
Effects of Geographical Origin and Timing of Broodstock Collection on Hatchery Conditioning of the Clam Ruditapes decussatus (L. 1758)
 , , , , and
, , , , and
Simple Summary
Abstract
1. Introduction
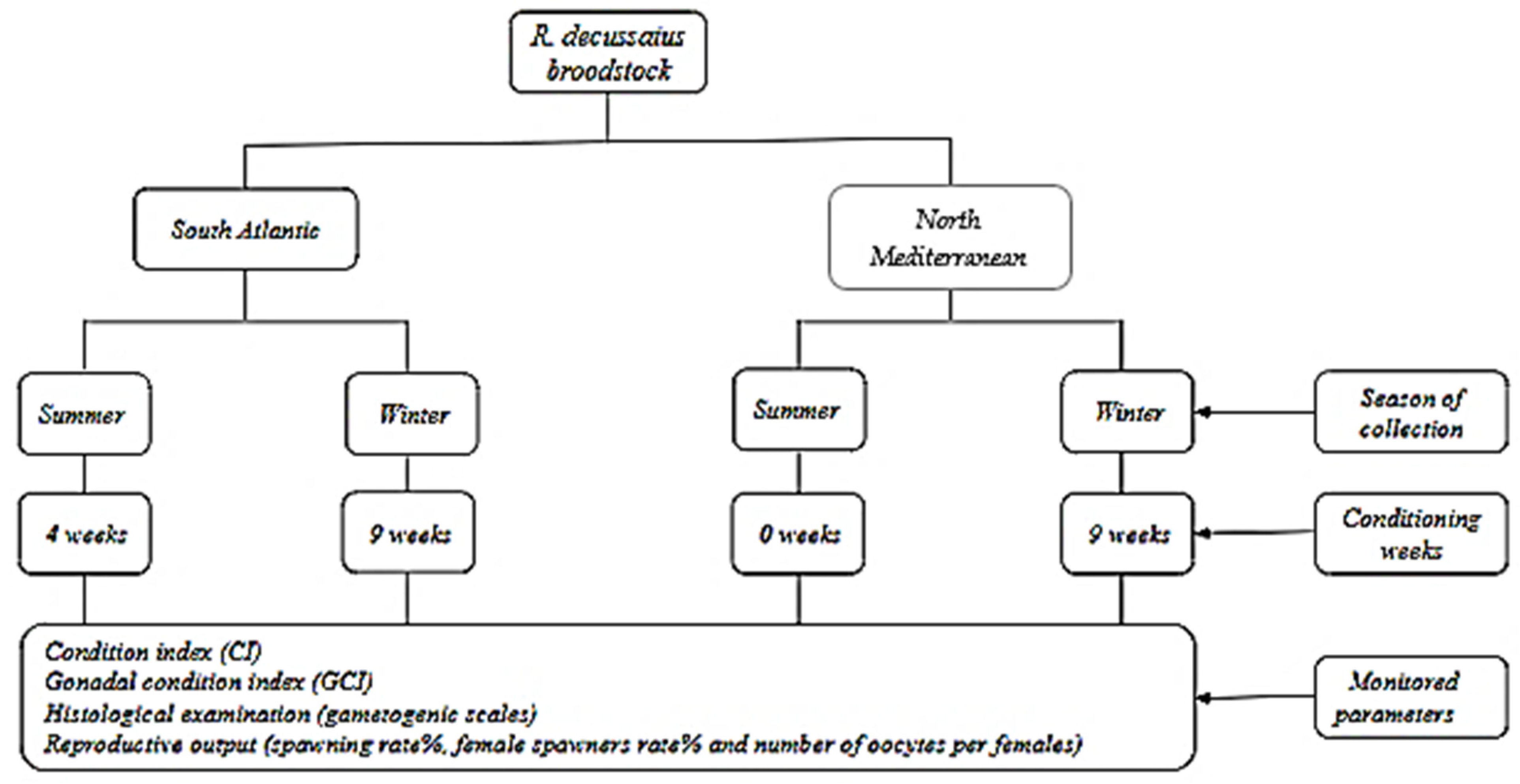
2. Materials and Methods
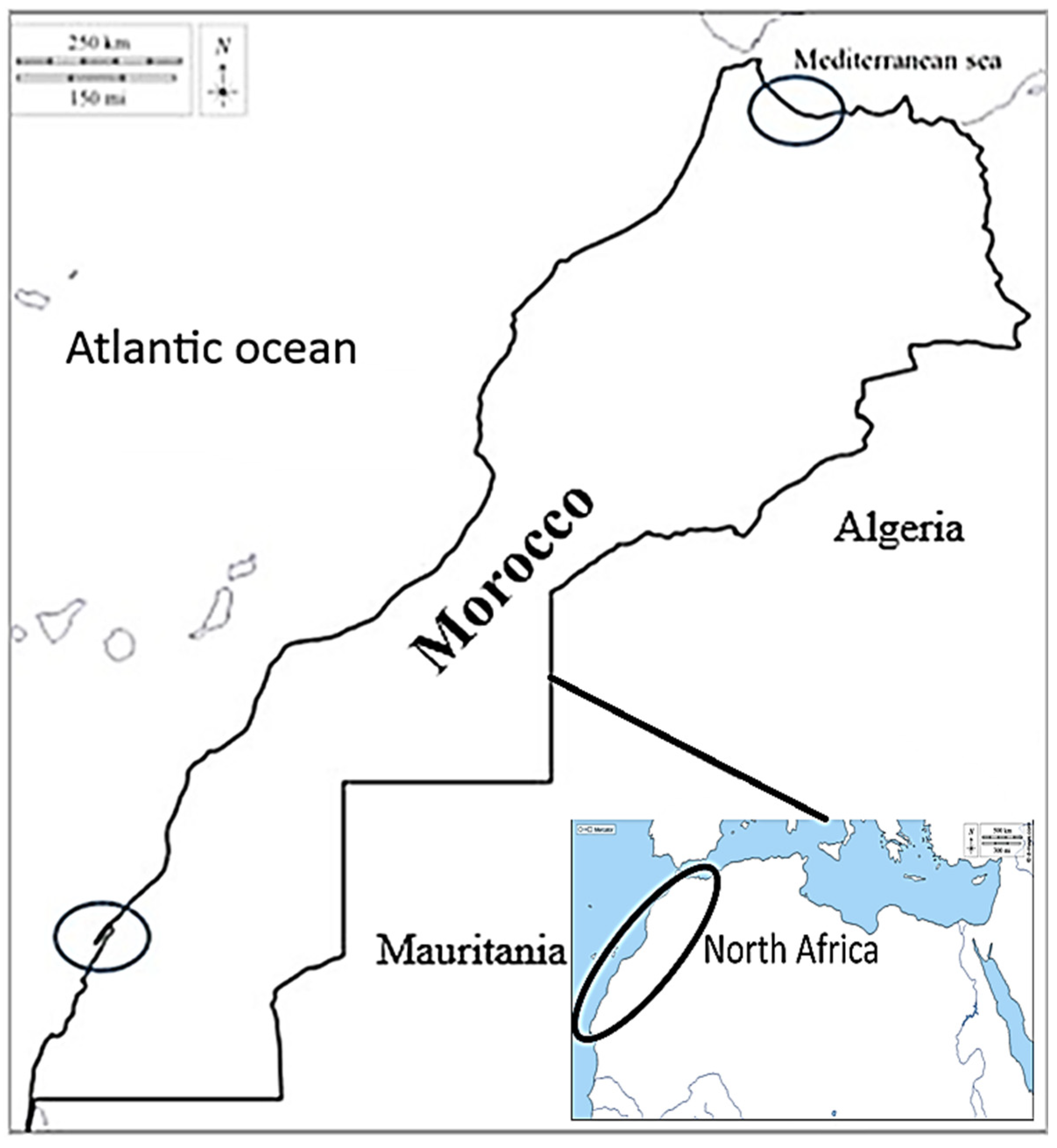
2.1. Broodstock Collection
2.2. Feeding Process
2.3. Broodstock Conditioning
2.4. Physiological Indices
2.5. Histological Examination of the Gonads
2.6. Spawning
2.7. Statistical Analysis
3. Results
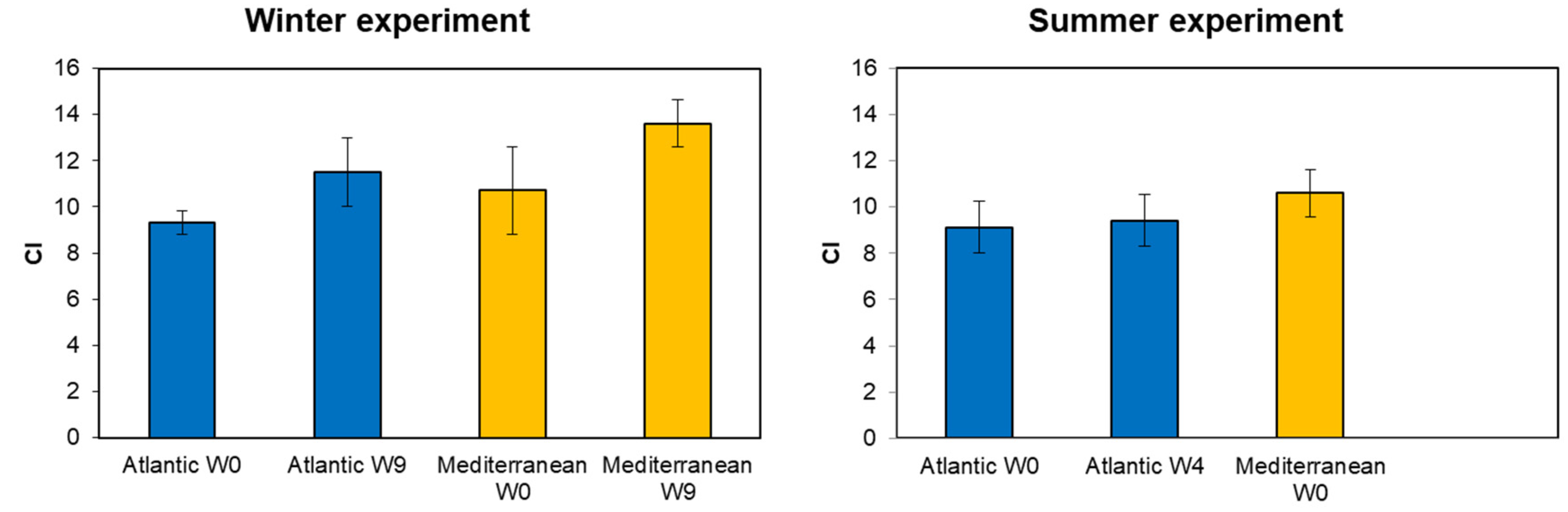
3.1. Condition Index (CI)
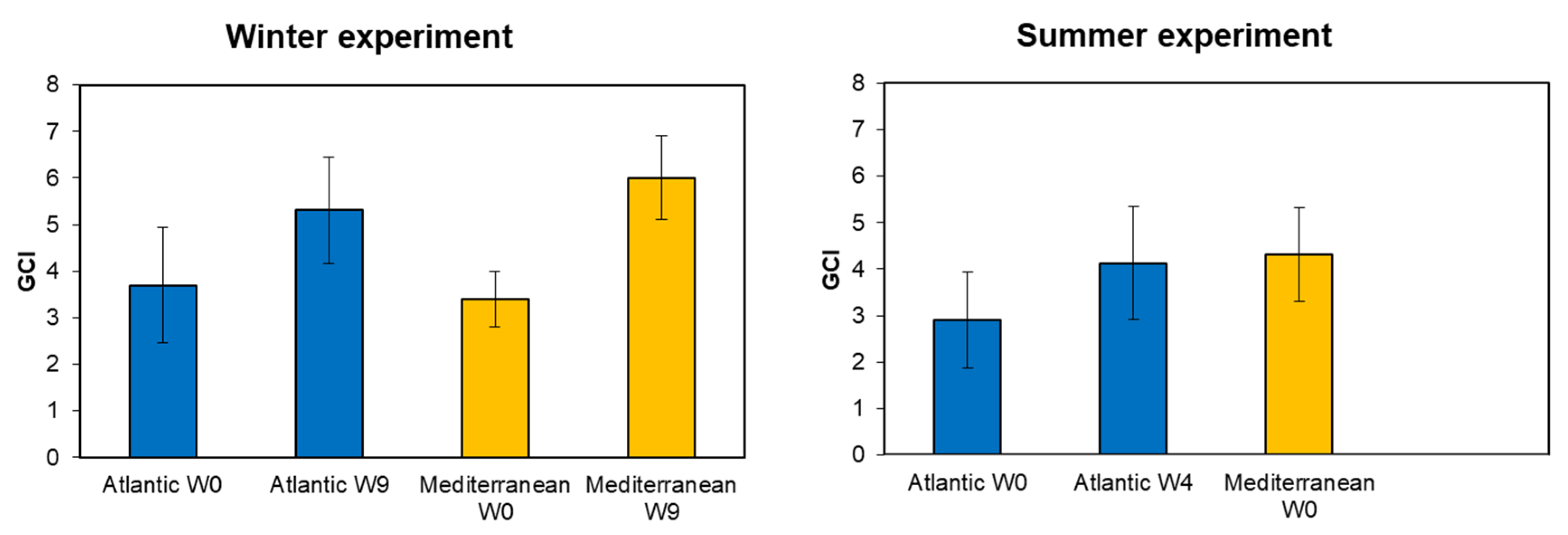
3.2. Gonadal Condition Index (GCI)
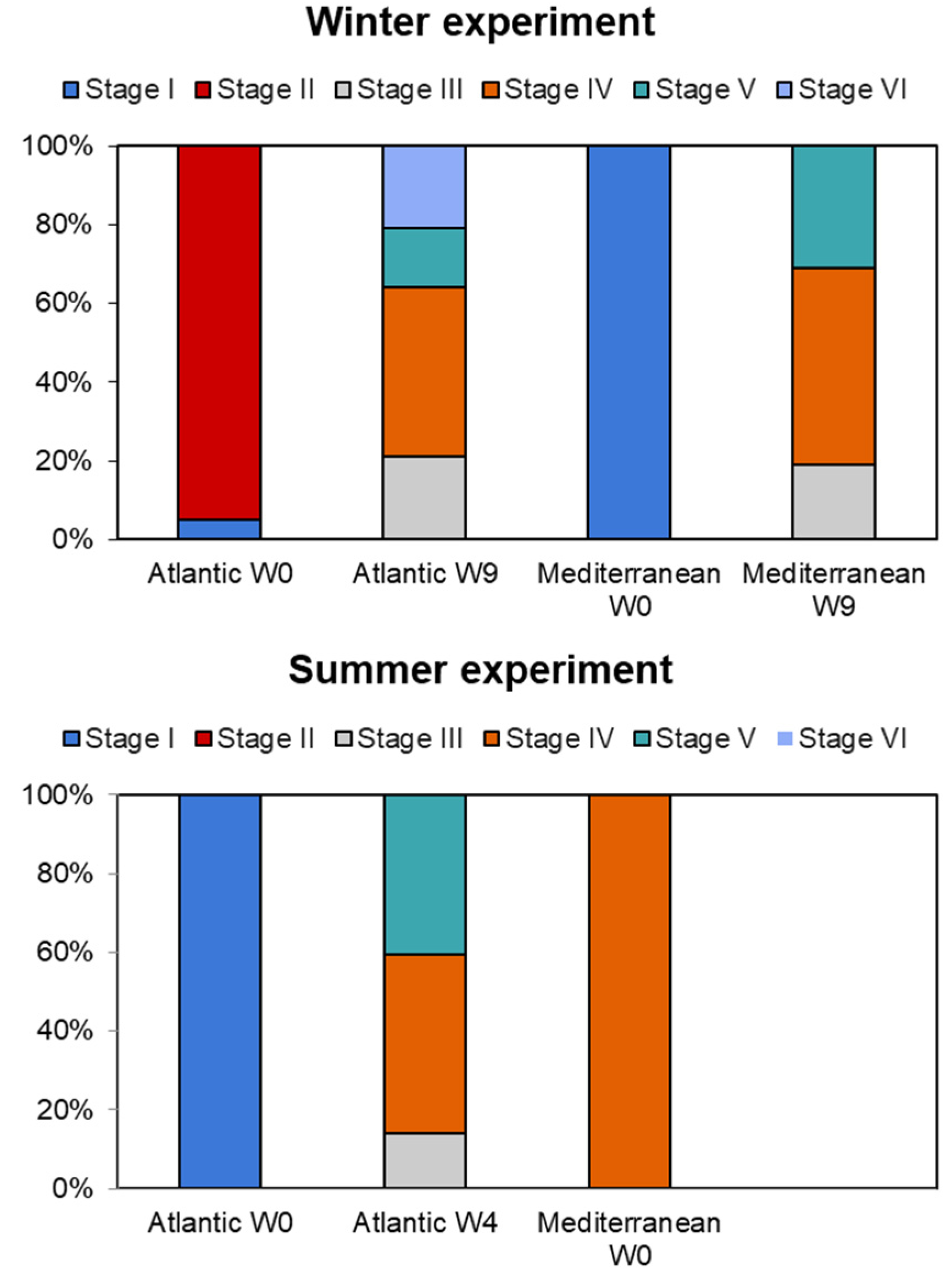
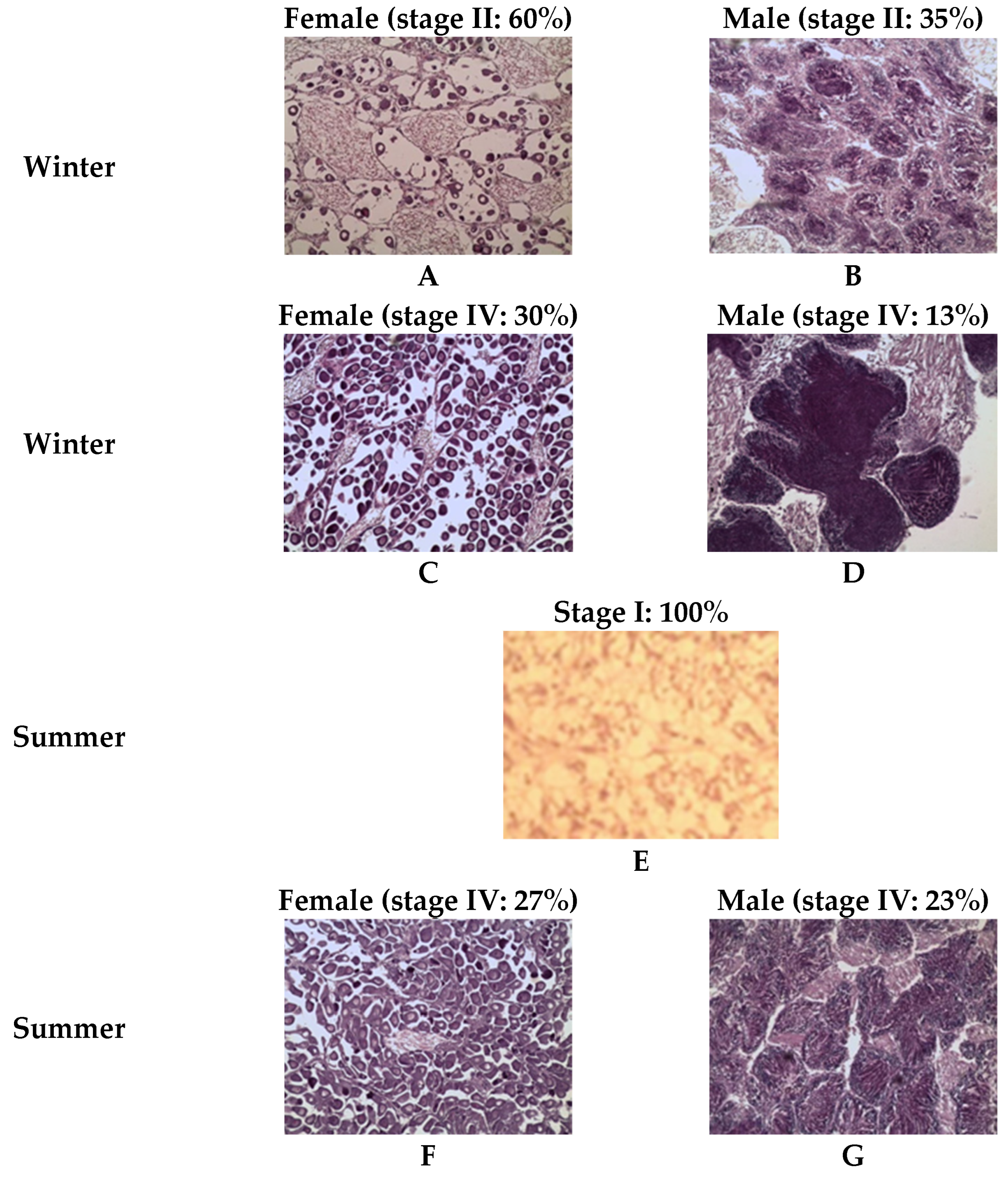
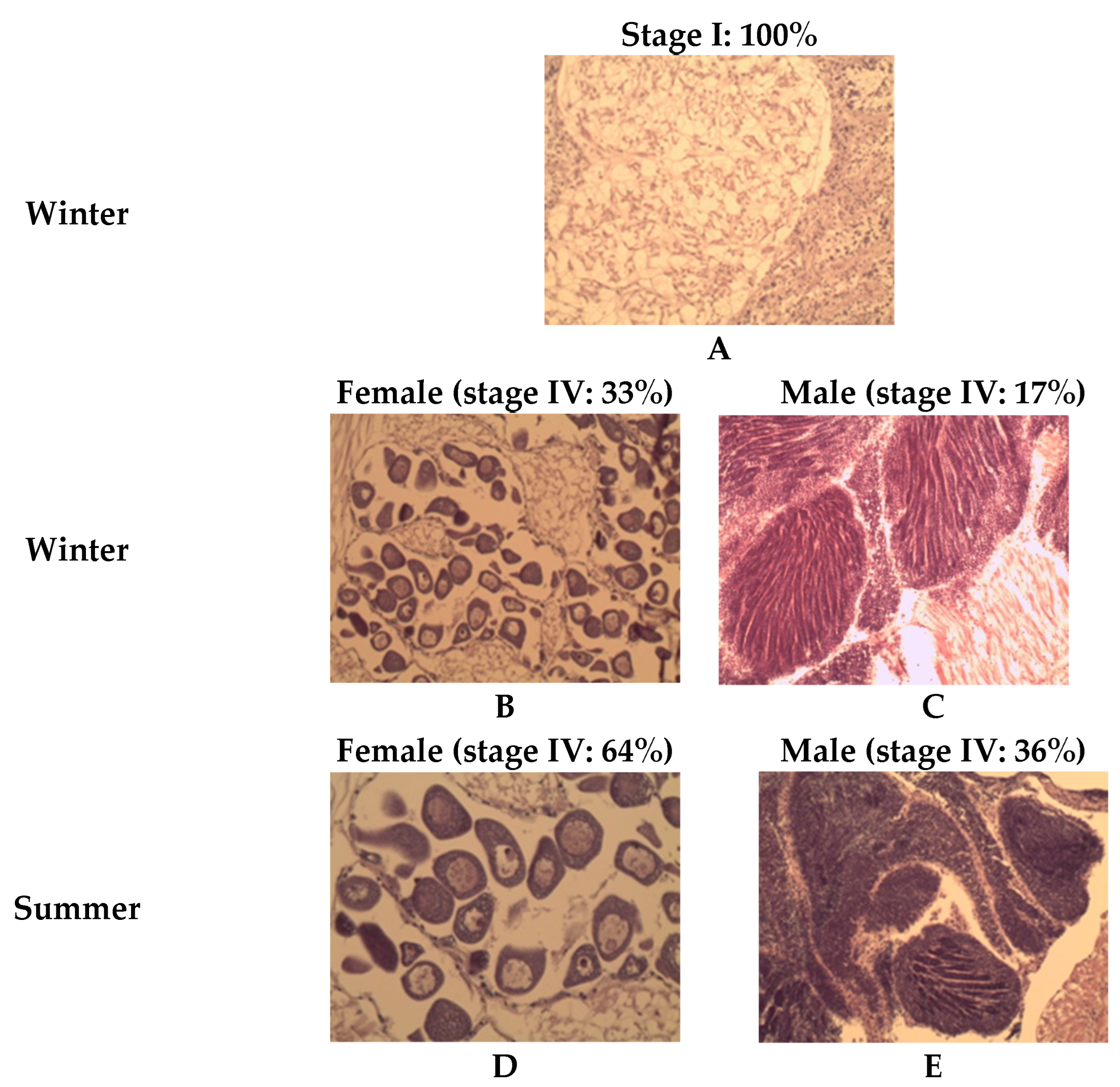
3.3. Gametogenic Scales
Sexual Differentiation and Aspects of Gametogenesis
3.4. Spawning Induction
4. Discussion
4.1. Maturity Development
4.2. Gametogenic Scale
4.3. Reproductive Output
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- da Costa, F.; Cerviño-Otero, A.; Iglesias, Ó.; Cruz, A.; Guévélou, E. Hatchery culture of European clam species (family Veneridae). Aquac. Int. 2020, 28, 1675–1708. [Google Scholar] [CrossRef]
- Fisheries, F.A.O. The State of World Fisheries and Aquaculture. Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar]
- Castilla, J.C.; Defeo, O. Latin American benthic shellfisheries: Emphasis on co-management and experimental practices. Rev. Fish Biol. Fish. 2001, 11, 1–30. [Google Scholar] [CrossRef]
- Fisheries, F.A.O. The State of World Fisheries and Aquaculture. Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2009. [Google Scholar]
- Wendling, C.C.; Huhn, M.; Ayu, N.; Bachtiar, R.; von Juterzenka, K.; Lenz, M. Habitat degradation correlates with tolerance to climate-change related stressors in the green mussel Perna viridis from West Java, Indonesia. Mar. Pollut. Bull. 2013, 71, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.K.; Donaghy, L.; Kang, C.K.; Kang, H.S.; Lee, H.J.; Park, H.S.; Choi, K.S. Substantial changes in hemocyte parameters of Manila clam Ruditapes philippinarum two years after the Hebei Spirit oil spill off the west coast of Korea. Mar. Pollut. Bull. 2016, 108, 171–179. [Google Scholar] [CrossRef]
- Moura, P.; Vasconcelos, P.; Pereira, F.; Chainho, P.; Costa, J.L.; Gaspar, M.B. Reproductive cycle of the Manila clam (Ruditapes philippinarum): An intensively harvested invasive species in the Tagus Estuary (Portugal). J. Mar. Biol. Assoc. United Kingd. 2018, 98, 1645–1657. [Google Scholar] [CrossRef]
- Velez, C.; Figueira, E.; Soares, A.M.; Freitas, R. Effects of seawater temperature increase on economically relevant native and introduced clam species. Mar. Environ. Res. 2017, 123, 62–70. [Google Scholar] [CrossRef]
- Velez, C.; Figueira, E.; Soares, A.M.; Freitas, R. Combined effects of seawater acidification and salinity changes in Ruditapes philippinarum. Aquat. Toxicol. 2016, 176, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Timmins-Schiffman, E.; O’Donnell, M.J.; Friedman, C.S.; Roberts, S.B. Elevated pCO2 causes developmental delay in early larval Pacific oysters, Crassostrea gigas. Mar. Biol. 2013, 160, 1973–1982. [Google Scholar] [CrossRef]
- da Silva, P.M.; Cremonte, F.; Sabry, R.C.; Rosa, R.D.; Cantelli, L.; Barracco, M.A. Presence and histopathological effects of the Parvatrema sp. (Digenea, Gymnophallidae) in the stout razor clam Tagelus plebeius (Bivalvia, Psammobiidae). J. Invertebr. Pathol. 2009, 102, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Gosling, E. Bivalve Molluscs: Biology, Ecology and Culture, 1st ed.; Wiley-Blackwell: Oxford, UK, 2003; pp. 131–161. [Google Scholar] [CrossRef]
- Casas, S.M.; Villalba, A. Study of perkinsosis in the grooved carpet shell clam Ruditapes decussatus in Galicia (NW Spain). III. The effects of Perkinsus olseni infection on clam reproduction. Aquaculture 2012, 356, 40–47. [Google Scholar] [CrossRef]
- Pretto, T.; Zambon, M.; Civettini, M.; Caburlotto, G.; Boffo, L.; Rossetti, E.; Arcangeli, G. Massive mortality in Manila clams (Ruditapes philippinarum) farmed in the Lagoon of Venice, caused by Perkinsus olseni. Bull. Eur. Assoc. Fish. Pathol. 2014, 34, 43–53. [Google Scholar]
- Ruano, F.; Batista, F.M.; Arcangeli, G. Perkinsosis in the clams Ruditapes decussatus and R. philippinarum in the Northeastern Atlantic and Mediterranean Sea: A review. J. Invertebr. Pathol. 2015, 131, 58–67. [Google Scholar] [CrossRef]
- Yoshinaga, T. Experimental evaluation of the impact of Perkinsus olseni on the physiological activities of juvenile Manila clams. J. Shellfish Res. 2018, 37, 29–39. [Google Scholar] [CrossRef]
- Matias, D.; Joaquim, S.; Leitao, A.; Massapina, C. Effect of geographic origin, temperature and timing of broodstock collection on conditioning, spawning success and larval viability of Ruditapes decussatus (Linné, 1758). Aquac. Int. 2009, 17, 257–271. [Google Scholar] [CrossRef]
- Amane, Z.; Tazi, L.; Idhalla, M.; Chlaida, M. Morphometric analysis of European clam Ruditapes decussatus in Morocco. Aquac. Aquar. Conserv. Legis. 2019, 12, 1623–1634. [Google Scholar]
- Chessa, L.A.; Paesanti, F.; Pais, A.; Scardi, M.; Serra, S.; Vitale, L. Perspectives for development of low impact aquaculture in a Western Mediterranean lagoon: The case of the carpet clam Tapes decussatus. Aquac. Int. 2005, 13, 147–155. [Google Scholar] [CrossRef]
- Ojea, J.; Pazos, A.J.; Martínez, D.; Novoa, S.; García-Martínez, P.; Sánchez, J.L.; Abad, M. Effects of temperature regime on broodstock conditioning of Ruditapes decussatus. J. Shellfish Res. 2008, 27, 1093–1100. [Google Scholar] [CrossRef]
- Bald, J.; Sinquin, A.; Borja, A.; Caill-Milly, N.; Duclercq, B.; Dang, C.; de Montaudouin, X. A system dynamics model for the management of the Manila clam, Ruditapes philippinarum (Adams and Reeve, 1850) in the Bay of Arcachon (France). Ecol. Modell. 2009, 220, 2828–2837. [Google Scholar] [CrossRef]
- da Costa, F.; Aranda-Burgos, J.A.; Cerviño-Otero, A.; Fernandez-Pardo, A.; Louzán, A.; Nóvoa, S.; Louzan, A.; Novoa, S.; Ojea, J.; Martínez-Patiño, D. Clam reproduction. In Clam Fisheries and Aquaculture; da Costa, F., Ed.; Nova Science Publishers: New York, NY, USA, 2013; pp. 45–71. ISBN 978-1-62257-518-3. [Google Scholar]
- Ouagajjou, Y.; El Aloua, T.; El Moussaoui, M.; Aghzar, A.; Saoud, Y. First insight into nutrition effect on spawning and larvae rearing of the clam Ruditapes decussatus L. from Dakhla Bay. Aquacult. Aquarium Conserv. Legis. 2020, 13, 3664–3673. [Google Scholar]
- Shafee, M.S.; Daoudi, M. Gametogenesis and spawning in the carpet-shell clam, Ruditapes decussatus (L.) (Mollusca: Bivalvia), from the Atlantic coast of Morocco. Aquac. Res. 1991, 22, 203–216. [Google Scholar] [CrossRef]
- Hamida, L.; Medhiouband, M.N.; Cochard, J.C.; Rhomdhane, M.S.; Le Pennec, M. Étude comparative du cycle de reproduction de la palourde Ruditapes decussatus en milieu naturel (sud Tunisie) et contrôlé (écloserie). Cah. Biol. Mar. 2004, 45, 291–303. [Google Scholar]
- Ojea, J.; Pazos, A.J.; Martınez, D.; Novoa, S.; Sanchez, J.L.; Abad, M. Seasonal variation in weight and biochemical composition of the tissues of Ruditapes decussatus in relation to the gametogenic cycle. Aquaculture 2004, 238, 451–468. [Google Scholar] [CrossRef]
- Kamara, A.; Rharbi, N.; Berraho, A.; Idhalla, M.; Ramdani, M. Comparative study of sexual cycle of the European clam Ruditapes decussatus from three paralic sites off the Moroccan coast. Mar. Life 2005, 15, 43–50. [Google Scholar]
- Helm, M.M.; Bourne, N.; Lovatelli, A. Hatchery Culture of Bivalves: A Practical Manual; FAO: Rome, Italy, 2004. [Google Scholar]
- Library of Congress. Available Online: Morocco (Introduction—Guide to Law Online: Morocco—Research Guides at Library of Congress). Available online: https://guides.loc.gov/law-morocco?loclr=bloglaw (accessed on 20 September 2021).
- Gabbott, P.A. Storage cycles in marine bivalve molluscs: An hypothesis concerning the relation between glycogen and gametogenesis. In Proceedings of the Ninth European Marine Biology Symposium, Oban, Scotland, 2–8 October 1975; Barnes, H., Ed.; Aberdeen University Press: Aberdeen, Scotland, 1975; pp. 191–211. [Google Scholar]
- Bayne, B.L. Marine Mussels: Their Ecology and Physiology, 1st ed.; Cambridge University Press: New York, NY, USA, 1976; pp. 293–337. [Google Scholar]
- Barber, B.J.; Blake, N.J. Substrate catabolism related to reproduction in the bay scallop Argopecten irradians concentricus, as determined by O/N and RQ physiological indexes. Mar. Biol. 1985, 87, 13–18. [Google Scholar] [CrossRef]
- Pauletto, M. Reproduction and Immunology: Transcriptomic Approaches to Improve Bivalves Farming. Ph.D. Thesis, Università Degli Studi di Padova, Padova, Italy, 2014. [Google Scholar]
- Dubert, J.; da Costa, F.; Aranda-Burgos, J.A.; Martínez-Patiño, D.; Prado, S.; Barja, J.L. Beneficial effects of carpet shell clam (Ruditapes decussatus) depuration during short periods of conditioning in shellfish hatchery: Role of the temperature and phytoplankton on reduction and diversity of vibrios. Aquaculture 2016, 459, 65–72. [Google Scholar] [CrossRef]
- Bhatkar, V.R.; Pawase, A.S.; Pair, R.; Shirdhankar, M.S.; Pathan, D. Effect of salinity and temperature on broodstock conditioning of mangrove clam Polymesoda erosa (Solander, 1786). Int. J. Fish. Aquat. Stud. 2021, 9, 340–344. [Google Scholar] [CrossRef]
- Marshall, R.; McKinley, R.S.; Pearce, C.M. Effect of temperature on gonad development of the Pacific geoduck clam (Panopea generosa Gould, 1850). Aquaculture 2012, 338, 264–273. [Google Scholar] [CrossRef]
- Domínguez, L.; Villalba, A.; Fuentes, J. Effects of photoperiod and the duration of conditioning on gametogenesis and spawning of the mussel Mytilus galloprovincialis (Lamarck). Aquac. Res. 2010, 41, e807–e818. [Google Scholar] [CrossRef]
- Maneiro, V.; Silva, A.; Pazos, A.J.; Sánchez, J.L.; Pérez-Parallé, M.L. Effects of temperature and photoperiod on the conditioning of the flat oyster (Ostrea edulis L.) in autumn. Aquac. Res. 2017, 48, 4554–4562. [Google Scholar] [CrossRef]
- Argüello-Guevara, W.; Loor, A.; Sonnenholzner, S. Broodstock conditioning, spawning induction, and early larval development of the tropical rock oyster Striostrea prismatica (Gray 1825). J. Shellfish Res. 2013, 32, 665–670. [Google Scholar] [CrossRef]
- Lannan, J.E.; Robinson, A.; Breese, W.P. Broodstock management of Crassostrea gigas: II. Broodstock conditioning to maximize larval survival. Aquaculture 1980, 21, 337–345. [Google Scholar] [CrossRef]
- Ríos-González, K.G.; López-Uriarte, E.; Chong-Carrillo, O.; Vega-Villasante, F.; Chávez-Villalba, J. Rock oyster Striostrea prismatica (Gray, 1825): Biology, exploitation and conservation. Lat. Am. J. Aquat. Res. 2018, 46, 626–644. [Google Scholar] [CrossRef]
- Albentosa, M.; Pérez-Camacho, A.; Labarta, U.; Fernández-Reiriz, M.J. Evaluation of live microalgal diets for the spat culture of Ruditapes decussatus using physiological and biochemical parameters. Aquaculture 1996, 148, 11–23. [Google Scholar] [CrossRef]
- Abbas, A.S.; El-Wazzan, E.; Khafage, A.R.; El-Sayed, A.F.M.; Abdel Razek, F.A. Influence of different microalgal diets on gonadal development of the carpet shell clam Ruditapes decussatus broodstock. Aquac. Int. 2018, 26, 1297–1309. [Google Scholar] [CrossRef]
- Brown, M.R. Nutritional value of microalgae for aquaculture. In Advances en Nutrición Acuícola VI. Proceedings of the Memorias del VI Simposium International de Nutrición Acuícocla, Cancún, Quintana Roo, México, 3–6 Septiembre 2002; Cruz-Suárez, L.E., RicqueMarie, D., Tapia-Salazar, M., Gaxiola-Cortés, M.G., Simoes, N., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2002. [Google Scholar] [CrossRef]
- Ouagajjou, Y.; Aghzar, A.; Ahensal, M.; Berday, N.; Idhalla, M. Behavioural pattern in changes to diets and food availability during broodstock conditioning of captive clam Ruditapes decussates. E3S Web Conf. 2022, 337, 03002. [Google Scholar] [CrossRef]
- Utting, S.D.; Millican, P.F. Techniques for the hatchery conditioning of bivalve broodstocks and the subsequent effect on egg quality and larval viability. Aquaculture 1997, 155, 45–54. [Google Scholar] [CrossRef]
- Walne, P.R.; Mann, R. Growth and biochemical composition in Ostrea edulis and Crassostrea gigas. In Proceedings of the Ninth European Marine Biology Symposium, Oban, Scotland, 2–8 October 1975; Aberdeen University Press: Scotland, UK, 1975; pp. 587–607. [Google Scholar]
- Delgado, M.; Pérez-Camacho, A. Histological study of the gonadal development of Ruditapes decussatus (L.) (Mollusca: Bivalvia) and its relationship with available food. Sci. Mar. 2005, 69, 87–97. [Google Scholar] [CrossRef]
- Laruelle, F.; Guillou, J.; Paulet, Y.M. Reproductive pattern of the clams, Ruditapes decussatus and R. philippinarum on intertidal flats in Brittany. J. Mar. Biol. Assoc. UK 1994, 74, 351–366. [Google Scholar] [CrossRef]
- Serdar, S.; Lök, A. Gametogenic cycle and biochemical composition of the transplanted carpet shell clam Tapes decussatus, Linnaeus 1758 in Sufa (Homa) Lagoon, Izmir, Turkey. Aquaculture 2009, 293, 81–88. [Google Scholar] [CrossRef]
- Boudry, P.; Collet, B.; Cornette, F.; Hervouet, V.; Bonhomme, F. High variance in reproductive success of the Pacific oyster (Crassostrea gigas, Thunberg) revealed by microsatellite-based parentage analysis of multifactorial crosses. Aquaculture 2002, 204, 283–296. [Google Scholar] [CrossRef]
- Figueras, A. Moluscos de las playas de la ría de Vigo: II. Crecimiento y reproducción. Investig. Pesq. 1975, 7, 49–97. [Google Scholar]
- Pérez-Camacho, A. Biología de Venerupis pullastra (Montagu, 1803) y Venerupis decussata (Linné, 1767) (Mollusca: Bivalvia) con especial referencia a factores determinantes de la producción. Bol. Inst. Esp. Oceanogr. 1980, 281, 353–358. [Google Scholar]
- Villalba, A.; Carballal, M.J.; López, M.C. Estudio del ciclo gonadal de tres especies de almeja, Ruditapes decussatus, Venerupis pullastra y Venerupis rhomboides de las rías gallegas. In Actas del IV Congreso nacional de Acuicultura; Centro de Investigacións Mariñas: Vilanova de Arousa, Spain, 1993; pp. 341–346. [Google Scholar]
- Breber, P. Annual gonadal cycle in the carpet-shell clam Venerupis decussata in Venice Lagoon, Italy. Proc. Nat. Shellfish. Assoc. 1980, 70, 31–35. [Google Scholar]
- Xie, Q.; Burnell, G.M. A comparative study of the gametogenic cycles of the clams Tapes philippinarum (Adams and Reeve, 1850) and Tapes decussatus (Linnaeus) on the south coast of Ireland. J. Shellfish Res. 1994, 13, 467–472. [Google Scholar]
- Borsa, P.; Millet, B. Recruitment of the clam Ruditapes decussatus in the Lagoon of Thau, mediterranean. Estuar. Coast. Shelf Sci. 1992, 35, 289–300. [Google Scholar] [CrossRef]
- Matias, D.; Ben-Hamadou, R.; Joaquim, S.; Matias, A.M.; Sobral, P.; Leitao, A. The influence of different microalgal diets on European clam (Ruditapes decussatus, Linnaeus, 1758) larvae culture performances. Aquac. Int. 2015, 46, 2527–2543. [Google Scholar] [CrossRef]
- Mann, R. The effect of temperature on growth, physiology, and gametogenesis in the Manila clam Tapes philippinarum (Adams & Reeve, 1850). J. Exp. Mar. Biol. Ecol. 1979, 38, 121–133. [Google Scholar] [CrossRef]
- Beninger, P.G.; Lucas, A. Seasonal variations in condition, reproductive activity, and gross biochemical composition of two species of adult clam reared in a common habitat: Tapes decussatus L. (Jeffreys) and Tapes philippinarum (Adams & Reeve). J. Exp. Mar. Biol. Ecol. 1984, 79, 19–37. [Google Scholar] [CrossRef]
- Pronker, A.E.; Nevejan, N.M.; Peene, F.; Geijsen, P.; Sorgeloos, P. Hatchery broodstock conditioning of the blue mussel Mytilus edulis (Linnaeus 1758). Part I. Impact of different micro-algae mixtures on broodstock performance. Aquac. Int. 2008, 16, 297–307. [Google Scholar] [CrossRef]
- Joaquim, S.; Matias, D.; Matias, A.M.; Moura, P.; Roque, C.; Chícharo, L.; Gaspar, M.B. Biochemical and energy dynamics throughout the reproductive cycle of the striped venus Chamelea gallina (Mollusca, Bivalvia). Invertebr. Reprod. Dev. 2014, 58, 284–293. [Google Scholar] [CrossRef]
- Honkoop, P.J.C.; Beukema, J.J. Loss of body mass in winter in three intertidal bivalve species: An experimental and observational study of the interacting effects between water temperature, feeding time and feeding behaviour. J. Exp. Mar. Biol. Ecol. 1997, 212, 277–297. [Google Scholar] [CrossRef]
- Rato, A.; Joaquim, S.; Matias, A.M.; Roque, C.; Marques, A.; Matias, D. The impact of climate change on bivalve farming: Combined effect of temperature and salinity on survival and feeding behavior of clams Ruditapes decussatus. Front. Mar. Sci. 2022, 9, 932310. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Spawning Parameters | Experiment (Season of Broodstock Collection) | |
|---|---|---|---|
| Winter | Summer | ||
| South Atlantic | Spawners (%) | 69 | 40 |
| Female spawners (%) Oocytes per female | 56 1.68 ± 0.23 (106) | 17 0.84 ± 0.11 (106) | |
| North Mediterranean | Spawners (%) | 42 | 42 |
| Female spawners (%) Oocytes per female | 19 2.34 ± 0.28 (106) | 18 0.11 ± 0.10 (106) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azirar, R.; Fettach, S.; da Costa, F.; Pérez, M.; Chiaar, A.; Aghzar, A.; Ouagajjou, Y. Effects of Geographical Origin and Timing of Broodstock Collection on Hatchery Conditioning of the Clam Ruditapes decussatus (L. 1758). Animals 2025, 15, 29. https://doi.org/10.3390/ani15010029
Azirar R, Fettach S, da Costa F, Pérez M, Chiaar A, Aghzar A, Ouagajjou Y. Effects of Geographical Origin and Timing of Broodstock Collection on Hatchery Conditioning of the Clam Ruditapes decussatus (L. 1758). Animals. 2025; 15(1):29. https://doi.org/10.3390/ani15010029
Chicago/Turabian StyleAzirar, Rania, Samah Fettach, Fiz da Costa, Montse Pérez, Abderrahim Chiaar, Adil Aghzar, and Yassine Ouagajjou. 2025. "Effects of Geographical Origin and Timing of Broodstock Collection on Hatchery Conditioning of the Clam Ruditapes decussatus (L. 1758)" Animals 15, no. 1: 29. https://doi.org/10.3390/ani15010029
APA StyleAzirar, R., Fettach, S., da Costa, F., Pérez, M., Chiaar, A., Aghzar, A., & Ouagajjou, Y. (2025). Effects of Geographical Origin and Timing of Broodstock Collection on Hatchery Conditioning of the Clam Ruditapes decussatus (L. 1758). Animals, 15(1), 29. https://doi.org/10.3390/ani15010029