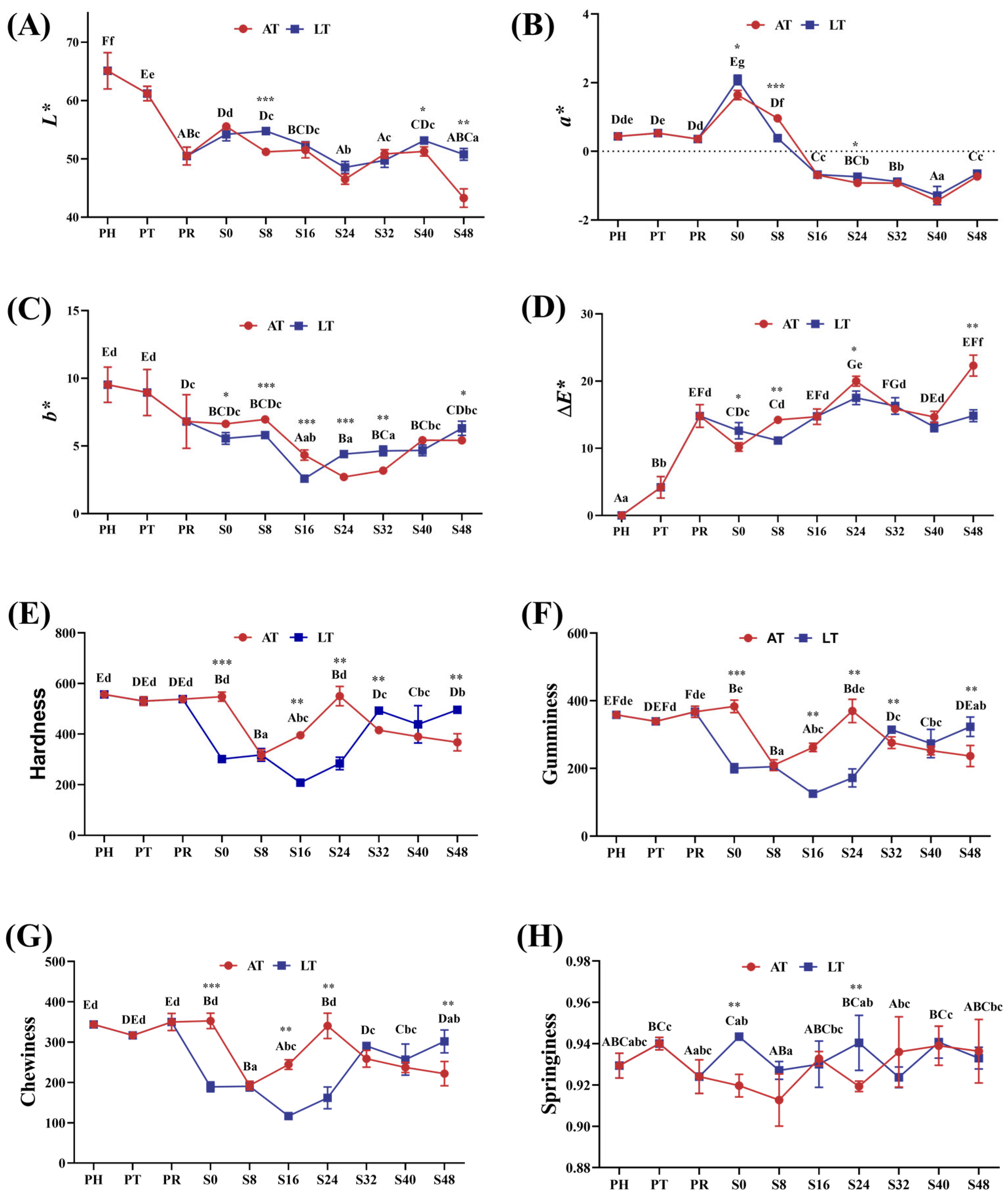
3.2.3. Color Characteristic Analysis
3.2.4. TPA
3.3. Changes in Gut Bacterial Communities of Shrimp
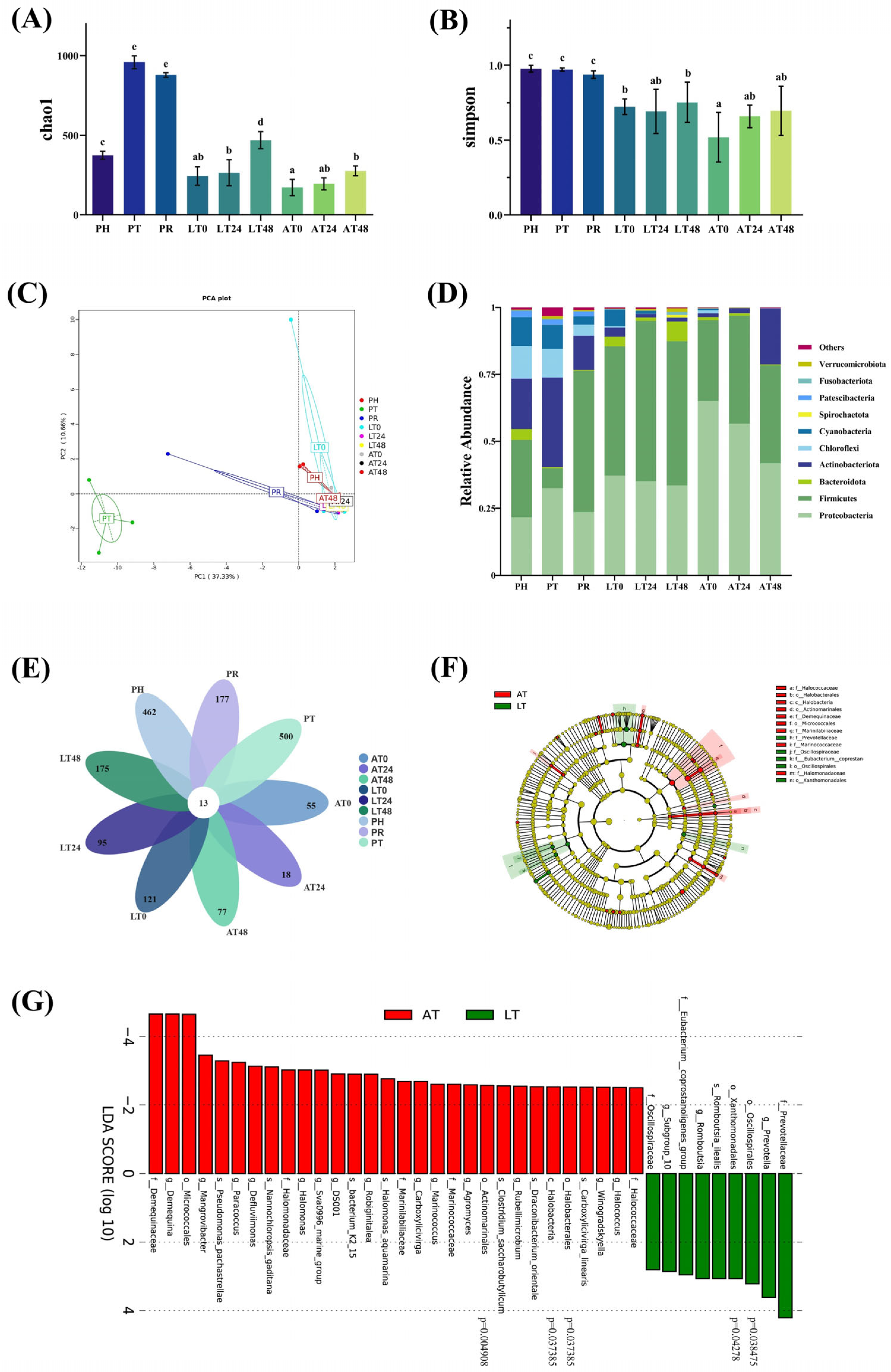
3.3.1. Diversity Analysis
3.3.2. Community Structure and Differential Analysis
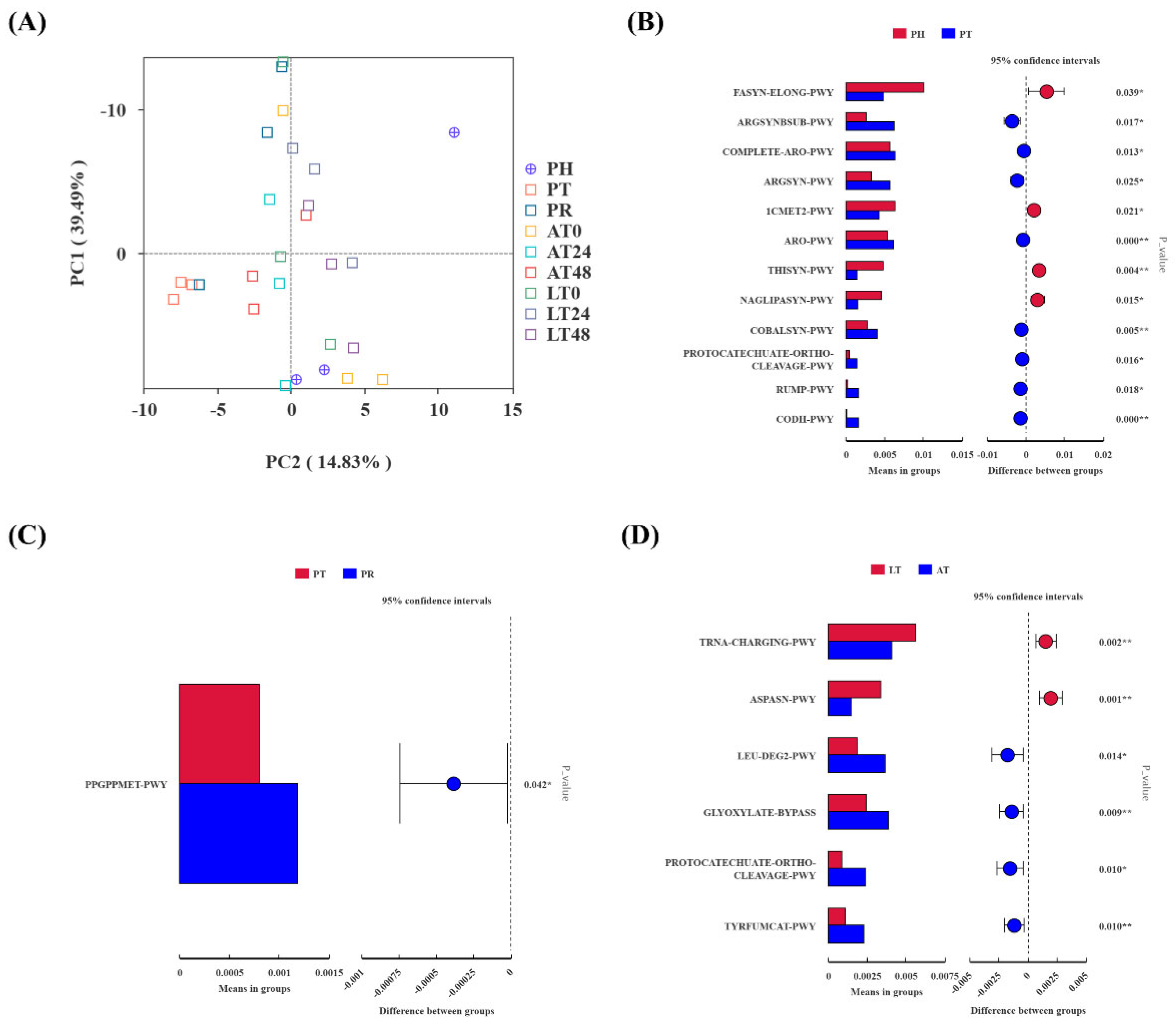
3.3.3. Predictive Analysis of PICRUSt2 Function Pathways in Gut Microbiota
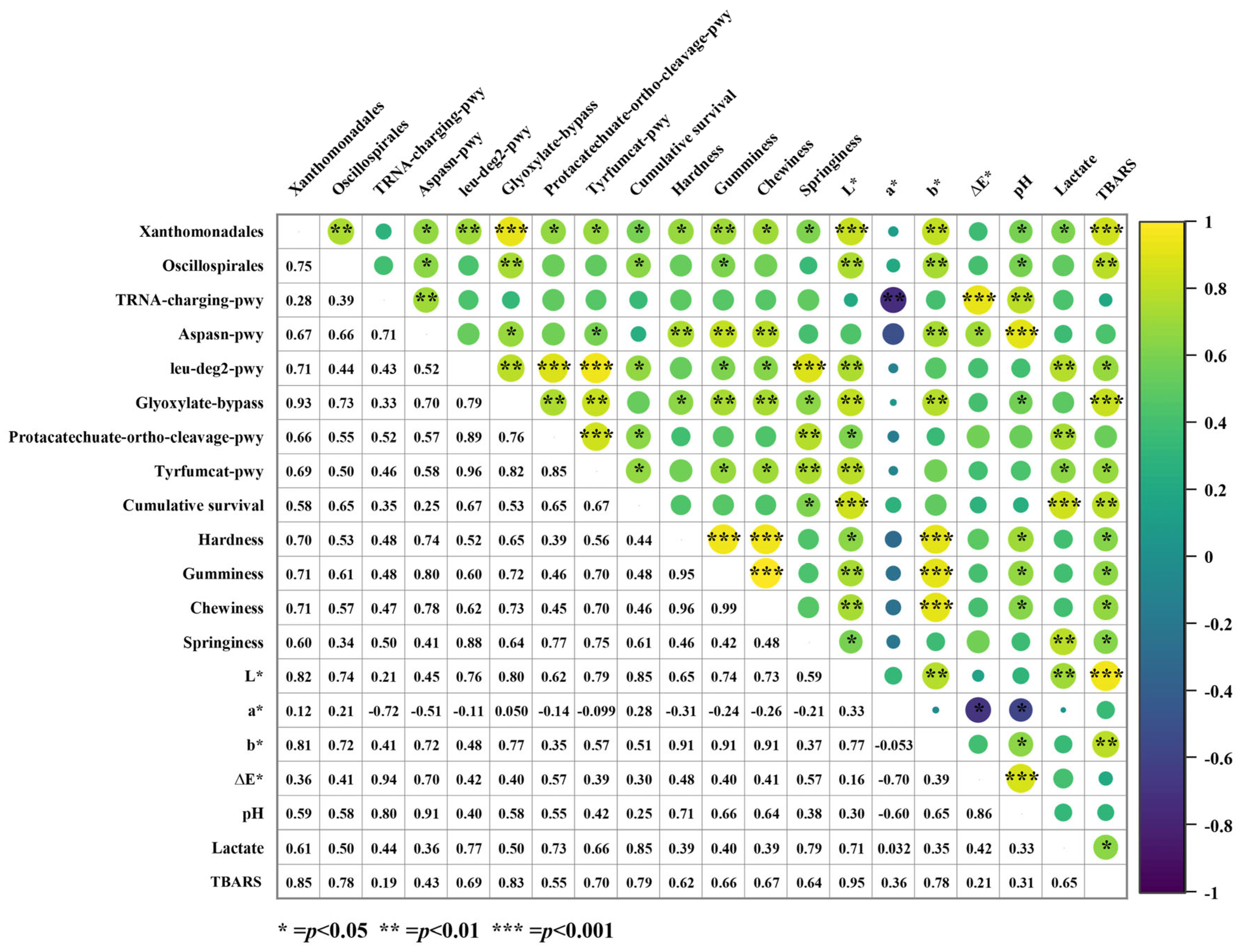
3.3.4. Correlation Analysis Between Differential Species, Differential Pathways, and Phenotypic Index in the LT Group
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sha, H.; Lu, J.; Chen, J.; Xiong, J. A meta-analysis study of the robustness and universality of gut microbiota-shrimp diseases relationship. Environ. Microbiol. 2022, 24, 3924–3938. [Google Scholar] [CrossRef]
- Emerenciano, M.G.C.; Rombenso, A.N.; Vieira, F.D.N.; Martins, M.A.; Coman, G.J.; Truong, H.H.; Noble, T.H.; Simon, C.J. Intensification of Penaeid Shrimp Culture: An Applied Review of Advances in Production Systems, Nutrition and Breeding. Animals 2022, 12, 236. [Google Scholar] [CrossRef]
- FAO. Global Aquaculture Production Quantity 1950–2020; FAO Fisheries and Aquaculture Division: Rome, Italy, 2024; Available online: https://www.fao.org/fishery/statistics-query/en/aquaculture/aquaculture_quantity (accessed on 28 March 2024).
- Wachirasiri, K.; Wanlapa, S.; Uttapap, D.; Puttanlek, C.; Rungsardthong, V. Effects of Multiple Freeze-Thaw Cycles on Biochemical and Physical Quality Changes of White Shrimp (Penaeus vannamei) Treated with Lysine and Sodium Bicarbonate. J. Food Sci. 2019, 84, 1784–1790. [Google Scholar] [CrossRef]
- Shi, J.; Lei, Y.; Shen, H.; Hong, H.; Yu, X.; Zhu, B.; Luo, Y. Effect of glazing and rosemary (Rosmarinus officinalis) extract on preservation of mud shrimp (Solenocera melantho) during frozen storage. Food Chem. 2019, 272, 604–612. [Google Scholar] [CrossRef]
- Sharifimehr, S.; Soltanizadeh, N.; Hossein Goli, S.A. Effects of edible coating containing nano-emulsion of Aloe vera and eugenol on the physicochemical properties of shrimp during cold storage. J. Sci. Food Agric. 2019, 99, 3604–3615. [Google Scholar] [CrossRef]
- DÍAz-Tenorio, L.M.; GarcÍA-CarreÑO, F.L.; Pacheco-Aguilar, R. Comparison of Freezing and Thawing Treatments on Muscle Properties of Whiteleg Shrimp (Litopenaeus vannamei). J. Food Biochem. 2007, 31, 563–576. [Google Scholar] [CrossRef]
- Hossain, A.; Nielsen, M.; Nielsen, R. The choice of prawn or shrimp? Lesson from consumer preferences and price acceptance in the Bangladesh wet fish market. Aquaculture 2024, 583, 740580. [Google Scholar] [CrossRef]
- Guan, W.; Nong, W.; Wei, X.; Chen, R.; Huang, Z.; Ding, Y.; Qin, X.; Cai, L.; Mao, L. Influences of two transport strategies on AMPK-mediated metabolism and flesh quality of shrimp (Litopenaeus vannamei). J. Sci. Food Agric. 2024, 104, 727–736. [Google Scholar] [CrossRef]
- Huang, Z.; Guan, W.; Lyu, X.; Chen, R.; Wu, Y.; Zheng, G.; Mao, L. Impacts of long-time transportation on whiteleg shrimp (Penaeus vannamei) muscle quality and underlying biochemical mechanisms. J. Sci. Food Agric. 2023, 103, 7590–7599. [Google Scholar] [CrossRef]
- Veiseth-Kent, E.; Grove, H.; Færgestad, E.M.; Fjæra, S.O. Changes in muscle and blood plasma proteomes of Atlantic salmon (Salmo salar) induced by crowding. Aquaculture 2010, 309, 272–279. [Google Scholar] [CrossRef]
- Lefèvre, F.; Bugeon, J.; Aupérin, B.; Aubin, J. Rearing oxygen level and slaughter stress effects on rainbow trout flesh quality. Aquaculture 2008, 284, 81–89. [Google Scholar] [CrossRef]
- Peng, L.; You, J.; Wang, L.; Xiong, S.; Huang, Q.; Yin, T. Effect of Respite Time before Live Transportation on Muscle Quality of Blunt Snout (Wuchang) Bream. Foods 2022, 11, 2254–2265. [Google Scholar] [CrossRef]
- Ha, H.J.; Min, J.; Lee, J.D.; Goo, J.S.; Kim, J.G.; Kim, T.J.; Patk, T.I. Optimal Conditions for Long-distance Transportation of Live Black Rockfish (Sebastes schlegeli) and Changes in their Characteristics during Transport. J. Aquat. Food Prod. Technol. 2019, 28, 762–771. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Z.; Lin, G.; Li, M.; Zhang, Y.; Mai, K. Intestinal microbiota differences associated with physiological health, meat quality, and regulating mineral interactions of Penaeus vannamei fed organic manganese. Aquaculture 2024, 582, e740537. [Google Scholar] [CrossRef]
- Lei, J.; Dong, Y.; Hou, Q.; He, Y.; Lai, Y.; Liao, C.; Kawamura, Y.; Li, J.; Zhang, B. Intestinal Microbiota Regulate Certain Meat Quality Parameters in Chicken. Front. Nutr. 2022, 9, 747705. [Google Scholar] [CrossRef]
- Liu, B.; Cui, Y.; Ali, Q.; Zhu, X.; Li, D.; Ma, S.; Wang, Z.; Wang, C.; Shi, Y. Gut Microbiota Modulate Rabbit Meat Quality in Response to Dietary Fiber. Front. Nutr. 2022, 9, 849429. [Google Scholar] [CrossRef]
- Chen, C.; Qu, M.; Li, G.; Wan, G.; Liu, P.; Omar, S.M.; Mei, W.; Hu, Z.; Zhou, Q.; Xu, L. Dietary Tributyrin Improves Growth Performance, Meat Quality, Muscle Oxidative Status, and Gut Microbiota in Taihe Silky Fowls under Cyclic Heat Stress. Animals 2024, 14, 3041. [Google Scholar] [CrossRef]
- Holmes, E.; Li, J.V.; Marchesi, J.R.; Nicholson, J.K. Gut microbiota composition and activity in relation to host metabolic phenotype and disease risk. Cell Metab. 2012, 16, 559–564. [Google Scholar] [CrossRef]
- Hou, D.; Zhou, R.; Zeng, S.; Wei, D.; Deng, X.; Xing, C.; Yu, L.; Deng, Z.; Wang, H.; Weng, S.; et al. Intestine Bacterial Community Composition of Shrimp Varies Under Low- and High-Salinity Culture Conditions. Front. Microbiol. 2020, 11, e589164. [Google Scholar] [CrossRef]
- Zhou, L.; Li, H.; Qin, J.G.; Wang, X.; Chen, L.; Xu, C.; Li, E. Dietary prebiotic inulin benefits on growth performance, antioxidant capacity, immune response and intestinal microbiota in Pacific white shrimp (Litopenaeus vannamei) at low salinity. Aquaculture 2020, 518, e734847. [Google Scholar] [CrossRef]
- Al-Masqari, Z.A.; Guo, H.; Wang, R.; Yan, H.; Dong, P.; Wang, G.; Zhang, D. Effects of high temperature on water quality, growth performance, enzyme activity and the gut bacterial community of shrimp (Litopenaeus vannamei). Aquac. Res. 2022, 53, 3283–3296. [Google Scholar] [CrossRef]
- Wongpracha, P.; Kanthawong, P.; Arun, J.; Jeyaraj, G.P.; Jongput, B.; Srijad, S.; Kitikiew, S. Effects of Bacillus probiotics, Bacillus subtilis and Bacillus cereus dietary additional to controlling Vibriosis infection of white shrimp (Litopenaeus vannamei). Int. J. Agric. Technol. 2024, 20, 427–440. [Google Scholar]
- T/SZS 3016-2020; Shenzhen Quality Food—Code of Good Practice for Fresh Aquatic Products. Shenzhen Standard Promotion Association: Shenzhen, China, 2020.
- Yuan, Q.; Qian, J.; Ren, Y.; Zhang, T.; Li, Z.; Liu, J. Effects of stocking density and water temperature on survival and growth of the juvenile Chinese mitten crab, Eriocheir sinensis, reared under laboratory conditions. Aquaculture 2018, 495, 631–636. [Google Scholar] [CrossRef]
- Zhu, Q.; Fu, S.; Yang, H.; Lin, B.; Lou, Y.; Li, Y. Effects of static magnetic field-assisted freezing on physicochemical indices and bacterial communities of shrimp (Penaeus vannamei). Food Biosci. 2023, 56, e103412. [Google Scholar] [CrossRef]
- Tan, M.; Lin, Z.; Zu, Y.; Zhu, B.; Cheng, S. Effect of multiple freeze-thaw cycles on the quality of instant sea cucumber: Emphatically on water status of by LF-NMR and MRI. Food Res. Int. 2018, 109, 65–71. [Google Scholar] [CrossRef]
- GB 5009.181-2016; Determination of Malondialdehyde in Foods. National Health Commission of the People’s Republic of China: Beijing, China, 2016.
- Chang, G.; Liu, Y.; Luo, Z.; Ni, K.; Zhang, P.; Zhou, T.; Bai, L.; Zhang, C.; Wang, X. Response surface methodology to optimize the sterilization process of slightly acidic electrolyzed water for Chinese shrimp (Fenneropenaeus chinensis) and to investigate its effect on shrimp quality. Food Chem. X 2024, 21, e101180. [Google Scholar] [CrossRef]
- Zeng, S.; Huang, Z.; Hou, D.; Liu, J.; Weng, S.; He, J. Composition, diversity and function of intestinal microbiota in pacific white shrimp (Litopenaeus vannamei) at different culture stages. PeerJ 2017, 5, e3986. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, C.; Xie, J.; Xu, C.; Zhao, Q.; Qin, J.G.; Chen, L.; Li, E. Intestinal bacterial signatures of the “cotton shrimp-like” disease explain the change of growth performance and immune responses in Pacific white shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 2019, 92, 629–636. [Google Scholar] [CrossRef]
- Wang, W.-z.; Huang, J.-s.; Zhang, J.-d.; Wang, Z.-l.; Li, H.-j.; Amenyogbe, E.; Chen, G. Effects of hypoxia stress on the intestinal microflora of juvenile of cobia (Rachycentron canadum). Aquaculture 2021, 536, 736419. [Google Scholar] [CrossRef]
- Cui, L.; Xie, Y.; Luo, K.; Wang, M.; Liu, L.; Li, C.; Tian, X. Physiological and intestinal microbiota responses of sea cucumber Apostichopus japonicus to various stress and signatures of intestinal microbiota dysbiosis. Front. Microbiol. 2024, 15, 1528275. [Google Scholar] [CrossRef]
- Xu, D.; Wu, J.; Sun, L.; Qin, X.; Fan, X.; Zheng, X. Combined stress of acute cold exposure and waterless duration at low temperature induces mortality of shrimp Litopenaeus vannamei through injuring antioxidative and immunological response in hepatopancreas tissue. J. Therm. Biol. 2021, 100, e103080. [Google Scholar] [CrossRef]
- Coyle, S.D.; Tidwell, J.H.; Yasharian, D.K.; Caporelli, A.; Skudlarek, N.A. The Effect of Biomass Density, Temperature, and Substrate on Transport Survival of Market-Size Freshwater Prawn, Macrobrachium rosenbergii. J. Appl. Aquac. 2005, 17, 61–71. [Google Scholar] [CrossRef]
- Brooks, G.A. Lactate as a fulcrum of metabolism. Redox Biol. 2020, 35, e101454. [Google Scholar] [CrossRef]
- Xu, D.; Sun, L.; Qin, X. Waterless live transport degrades the flesh quality of Litopenaeus vannamei by disturbing neuroendocrine response: Based on physiology and metabolomics. J. Sci. Food Agric. 2022, 103, 3882–3895. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Moreno, D.; Valenzuela-Soto, E.; Peregrino-Uriarte, A.; Leyva Carrillo, L.; Soñanez-Organis, J.; Yepiz-Plascencia, G. The pyruvate kinase of the whiteleg shrimp Litopenaeus vannamei: Gene structure and responses to short term hypoxia. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2023, 283, e111468. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, M.; Wang, Y.; Wang, Y.; Wei, X.; Wei, X.; Cai, W.-F.; Cai, W.-F.; Wu, J.; Wu, J.; et al. AMPK targets PDZD8 to trigger carbon source shift from glucose to glutamine. Cell Res. 2024, 34, 683–706. [Google Scholar] [CrossRef]
- Li, V.L.; He, Y.; Contrepois, K.; Liu, H.; Kim, J.T.; Wiggenhorn, A.L.; Tanzo, J.T.; Tung, A.S.-H.; Lyu, X.; Zushin, P.-J.H.; et al. An exercise-inducible metabolite that suppresses feeding and obesity. Nature 2022, 606, 785–790. [Google Scholar] [CrossRef]
- Yu, D.; Jiang, Q.; Xu, Y.; Xia, W. The shelf life extension of refrigerated grass carp (Ctenopharyngodon idellus) fillets by chitosan coating combined with glycerol monolaurate. Int. J. Biol. Macromol. 2017, 101, 448–454. [Google Scholar] [CrossRef]
- Liang, C.; Zhao, X.; Jiao, L.; Shen, Y.; Luo, J.; Zhu, T.; Zhao, W.; Gen, Z.; Zhou, Q.; Jin, M. Effects of different lipid sources on growth performance, fatty acids composition in tissue and expression of genes related to lipid metabolism in largemouth bass (Micropterus salmoides). Aquac. Rep. 2022, 23, 101013. [Google Scholar] [CrossRef]
- Haque, P.S.; Kapur, N.; Barrett, T.A.; Theiss, A.L. Mitochondrial function and gastrointestinal diseases. Nat. Rev. Gastroenterol. Hepatol. 2024, 21, 537–555. [Google Scholar] [CrossRef]
- Zerai, B.; Fitzsimmons, K.; Collier, R. Transcriptional Response of Delta-9-Desaturase Gene to Acute and Chronic Cold Stress in Nile Tilapia, Oreochromis niloticus. J. World Aquac. Soc. 2010, 41, 800–806. [Google Scholar] [CrossRef]
- Sun, Z.; Tan, X.; Liu, Q.; Ye, H.; Zou, C.; Xu, M.; Zhang, Y.; Ye, C. Physiological, immune responses and liver lipid metabolism of orange-spotted grouper (Epinephelus coioides) under cold stress. Aquaculture 2019, 498, 545–555. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; He, Y.; Ao, C.; Jiang, Y.; Tian, Y.; Zhao, H.; Lu, H. Effect of three unsaturated fatty acids on the protein oxidation and structure of myofibrillar proteins from rainbow trout (Oncorhynchus mykiss). Food Chem. 2024, 451, 139403. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Chen, J.; Zheng, Y.; Huang, J.; Xie, K.; Li, Y. Effect of atmospheric cold plasma treatment modes on the quality of red shrimp (Solenocera crassicornis) during cold chain storage. LWT 2023, 190, 115543. [Google Scholar] [CrossRef]
- Yeh, M.-S.; Lai, C.-Y.; Liu, C.-H.; Kuo, C.-M.; Cheng, W. A second proPO present in white shrimp Litopenaeus vannamei and expression of the proPOs during a Vibrio alginolyticus injection, molt stage, and oral sodium alginate ingestion☆. Fish Shellfish Immunol. 2009, 26, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Aaslyng, M.D.; Bejerholm, C.; Ertbjerg, P.; Bertram, H.C.; Andersen, H.J. Cooking loss and juiciness of pork in relation to raw meat quality and cooking procedure. Food Qual. Prefer. 2003, 14, 277–288. [Google Scholar] [CrossRef]
- Bordin, D.; Freire, C.A. Remarkable variability in stress responses among subtropical coastal marine teleosts. Mar. Biol. 2021, 168, 122–139. [Google Scholar] [CrossRef]
- Peng, Y.; Chen, S.; Ji, H.; Liu, S. Localization of trypsin-like protease in postmortem tissue of white shrimp (Litopenaeus vannamei) and its effect in muscle softening. Food Chem. 2019, 290, 277–285. [Google Scholar] [CrossRef]
- Sato, D.X.; Matsuda, Y.; Usio, N.; Funayama, R.; Nakayama, K.; Makino, T. Genomic adaptive potential to cold environments in the invasive red swamp crayfish. iScience 2023, 26, e107267. [Google Scholar] [CrossRef]
- Rungrassamee, W.; Klanchui, A.; Maibunkaew, S.; Chaiyapechara, S.; Jiravanichpaisal, P.; Karoonuthaisiri, N. Characterization of intestinal bacteria in wild and domesticated adult black tiger shrimp (Penaeus monodon). PLoS ONE 2014, 9, e91853. [Google Scholar] [CrossRef]
- Cheng, Y.; Ge, C.; Li, W.; Yao, H. The Intestinal Bacterial Community and Functional Potential of Litopenaeus vannamei in the Coastal Areas of China. Microorganisms 2021, 9, 1793–1809. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J. Progress in the gut microbiota in exploring shrimp disease pathogenesis and incidence. Appl. Microbiol. Biotechnol. 2018, 102, 7343–7350. [Google Scholar] [CrossRef]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A Common Factor in Human Diseases. Biomed. Res. Int. 2017, 2017, e9351507. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, X. Core Gut Microbiota of Shrimp Function as a Regulator to Maintain Immune Homeostasis in Response to WSSV Infection. Microbiol. Spectr. 2022, 10, e0246521. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E.; Harikrishnan, R.; Soltani, M.; Ghosh, K. The Effect of Gut Microbiota and Probiotics on Metabolism in Fish and Shrimp. Animals 2022, 12, 3016. [Google Scholar] [CrossRef]
- Xiao, X.; Zhou, Y.; Li, X.; Jin, J.; Durham, J.; Ye, Z.; Wang, Y.; Hennig, B.; Deng, P. 13C-Stable isotope resolved metabolomics uncovers dynamic biochemical landscape of gut microbiome-host organ communications in mice. Microbiome 2024, 12, 90. [Google Scholar] [CrossRef]
- Brown, E.M.; Ke, X.; Hitchcock, D.; Jeanfavre, S.; Avila-Pacheco, J.; Nakata, T.; Arthur, T.D.; Fornelos, N.; Heim, C.; Franzosa, E.A.; et al. Bacteroides-derived sphingolipids are critical for maintaining intestinal homeostasis and symbiosis. Cell Host Microbe 2019, 25, 668–680. [Google Scholar] [CrossRef]
- Lu, J.; Mao, J.; Qi, X.; Chen, J.; Xiong, J. The assembly of gut microbiota implicates shrimp acute hepatopancreas necrosis disease progression. Appl. Microbiol. Biotechnol. 2023, 107, 7489–7500. [Google Scholar] [CrossRef]
- Sun, F.; Wang, C.; Chen, L.; Weng, G.; Zheng, Z. The intestinal bacterial community of healthy and diseased animals and its association with the aquaculture environment. Appl. Microbiol. Biotechnol. 2020, 104, 775–783. [Google Scholar] [CrossRef]
- Dai, W.-F.; Zhang, J.-J.; Qiu, Q.-F.; Chen, J.; Yang, W.; Ni, S.; Xiong, J.-B. Starvation stress affects the interplay among shrimp gut microbiota, digestion and immune activities. Fish Shellfish Immunol. 2018, 80, 191–199. [Google Scholar] [CrossRef]
- Schicketanz, M.; Petrova, M.; Rejman, D.; Sosio, M.; Donadio, S.; Zhang, Y.E. Direct detection of stringent alarmones (pp)pGpp using malachite green. Microb. Cell 2024, 11, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Ran, Y.; Xu, H.; Yang, Q.; Xu, Y.; Yang, H.; Qiao, D.; Cao, Y. GATA-type transcriptional factor SpGAT1 interacts with SpMIG1 and promotes lipid accumulation in the oleaginous yeast Saitozyma podzolica zwy-2-3. Biotechnol. Biofuels Bioprod. 2022, 15, 103. [Google Scholar] [CrossRef]
- Peng, Q.; Dong, Y.; Chen, Y.; Glidle, A.; Kong, L.; Yin, H.; Xu, J.; Yang, K. Rapid profiling of fish cell nitrogen metabolism with single-cell Raman spectroscopy: Unveiling enzyme’s role in ammonia detoxification. Talanta 2024, 277, 126389. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Lv, A.-J.; Dong, R.-L.; Li, Y.-C.; Zeng, L.-T.; Wang, Y.; Li, H.-S.; Qi, J.; Wang, H.-H.; Zhang, C.-H.; et al. The research on the synergistic improvement of water retention capacity and eating quality of marinated pork meat by the combination of basic arginine and acidic aspartic acid. Food Chem. 2025, 470, 142649. [Google Scholar] [CrossRef] [PubMed]
- Fachri, M.; Amoah, K.; Huang, Y.; Cai, J.; Alfatat, A.; Ndandala, C.B.; Shija, V.M.; Jin, X.; Bissih, F.; Chen, H. Probiotics and paraprobiotics in aquaculture: A sustainable strategy for enhancing fish growth, health and disease prevention—A review. Front. Mar. Sci. 2024, 11, 1499228. [Google Scholar] [CrossRef]
- Gao, S.; Pan, L.; Huang, F.; Song, M.; Tian, C.; Zhang, M. Metagenomic insights into the structure and function of intestinal microbiota of the farmed Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2019, 499, 109–118. [Google Scholar] [CrossRef]
- T/HSKX 002-2021; Code of Practice for Transportation of Channel Catfish. Hubei Standardization Association: Wuhan, China, 2021.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Jiang, Z.; Zhang, Y.; Leng, L.; Yin, Z.; He, W.; Zeng, X.; Pan, D. Changes in Muscle Quality and Gut Microbiota of Whiteleg Shrimp (Penaeus vannamei) Within a Live Supply Chain. Animals 2025, 15, 1431. https://doi.org/10.3390/ani15101431
Zhang P, Jiang Z, Zhang Y, Leng L, Yin Z, He W, Zeng X, Pan D. Changes in Muscle Quality and Gut Microbiota of Whiteleg Shrimp (Penaeus vannamei) Within a Live Supply Chain. Animals. 2025; 15(10):1431. https://doi.org/10.3390/ani15101431
Chicago/Turabian StyleZhang, Ping, Zian Jiang, Yuwei Zhang, Lele Leng, Ziyi Yin, Weining He, Xiaoqun Zeng, and Daodong Pan. 2025. "Changes in Muscle Quality and Gut Microbiota of Whiteleg Shrimp (Penaeus vannamei) Within a Live Supply Chain" Animals 15, no. 10: 1431. https://doi.org/10.3390/ani15101431
APA StyleZhang, P., Jiang, Z., Zhang, Y., Leng, L., Yin, Z., He, W., Zeng, X., & Pan, D. (2025). Changes in Muscle Quality and Gut Microbiota of Whiteleg Shrimp (Penaeus vannamei) Within a Live Supply Chain. Animals, 15(10), 1431. https://doi.org/10.3390/ani15101431