
Exploring Bile-Acid Changes and Microflora Profiles in Chicken Fatty Liver Disease Model

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sample Collection
2.3. Bile-Acid Extraction and Analysis
2.4. RNA Extraction and Real-Time PCR Analysis
2.5. DNA Extraction of and 16S rDNA Amplicon Pyrosequencing
2.6. Sequence Analysis
2.7. Statistical Analysis
3. Results
3.1. Bile-Acid Profile in Serum and Cecal Contents
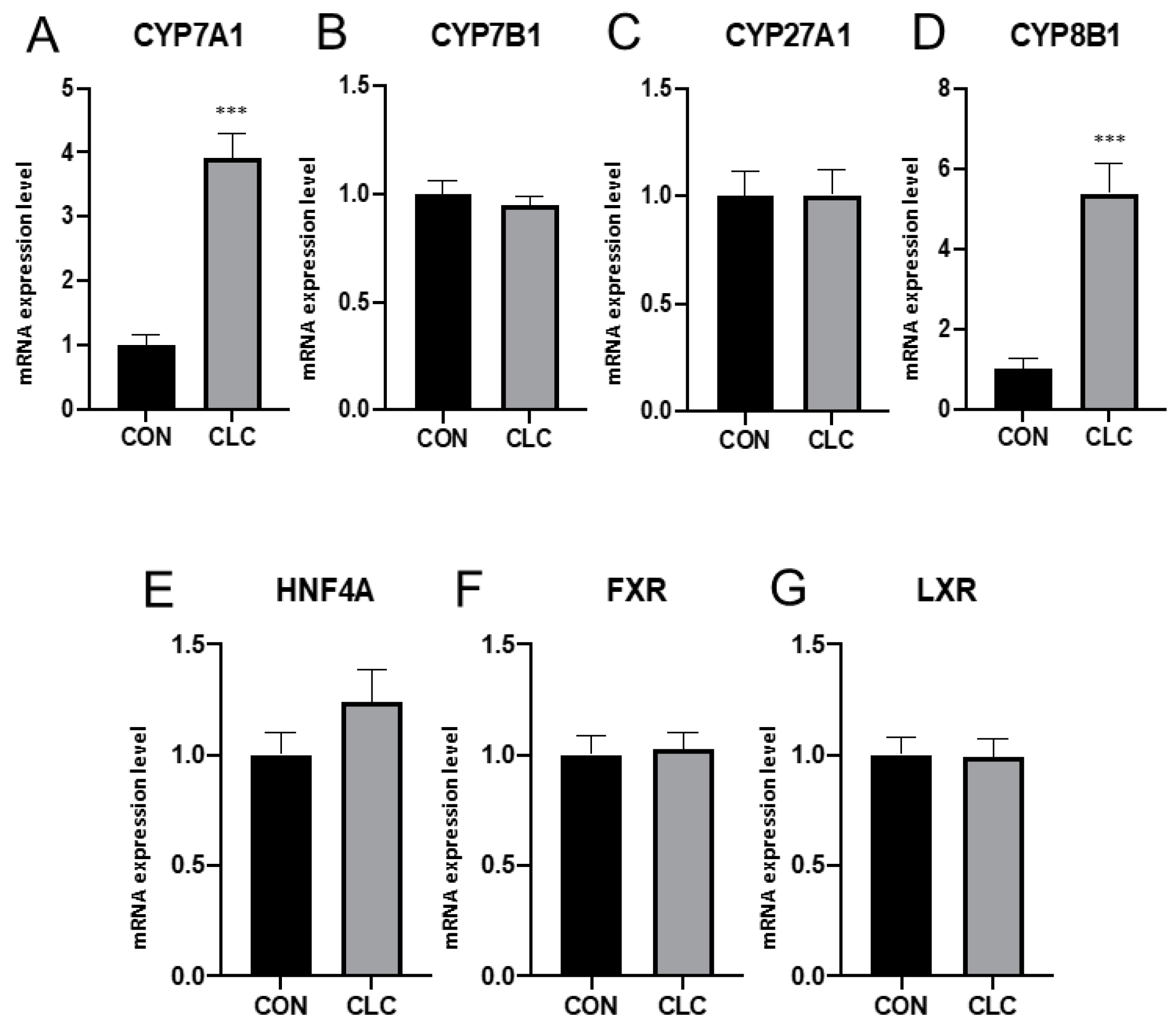
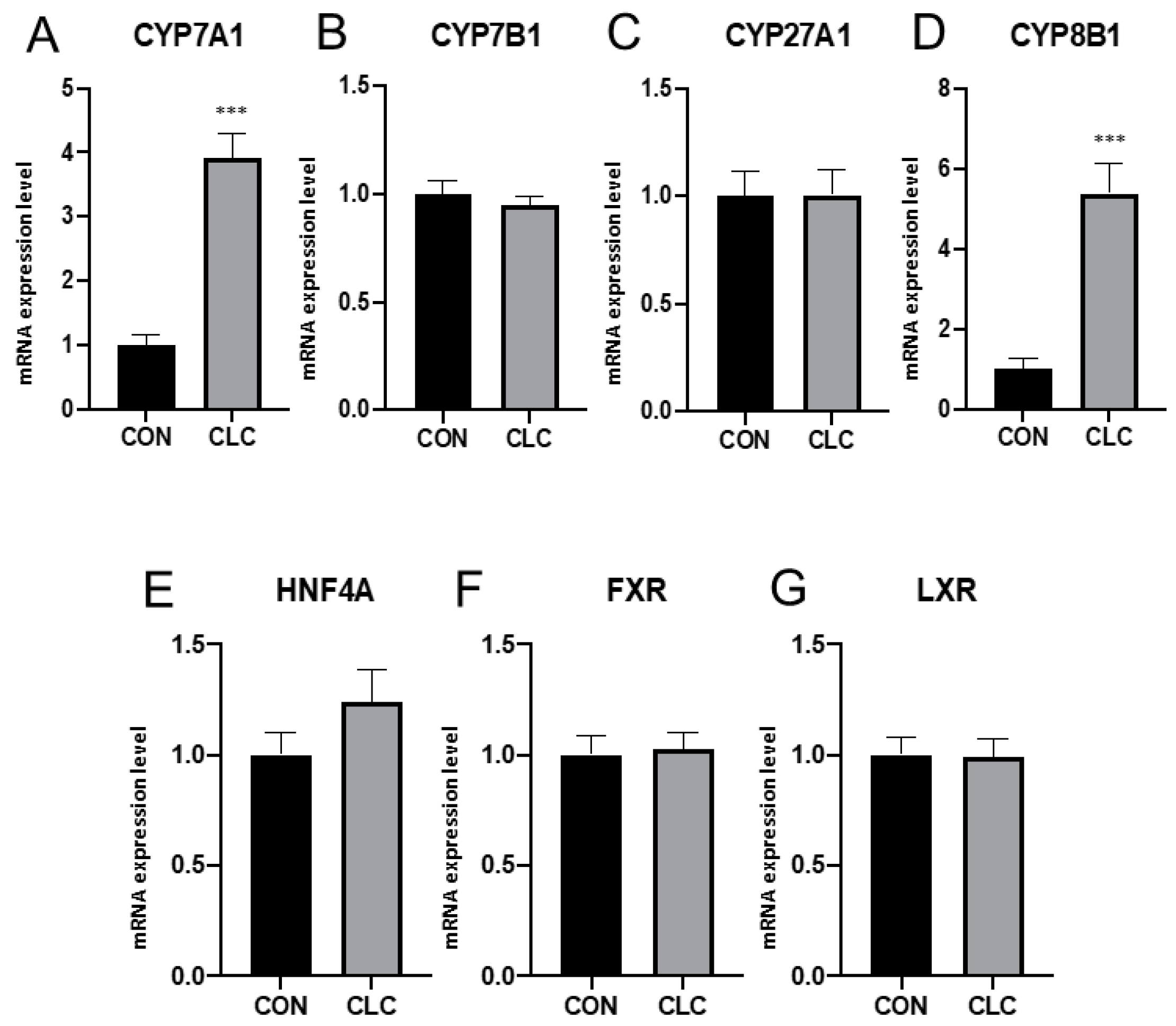
3.2. Hepatic Gene Expression
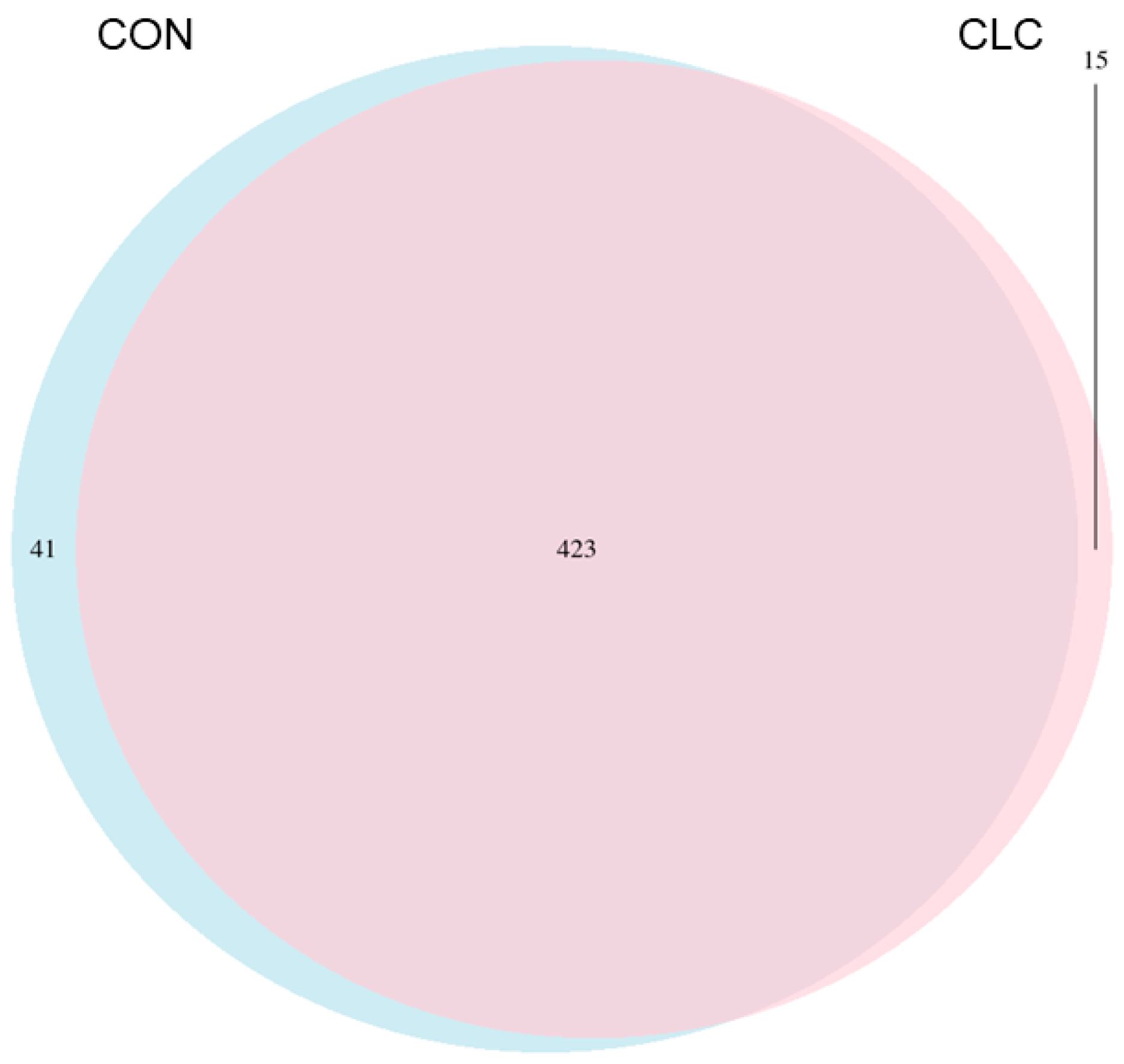
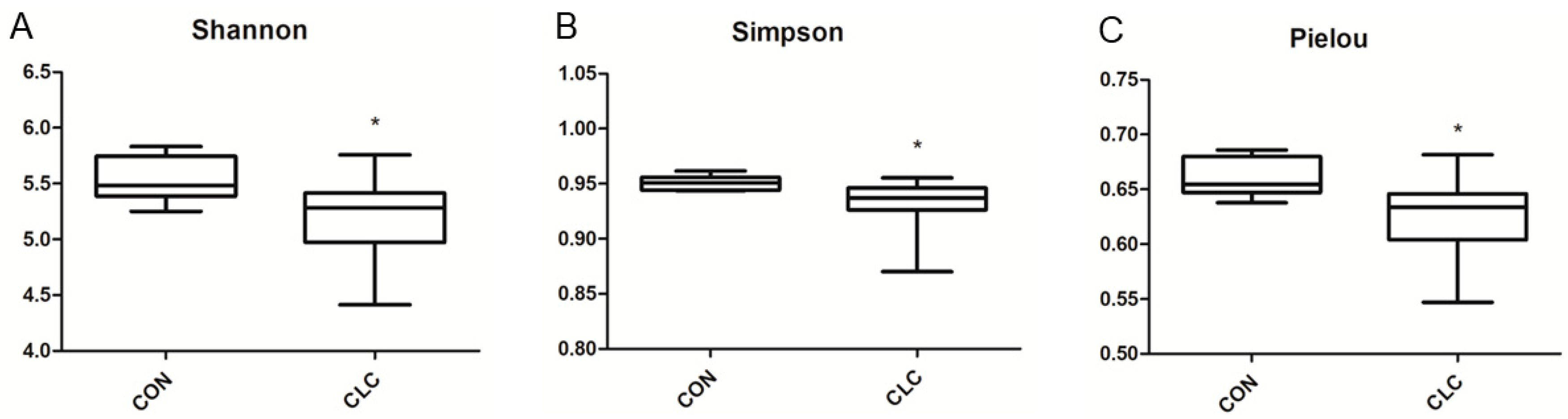
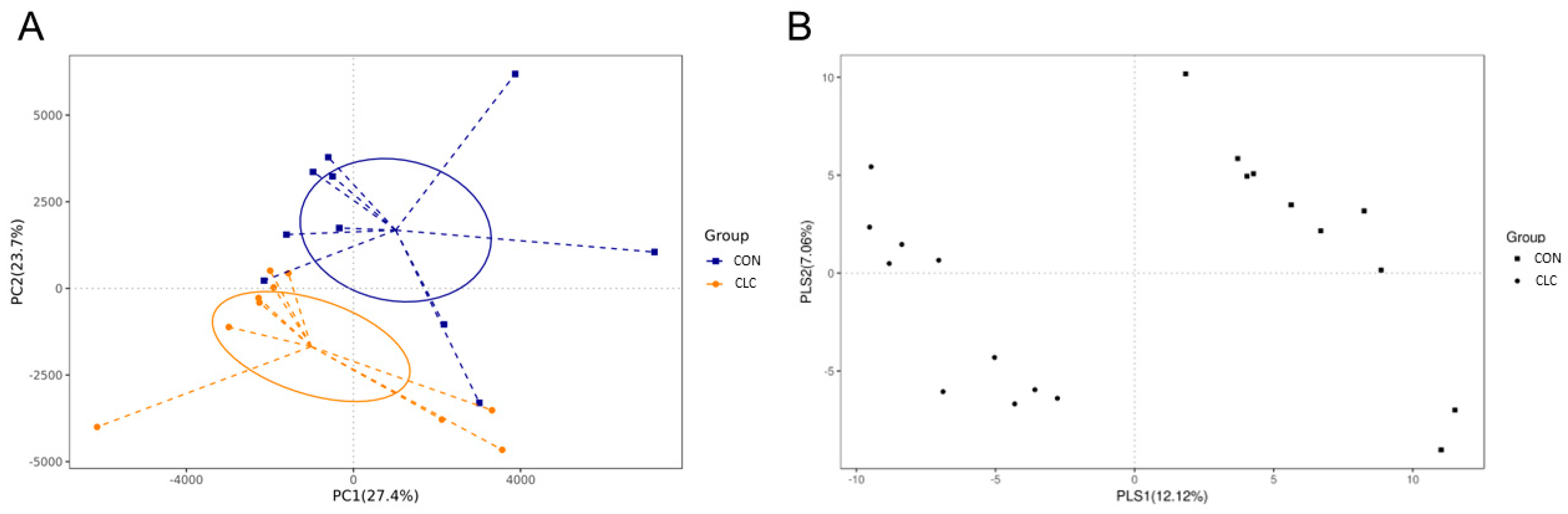
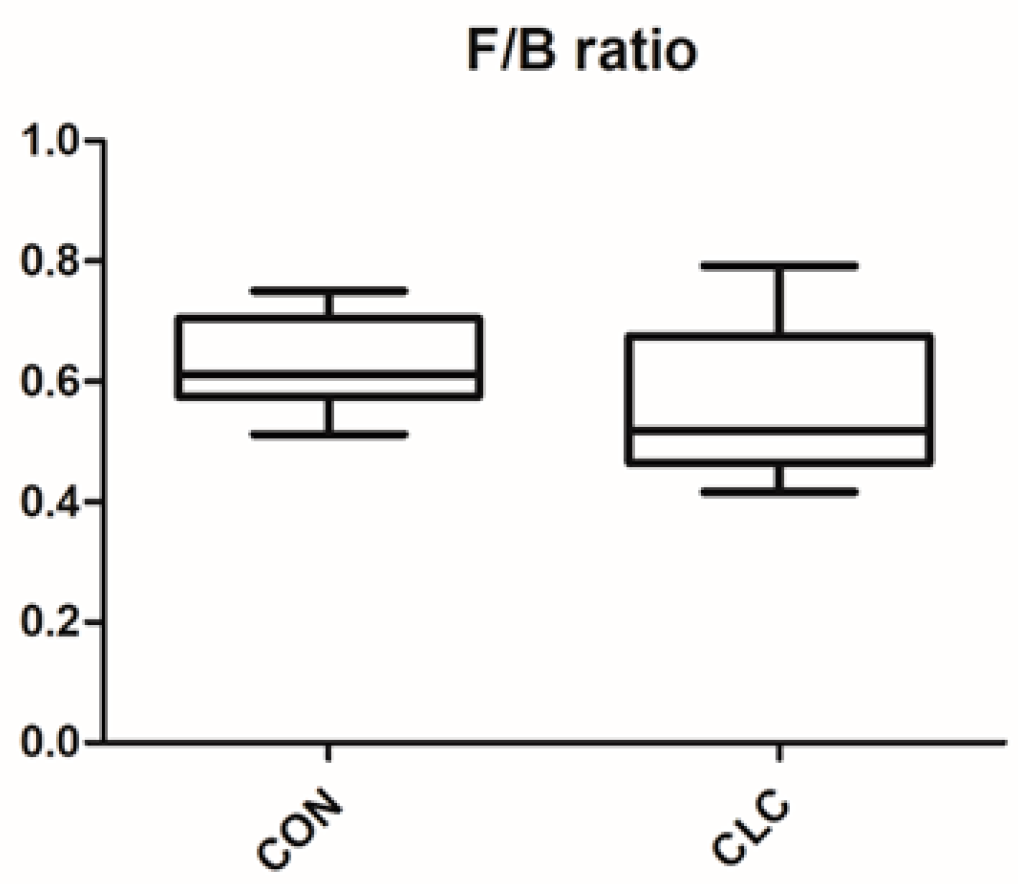
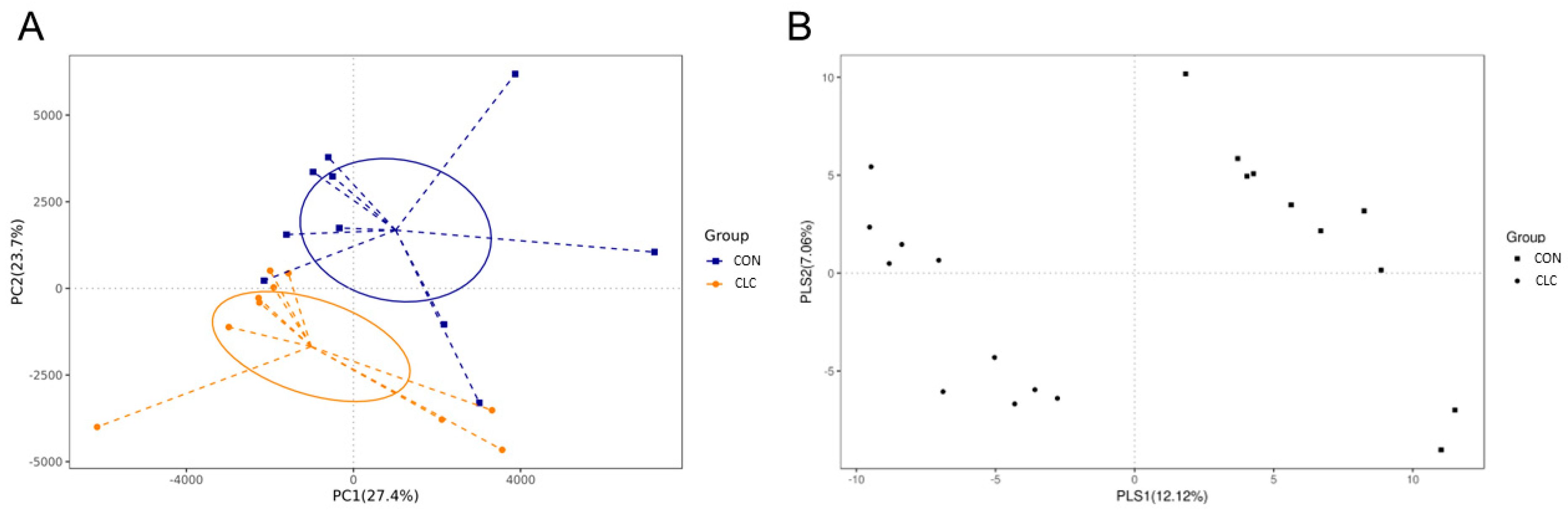
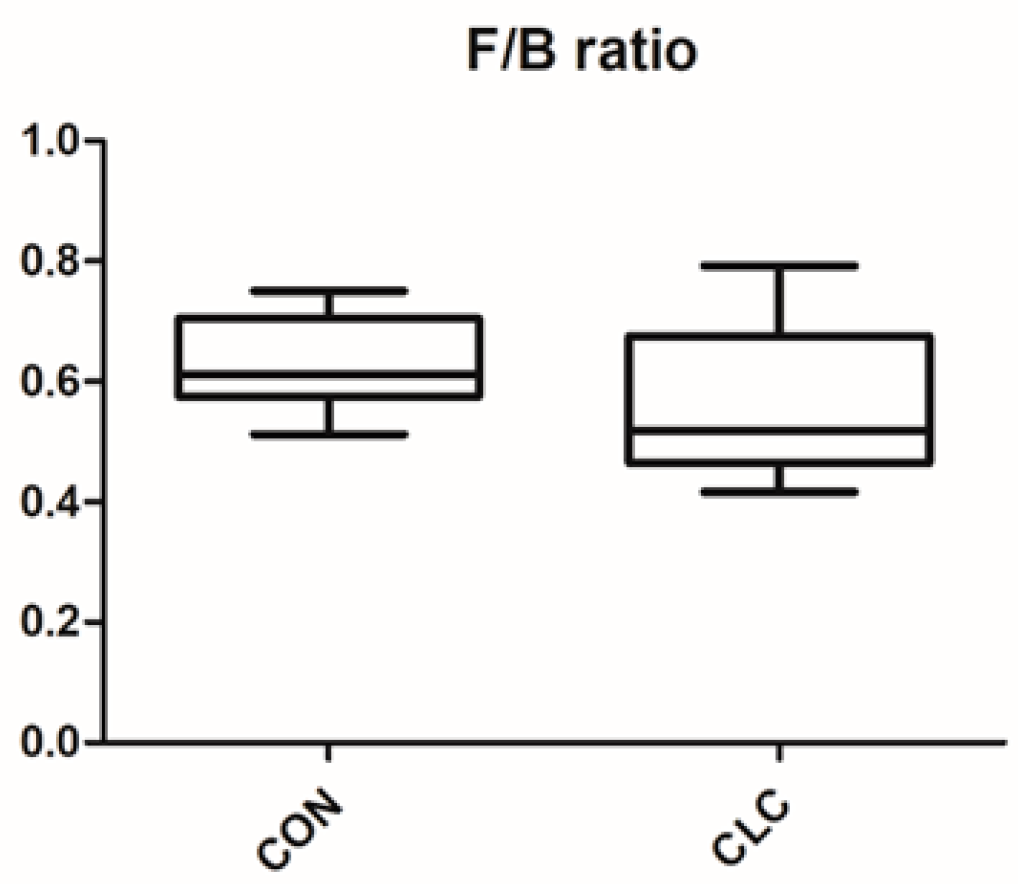
3.3. Effect of CLC Diet on Cecal Digesta Microbiota
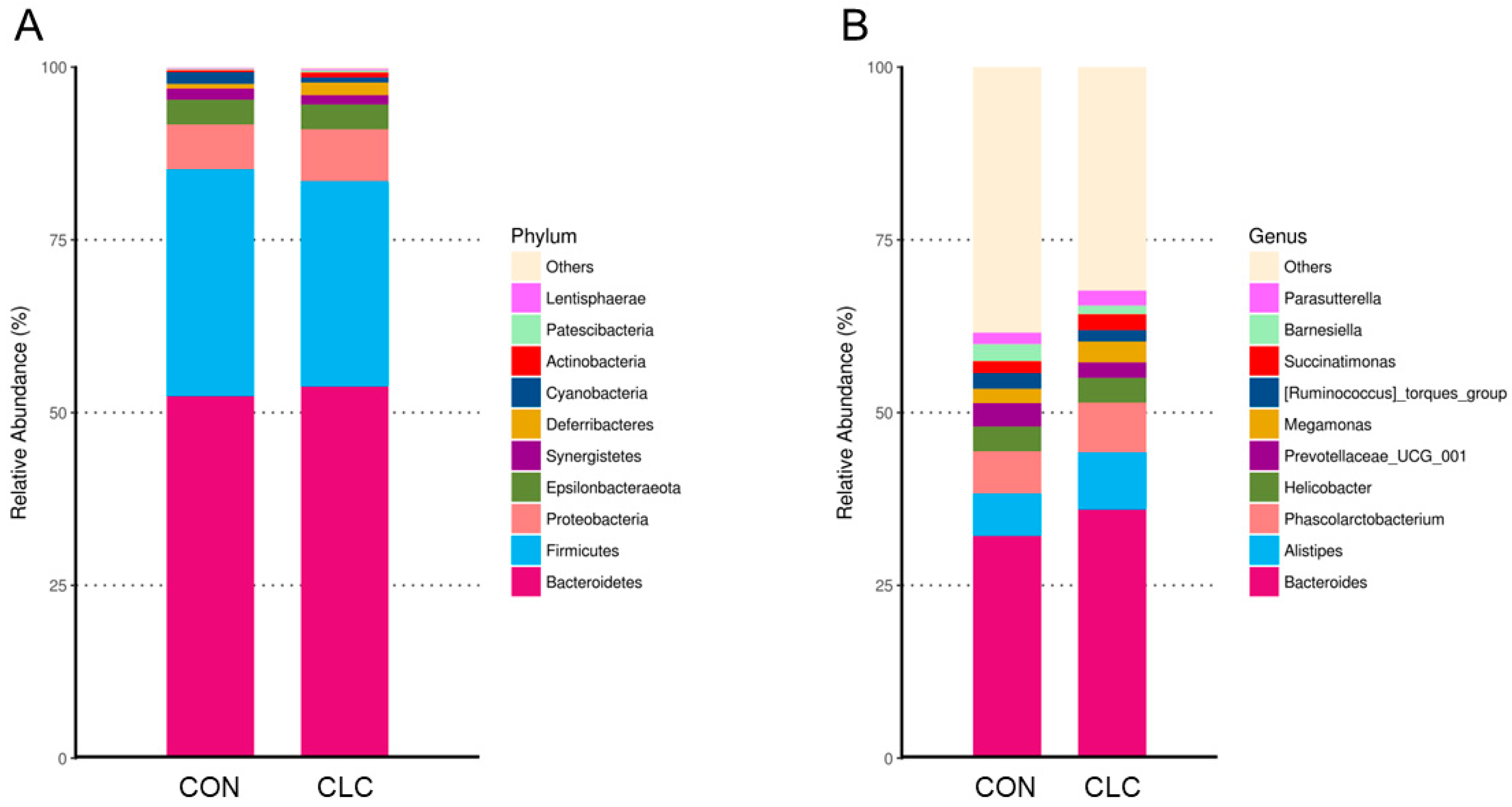
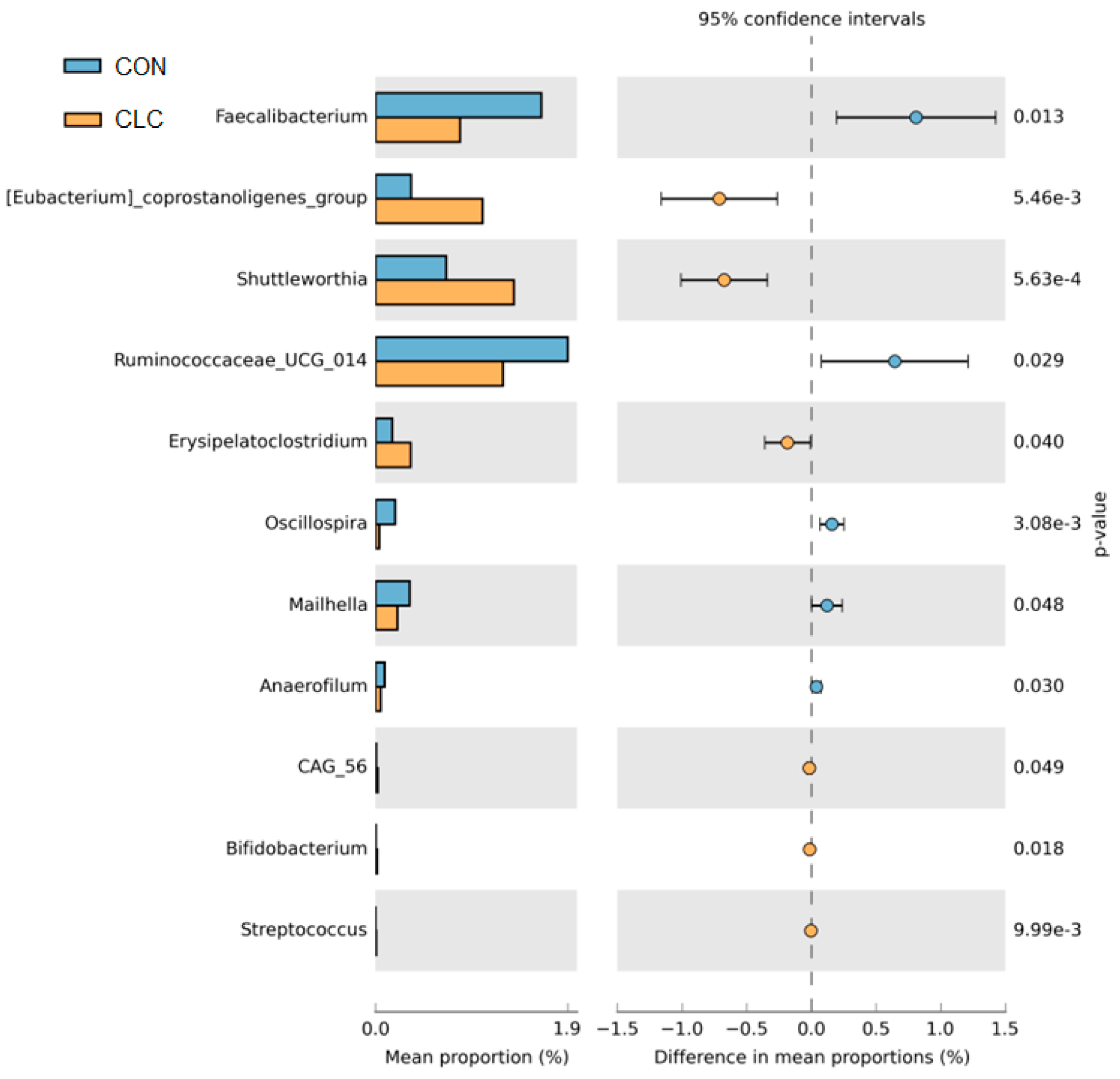
3.4. Effect of CLC Diet on Bacterial Taxonomic Composition of Cecal Digesta
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cherian, G.; Holsonbake, T.B.; Goeger, M.P.; Bildfell, R. Dietary CLA alters yolk and tissue FA composition and hepatic histopathology of laying hens. Lipids 2002, 37, 751–757. [Google Scholar] [CrossRef]
- Krogdahl, A. Digestion and absorption of lipids in poultry. J. Nutr. 1985, 115, 675–685. [Google Scholar] [CrossRef]
- Sklan, D.; Geva, A.; Budowski, P.; Hurwitz, S. Intestinal absorption and plasma transport of lipids in chicks and rats. Comp. Biochem. Physiol. A Comp. Physiol. 1984, 78, 507–510. [Google Scholar] [CrossRef]
- Butler, E.J. Fatty liver diseases in the domestic fowl—A review. Avian Pathol. 1976, 5, 1–14. [Google Scholar] [CrossRef]
- Julian, R.J. Production and growth related disorders and other metabolic diseases of poultry—A review. Vet. J. 2005, 169, 350–369. [Google Scholar] [CrossRef]
- Trott, K.A.; Giannitti, F.; Rimoldi, G.; Hill, A.; Woods, L.; Barr, B.; Anderson, M.; Mete, A. Fatty liver hemorrhagic syndrome in the backyard chicken: A retrospective histopathologic case series. Vet. Pathol. 2014, 51, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Liu, L.; He, Y.; Dou, T.; Jia, J.; Ge, C. Endocrine and genetic factors affecting egg laying performance in chickens: A review. Br. Poult. Sci. 2020, 61, 538–549. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.W.; Huang, T.W.; Peng, Y.J.; Lin, Y.Y.; Mersmann, H.J.; Ding, S.T. A novel chicken model of fatty liver disease induced by high cholesterol and low choline diets. Poult. Sci. 2021, 100, 100869. [Google Scholar] [CrossRef] [PubMed]
- Shini, A.; Shini, S.; Bryden, W.L. Fatty liver haemorrhagic syndrome occurrence in laying hens: Impact of production system. Avian Pathol. 2019, 48, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Thomson, A.E.; Gentry, P.A.; Squires, E.J. Comparison of the coagulation profile of fatty liver haemorrhagic syndrome-susceptible laying hens and normal laying hens. Br. Poult. Sci. 2003, 44, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Xing, C.; Cao, H.; Zhang, C.; Luo, J.; Guo, X.; Hu, G. Insulin resistance and metabonomics analysis of fatty liver haemorrhagic syndrome in laying hens induced by a high-energy low-protein diet. Sci. Rep. 2019, 9, 10141. [Google Scholar] [CrossRef]
- Schwarz, M.; Lund, E.G.; Setchell, K.D.; Kayden, H.J.; Zerwekh, J.E.; Björkhem, I.; Herz, J.; Russell, D.W. Disruption of cholesterol 7α-hydroxylase gene in mice. II. Bile acid deficiency is overcome by induction of oxysterol 7α-hydroxylase. J. Biol. Chem. 1996, 271, 18024–18031. [Google Scholar] [CrossRef] [PubMed]
- Jiao, N.; Baker, S.S.; Chapa-Rodriguez, A.; Liu, W.; Nugent, C.A.; Tsompana, M.; Mastrandrea, L.; Buck, M.J.; Baker, R.D.; Genco, R.J.; et al. Suppressed hepatic bile acid signalling despite elevated production of primary and secondary bile acids in NAFLD. Gut 2018, 67, 1881–1891. [Google Scholar] [CrossRef] [PubMed]
- Aranha, M.M.; Cortez-Pinto, H.; Costa, A.; da Silva, I.B.; Camilo, M.E.; de Moura, M.C.; Rodrigues, C.M. Bile acid levels are increased in the liver of patients with steatohepatitis. Eur. J. Gastroenterol. Hepatol. 2008, 20, 519–525. [Google Scholar] [CrossRef]
- Ferslew, B.C.; Xie, G.; Johnston, C.K.; Su, M.; Stewart, P.W.; Jia, W.; Brouwer, K.L.; Barritt, A.S. Altered Bile Acid Metabolome in Patients with Nonalcoholic Steatohepatitis. Dig. Dis. Sci. 2015, 60, 3318–3328. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- di Gregorio, M.C.; Cautela, J.; Galantini, L. Physiology and Physical Chemistry of Bile Acids. Int. J. Mol. Sci. 2021, 22, 1780. [Google Scholar] [CrossRef]
- Li, J.; Dawson, P.A. Animal models to study bile acid metabolism. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 895–911. [Google Scholar] [CrossRef]
- Moschetta, A.; Xu, F.; Hagey, L.R.; van Berge-Henegouwen, G.P.; van Erpecum, K.J.; Brouwers, J.F.; Cohen, J.C.; Bierman, M.; Hobbs, H.H.; Steinbach, J.H.; et al. A phylogenetic survey of biliary lipids in vertebrates. J. Lipid Res. 2005, 46, 2221–2232. [Google Scholar] [CrossRef]
- Hooper, L.V.; Midtvedt, T.; Gordon, J.I. How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu. Rev. Nutr. 2002, 22, 283–307. [Google Scholar] [CrossRef]
- Rhee, K.J.; Sethupathi, P.; Driks, A.; Lanning, D.K.; Knight, K.L. Role of commensal bacteria in development of gut-associated lymphoid tissues and preimmune antibody repertoire. J. Immunol. 2004, 172, 1118–1124. [Google Scholar] [CrossRef]
- Wexler, H.M. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria With Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef]
- Urdaneta, V.; Casadesús, J. Interactions between Bacteria and Bile Salts in the Gastrointestinal and Hepatobiliary Tracts. Front. Med. 2017, 4, 163. [Google Scholar] [CrossRef]
- Tang, W.; Yao, X.; Xia, F.; Yang, M.; Chen, Z.; Zhou, B.; Liu, Q. Modulation of the Gut Microbiota in Rats by Hugan Qingzhi Tablets during the Treatment of High-Fat-Diet-Induced Nonalcoholic Fatty Liver Disease. Oxid. Med. Cell. Longev. 2018, 2018, 7261619. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Wu, N.; Wang, X.; Chi, Y.; Zhang, Y.; Qiu, X.; Hu, Y.; Li, J.; Liu, Y. Dysbiosis gut microbiota associated with inflammation and impaired mucosal immune function in intestine of humans with non-alcoholic fatty liver disease. Sci. Rep. 2015, 5, 8096. [Google Scholar] [CrossRef] [PubMed]
- Torok, V.A.; Hughes, R.J.; Mikkelsen, L.L.; Perez-Maldonado, R.; Balding, K.; MacAlpine, R.; Percy, N.J.; Ophel-Keller, K. Identification and characterization of potential performance-related gut microbiotas in broiler chickens across various feeding trials. Appl. Environ. Microbiol. 2011, 77, 5868–5878. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Apte, U. Bile Acid Metabolism and Signaling in Cholestasis, Inflammation, and Cancer. Adv. Pharmacol. 2015, 74, 263–302. [Google Scholar] [CrossRef]
- Zhang, X.L.; Chen, L.; Yang, J.; Zhao, S.S.; Jin, S.; Ao, N.; Yang, J.; Liu, H.X.; Du, J. Vitamin D alleviates non-alcoholic fatty liver disease via restoring gut microbiota and metabolism. Front. Microbiol. 2023, 14, 1117644. [Google Scholar] [CrossRef] [PubMed]
- Milosavljevic, M.N.; Kostic, M.; Milovanovic, J.; Zaric, R.Z.; Stojadinovic, M.; Jankovic, S.M.; Stefanovic, S.M. Antimicrobial treatment of Erysipelatoclostridium ramosum invasive infections: A systematic review. Rev. Inst. Med. Trop. Sao Paulo 2021, 63, e30. [Google Scholar] [CrossRef] [PubMed]
- Zakham, F.; Pillonel, T.; Brunel, A.S.; Zambelli, P.Y.; Greub, G.; Croxatto, A.; Bertelli, C. Molecular diagnosis and enrichment culture identified a septic pseudoarthrosis due to an infection with Erysipelatoclostridium ramosum. Int. J. Infect. Dis. 2019, 81, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Wang, X.; Xu, G.; Yan, Q.; Huang, W. Bile acid signaling and liver regeneration. Biochim. Biophys. Acta 2015, 1849, 196–200. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient, % | Control | CLC |
|---|---|---|
| Corn Meal | 67.08 | 67.08 |
| Soybean Protein (36%) | 8.05 | 8.05 |
| Soybean Meal (44%) | 9.84 | 9.84 |
| Wheat Bran | 4.47 | 4.47 |
| Rice Bran | 4.415 | 2.465 |
| CaCO3 | 2.24 | 2.24 |
| CaHPO4 | 1.79 | 1.79 |
| Beef Tallow | 1.34 | 1.34 |
| NaCl | 0.36 | 0.36 |
| DL-Methionine | 0.36 | 0.36 |
| Premix 1 | 0.005 | 0.005 |
| Choline | 0.05 | - |
| Cholesterol | - | 2.00 |
| Total | 100 | 100 |
| Nutrient Composition, % | ||
| Crude Protein | 16.55 | 16.94 |
| Crude Fat | 4.72 | 4.72 |
| Methionine | 0.63 | 0.63 |
| Methionine + Cystine | 0.91 | 0.91 |
| Choline, mg/kg | 0.13 | 0.08 |
| ME, kcal/kg 2 | 3043 | 3043 |
| CON (n = 10) | CLC (n = 10) | |
|---|---|---|
| Primary Bile Acids (nM) | ||
| TCA | 122.3 ± 34.64 | 82.07 ± 17.26 |
| CDCA | 3.93 ± 1.52 | 38.13 ± 13.21 * |
| GCA | 0.58 ± 0.11 | 0.47 ± 0.08 |
| TCDCA | 6080 ± 1153 | 14,556 ± 1868 ** |
| isoDCA | 0.25 ± 0.16 | 3.21 ± 0.59 ** |
| T-α-MCA | 9.39 ± 2.21 | 16.23 ± 3.34 |
| Secondary Bile Acids (nM) | ||
| TLCA | 28.43 ± 7.77 | 2123 ± 481.8 *** |
| TUDCA | 0.42 ± 0.10 | 13.37 ± 3.91 * |
| THDCA | 19.35 ± 3.21 | 63.66 ± 6.99 *** |
| TDCA | 1.51 ± 0.24 | 35.06 ± 7.67 ** |
| 5β-Cholenic Acid-7α-ol-3-one | 1.00 ± 0.07 | 3.87 ± 0.41 *** |
| CON (n = 10) | CLC (n = 10) | |
|---|---|---|
| Primary Bile acid (nM) | ||
| CA | 315.4 ± 133 | 116.7 ± 38.26 |
| CDCA | 1289 ± 369 | 1151 ± 293 |
| GCDCA | 8.25 ± 2.26 | 21.35 ± 4.04 * |
| GCA | 1.72 ± 0.60 | 0.711 ± 0.10 |
| TCA | 10,300 ± 3118 | 12,302 ± 2208 |
| T-α-MCA | 447 ± 196.4 | 785 ± 169.6 |
| HCA | 1.197 ± 0.59 | 0.37 ± 0.10 |
| Secondary Bile acid (nM) | ||
| HDCA | 0.56 ± 0.23 | 1.717 ± 0.34 * |
| UDCA | 9.21 ± 2.57 | 11.88 ±1.96 |
| LCA | 4.32 ± 0.82 | 22.88 ± 6.85 ** |
| isoLCA | 3.48 ± 2.58 | 3.62 ± 1.51 |
| TLCA | 109.8 ± 27.36 | 8897 ± 1701 *** |
| TUDCA | 15.82 ± 6.19 | 985 ± 377 * |
| THDCA | 718.7 ± 219.2 | 3084 ± 502.9 *** |
| TDCA | 10.12 ± 2.65 | 172.3 ± 37.41 *** |
| THCA | 285.2 ± 76.32 | 131.8 ± 37.3 |
| 5β-Cholenic Acid-7α-ol-3-one | 13.47 ± 8.16 | 8.07 ± 3.64 |
| 6,7-diketoLCA | 1.54 ± 0.26 | 3.139 ± 0.40 * |
| 7-ketoLCA | 15.01 ± 6.69 | 11.81 ± 4.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.-Y.; Chang, P.-E.; Li, S.-J.; Ding, S.-T.; Lin, Y.-Y. Exploring Bile-Acid Changes and Microflora Profiles in Chicken Fatty Liver Disease Model. Animals 2024, 14, 992. https://doi.org/10.3390/ani14070992
Yang W-Y, Chang P-E, Li S-J, Ding S-T, Lin Y-Y. Exploring Bile-Acid Changes and Microflora Profiles in Chicken Fatty Liver Disease Model. Animals. 2024; 14(7):992. https://doi.org/10.3390/ani14070992
Chicago/Turabian StyleYang, Wen-Yuan, Pei-En Chang, Sin-Jin Li, Shih-Torng Ding, and Yuan-Yu Lin. 2024. "Exploring Bile-Acid Changes and Microflora Profiles in Chicken Fatty Liver Disease Model" Animals 14, no. 7: 992. https://doi.org/10.3390/ani14070992
APA StyleYang, W.-Y., Chang, P.-E., Li, S.-J., Ding, S.-T., & Lin, Y.-Y. (2024). Exploring Bile-Acid Changes and Microflora Profiles in Chicken Fatty Liver Disease Model. Animals, 14(7), 992. https://doi.org/10.3390/ani14070992