
A Comparative Analysis of the Venom System between Two Morphotypes of the Sea Anemone Actinia equina
 , , , ,
, , , ,  , ,
, ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Collection
2.2. Microscopy Analyses
2.3. Characterisation of the Venom Proteome
2.3.1. Venom Extraction
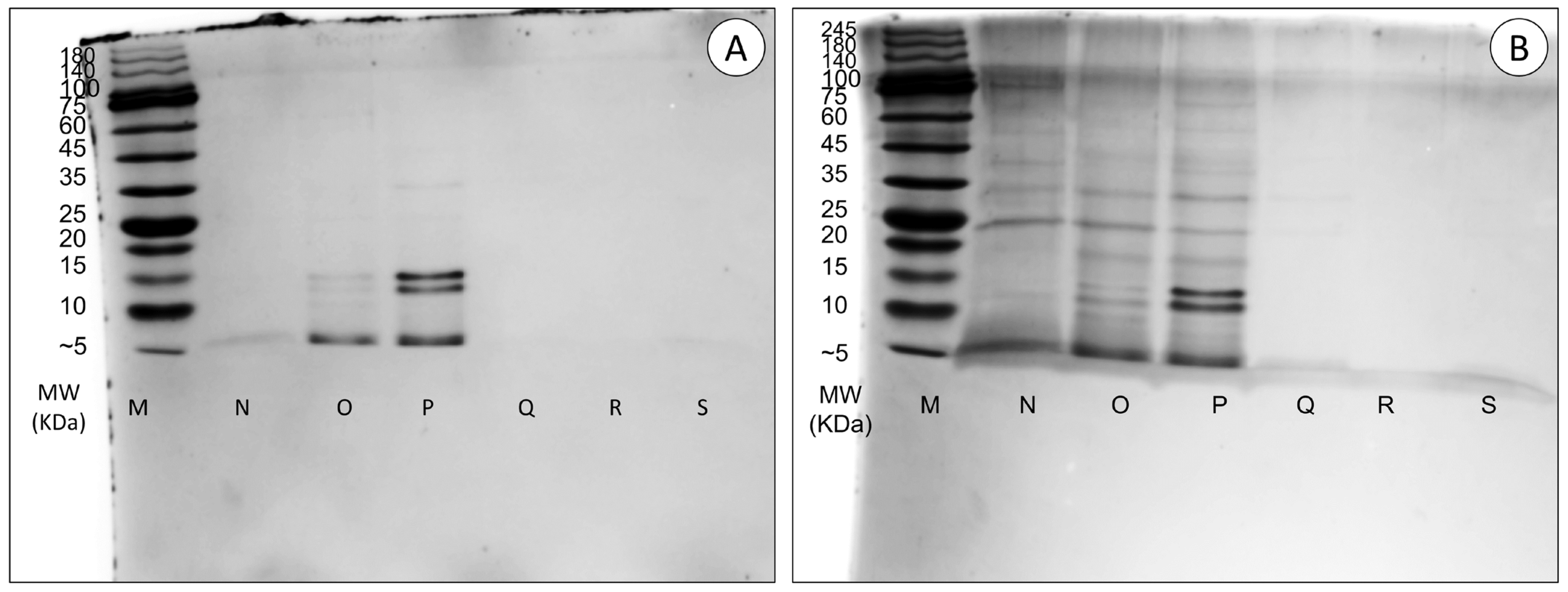
2.3.2. SDS-PAGE Analysis
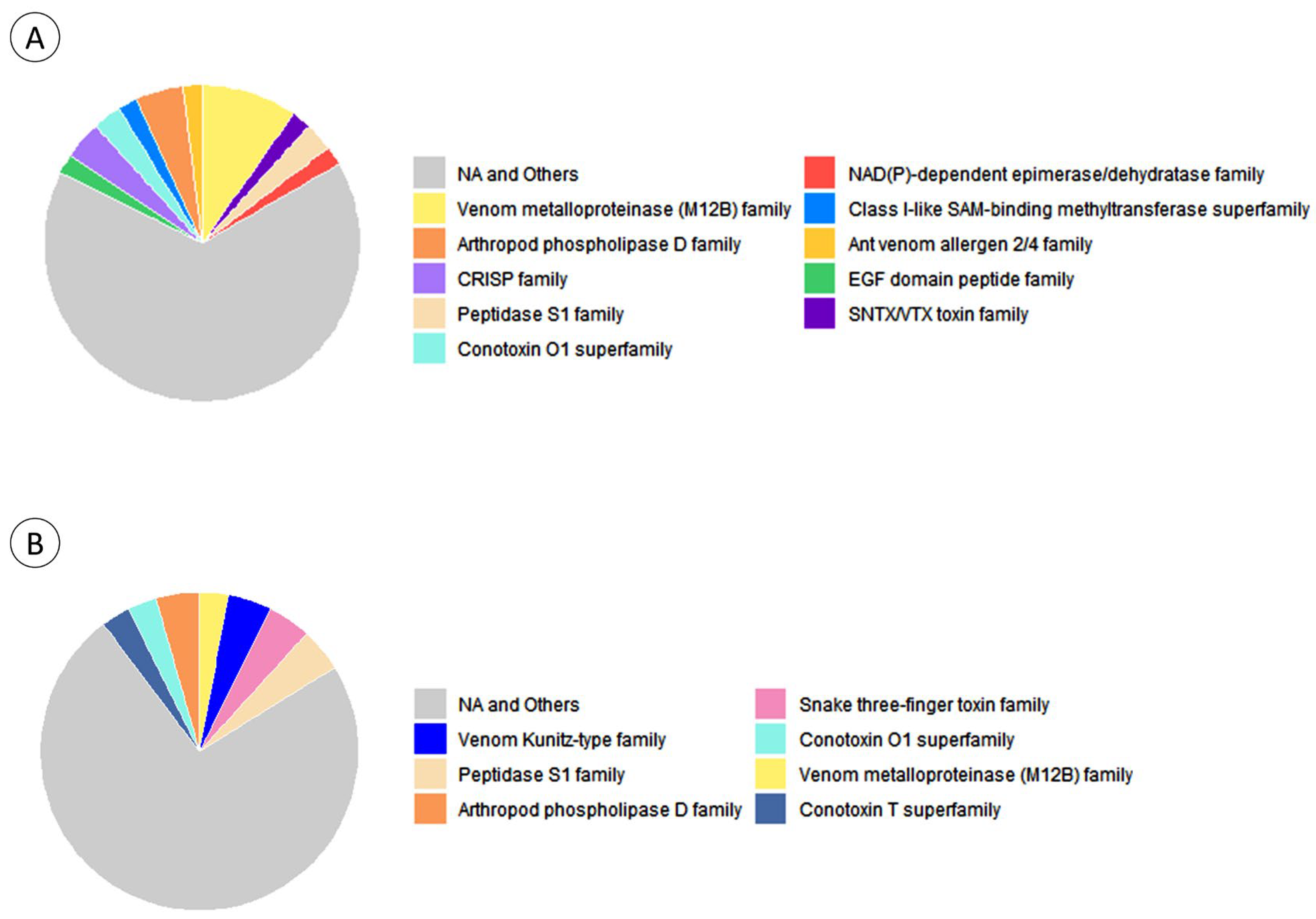
2.3.3. Toxin Identification by LC-MS/MS
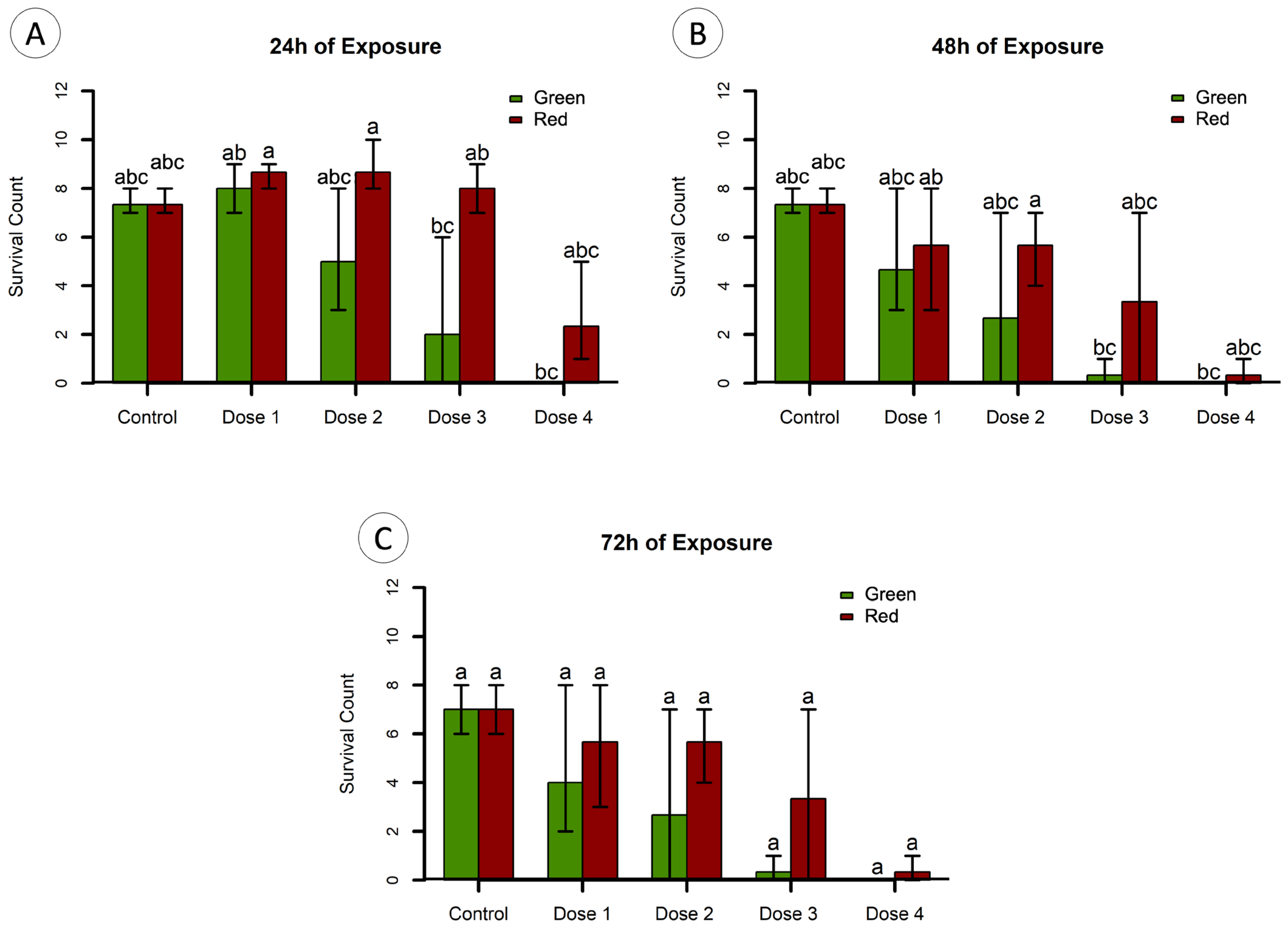
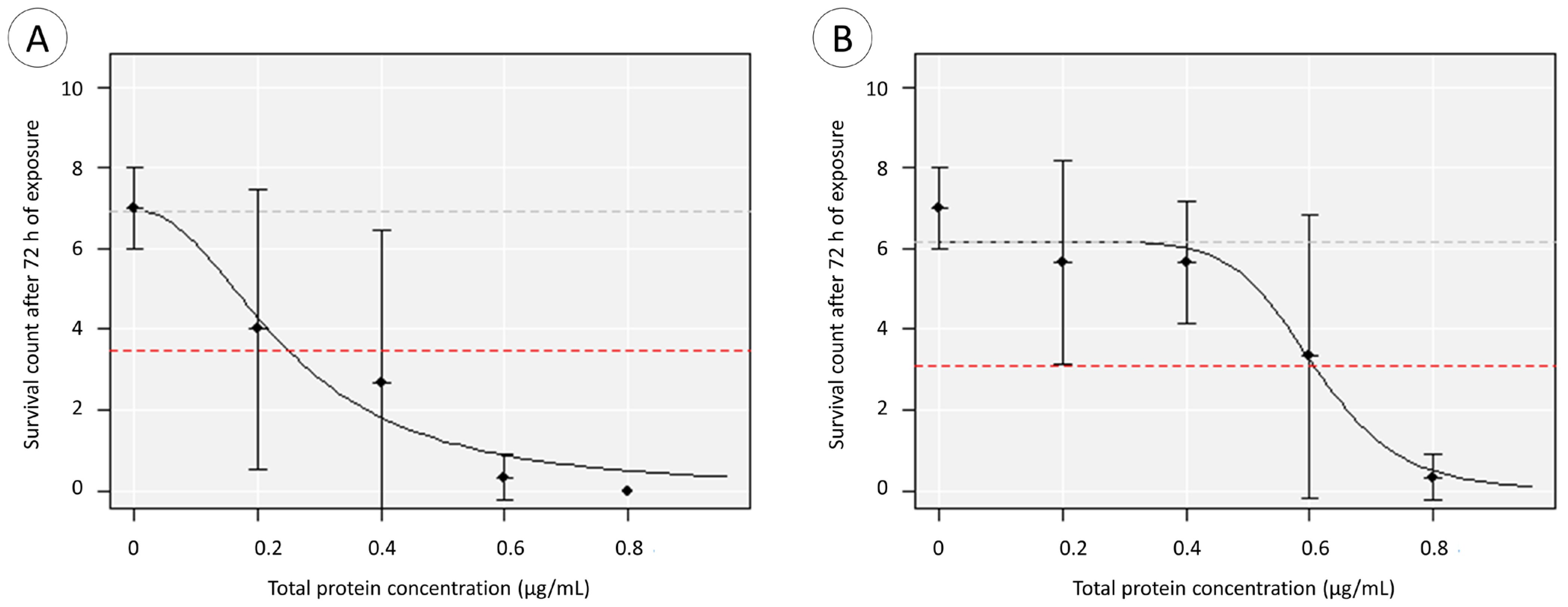
2.4. Toxicity Testing
2.5. Statistical Analysis
3. Results
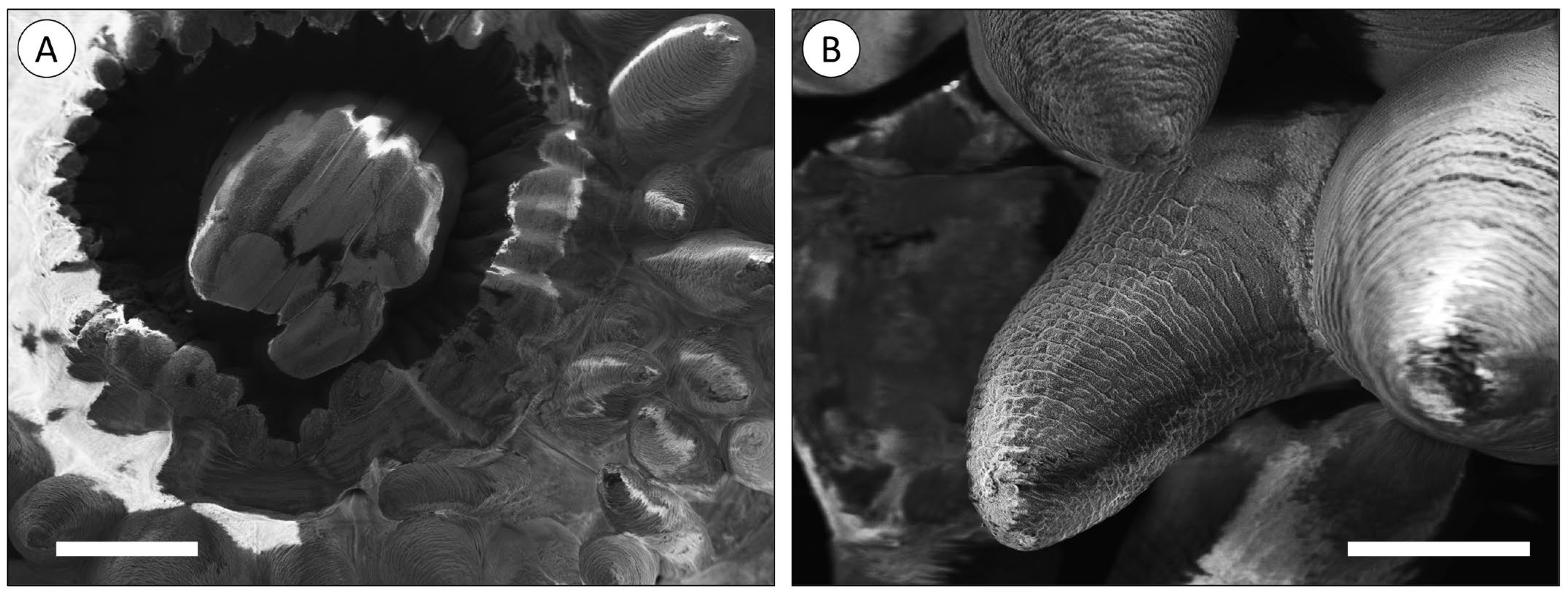
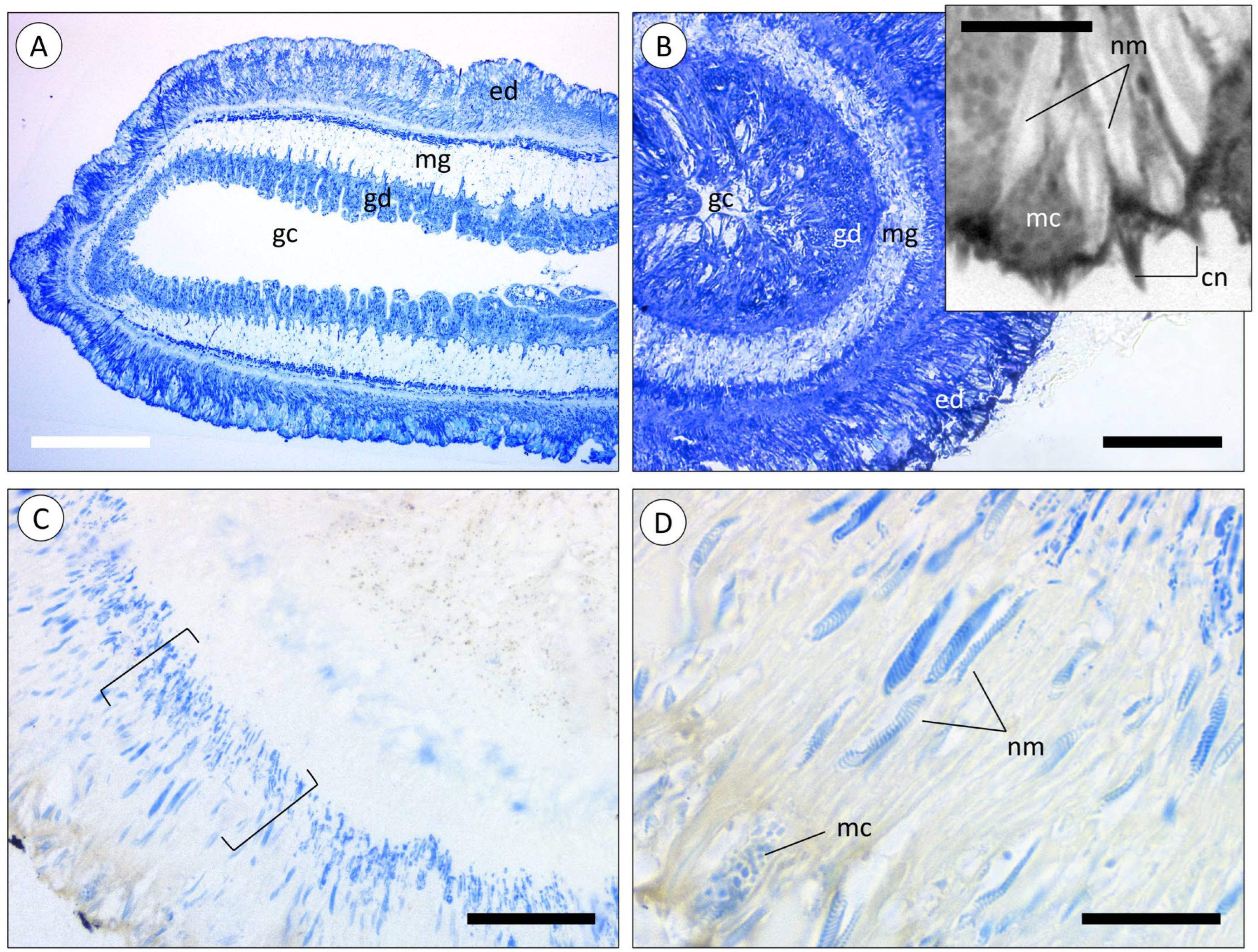
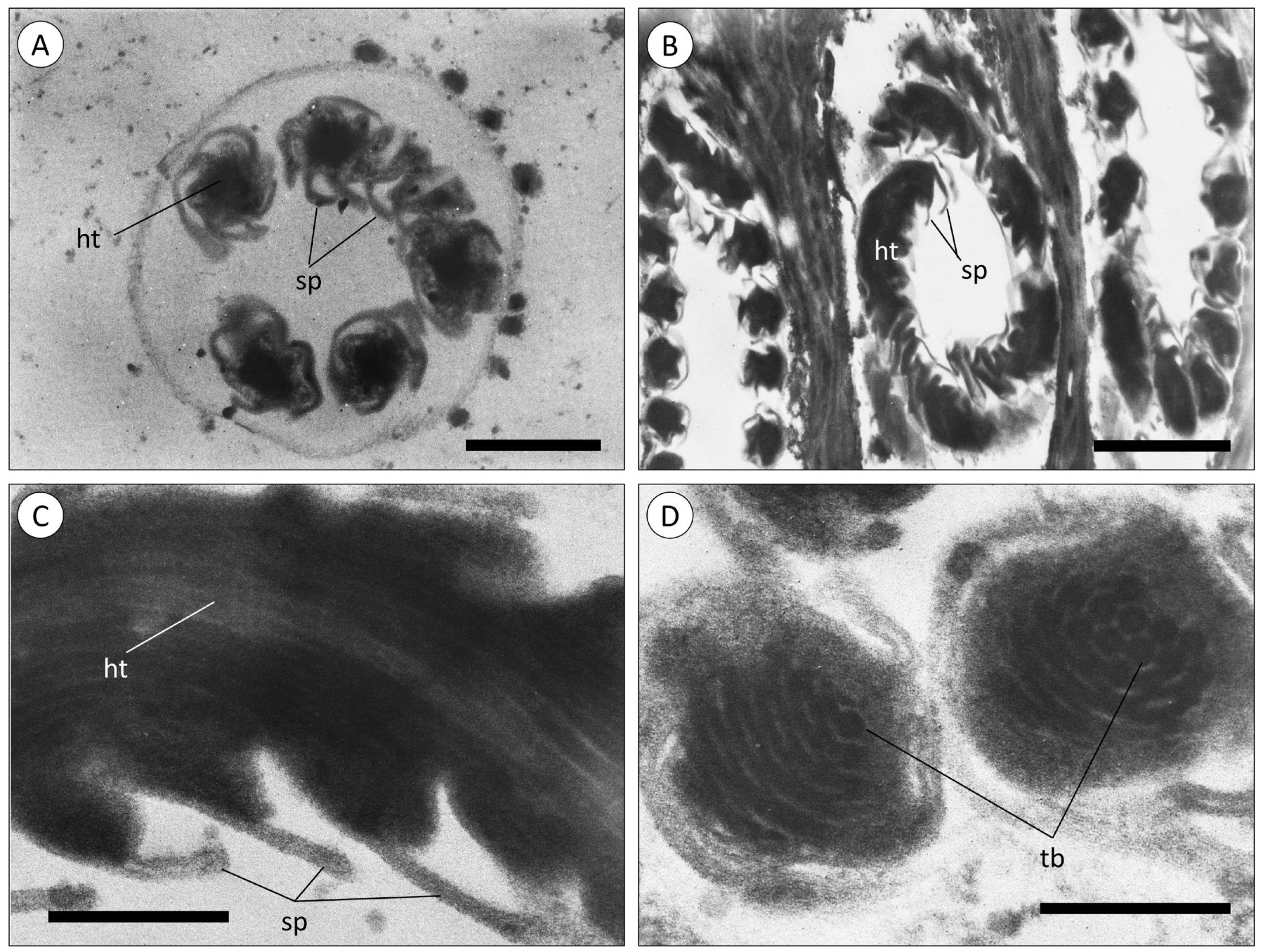
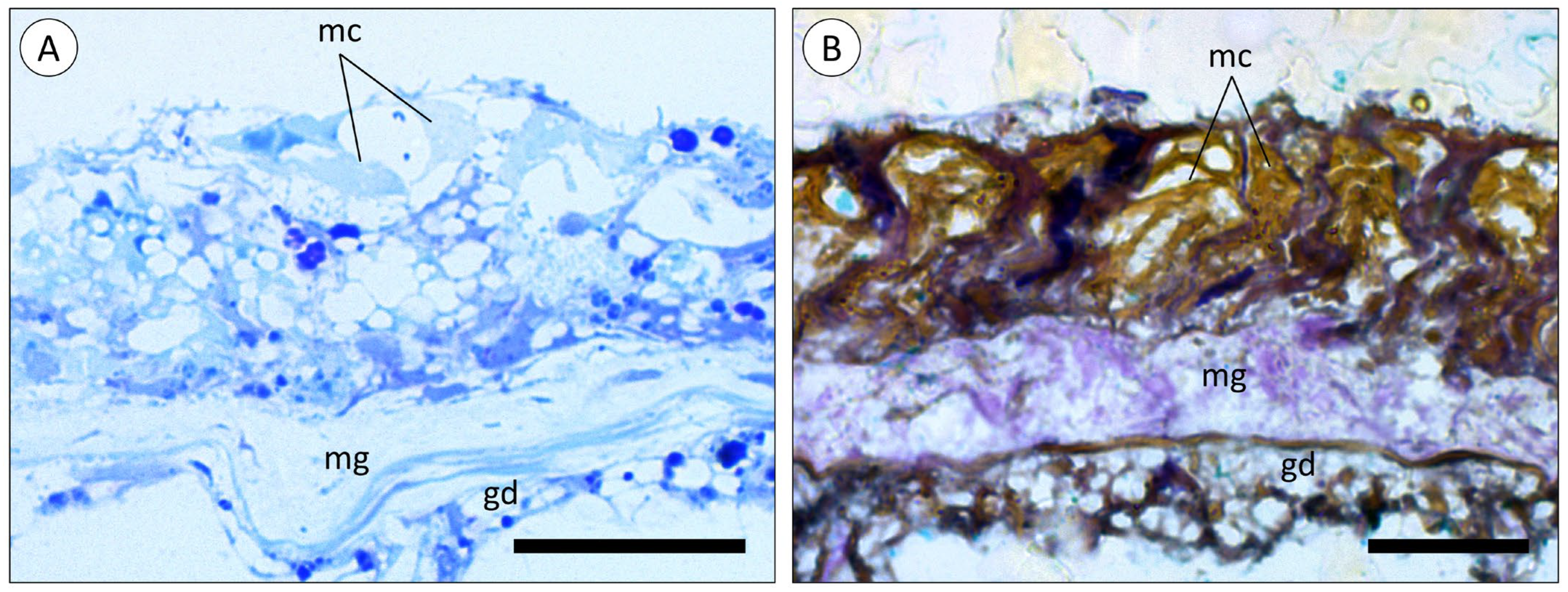
3.1. Morphoanatomic Characterisation of A. equina Venom-Delivery Apparatus
3.2. Toxicity Testing
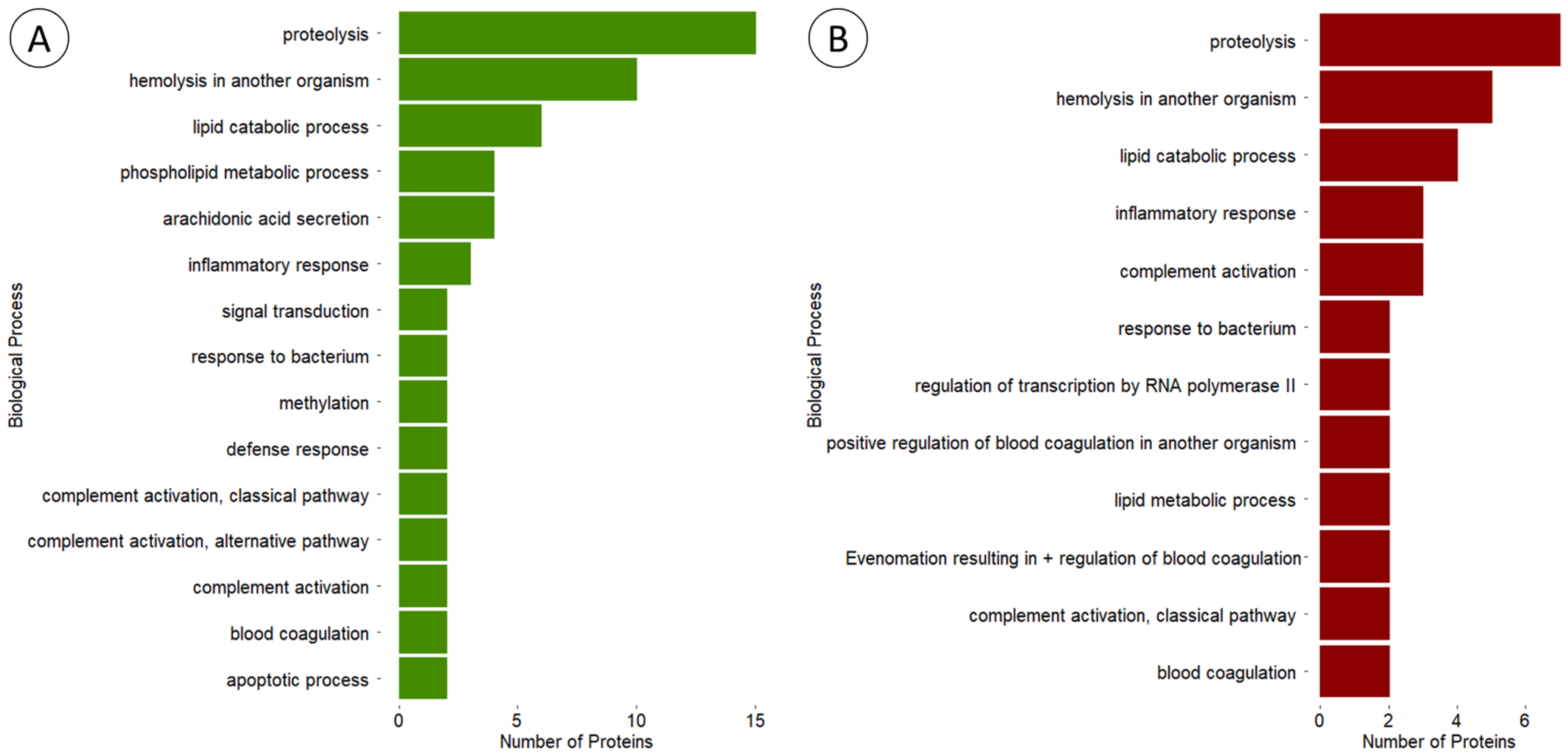
3.3. Characterisation of Venom Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindequist, U. Marine-Derived Pharmaceuticals—Challenges and Opportunities. Biomol. Ther. 2016, 24, 561–571. [Google Scholar] [CrossRef]
- Calvete, J.J.; Sanz, L.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M. Venoms, Venomics, Antivenomics. FEBS Lett. 2009, 583, 1736–1743. [Google Scholar] [CrossRef]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug Development from Marine Natural Products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar] [CrossRef]
- Williams, J.A.; Day, M.; Heavner, J.E. Ziconotide: An Update and Review. Expert. Opin. Pharmacother. 2008, 9, 1575–1583. [Google Scholar] [CrossRef]
- Christinat, A.; Leyvraz, S. Role of Trabectedin in the Treatment of Soft Tissue Sarcoma. OncoTargets Ther. 2009, 2, 105–113. [Google Scholar] [CrossRef][Green Version]
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient Venom Systems: A Review on Cnidaria Toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef] [PubMed]
- Rocha, J.; Peixe, L.; Gomes, N.C.M.; Calado, R. Cnidarians as a Source of New Marine Bioactive Compounds—An Overview of the Last Decade and Future Steps for Bioprospecting. Mar. Drugs 2011, 9, 1860–1886. [Google Scholar] [CrossRef]
- Sher, D.; Fishman, Y.; Zhang, M.; Lebendiker, M.; Gaathon, A.; Mancheño, J.M.; Zlotkin, E. Hydralysins, a New Category of β-Pore-Forming Toxins in Cnidaria. J. Biol. Chem. 2005, 280, 22847–22855. [Google Scholar] [CrossRef]
- D’Ambra, I.; Lauritano, C. A Review of Toxins from Cnidaria. Mar. Drugs 2020, 18, 507. [Google Scholar] [CrossRef] [PubMed]
- Tarcha, E.J.; Olsen, C.M.; Probst, P.; Peckham, D.; Muñoz-Elías, E.J.; Kruger, J.G.; Iadonato, S.P. Safety and Pharmacodynamics of Dalazatide, a Kv1.3 Channel Inhibitor, in the Treatment of Plaque Psoriasis: A Randomized Phase 1b Trial. PLoS ONE 2017, 12, e0180762. [Google Scholar] [CrossRef]
- Beeton, C.; Pennington, M.W.; Wulff, H.; Singh, S.; Nugent, D.; Crossley, G.; Khaytin, I.; Calabresi, P.A.; Chen, C.Y.; Gutman, G.A.; et al. Targeting Effector Memory T Cells with a Selective Peptide Inhibitor of Kv1.3 Channels for Therapy of Autoimmune Diseases. Mol. Pharmacol. 2005, 67, 1369–1381. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Eckel-Mahan, K.L.; Mirbolooki, M.R.; Tjong, I.; Griffey, S.M.; Schmunk, G.; Koehne, A.; Halbout, B.; Iadonato, S.; Pedersen, B.; et al. Selective Kv1.3 Channel Blocker as Therapeutic for Obesity and Insulin Resistance. Proc. Natl. Acad. Sci. USA 2013, 110, E2239–E2248. [Google Scholar] [CrossRef]
- Frazão, B.; Vasconcelos, V.; Antunes, A. Sea Anemone (Cnidaria, Anthozoa, Actiniaria) Toxins: An Overview. Mar. Drugs 2012, 10, 1812–1851. [Google Scholar] [CrossRef]
- Fish, J.D.; Fish, S. Cnidaria. In A Student’s Guide to the Seashore; Cambridge University Press: Cambridge, UK, 2011; pp. 86–125. [Google Scholar]
- Mariscal, R.N. Cnidaria: Cnidae. In Biology of the Integument; Springer: Berlin/Heidelberg, Germany, 1984; pp. 57–68. [Google Scholar] [CrossRef]
- Turk, T.; Kem, W.R. The Phylum Cnidaria and Investigations of its Toxins and Venoms until 1990. Toxicon 2009, 54, 1031–1037. [Google Scholar] [CrossRef]
- Shick, J.M. Overview of Sea Anemones. In A Functional Biology of Sea Anemones; Calow, P., Ed.; Springer: Dordrecht, The Netherlands, 1991; pp. 1–35. [Google Scholar] [CrossRef]
- Menezes, C.; Thakur, N.L. Sea Anemone Venom: Ecological Interactions and Bioactive Potential. Toxicon 2022, 208, 31–46. [Google Scholar] [CrossRef]
- Solé-Cava, A.M.; Thorpe, J.P. Genetic Divergence between Colour Morphs in Populations of the Common Intertidal Sea Anemones Actinia equina and A. prasina (Anthozoa: Actiniaria) in the Isle of Man. Mar. Biol. 1992, 112, 243–252. [Google Scholar] [CrossRef]
- Pereira, A.M.; Brito, C.; Sanches, J.; Sousa-Santos, C.; Robalo, J.I. Absence of Consistent Genetic Differentiation among Several Morphs of Actinia (Actiniaria: Actiniidae) Occurring in the Portuguese Coast. Zootaxa 2014, 3893, 595–600. [Google Scholar] [CrossRef]
- Costa, P.M. The Handbook of Histopathological Practices in Aquatic Environments: Guide to Histology for Environmental Toxicology; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Venable, J.H.; Coggeshall, R. A Simplified Lead Citrate Stain for Use in Electron Microscopy. J. Cell Biol. 1965, 25, 407–408. [Google Scholar] [CrossRef] [PubMed]
- Inoué, T.; Osatake, H. A New Drying Method of Biological Specimens for Scanning Electron Microscopy: The T-Butyl Alcohol Freeze-Drying Method. Arch. Histol. Cytol. 1988, 51, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Macek, P.; Lebez, D. Isolation and Characterization of Three Lethal and Hemolytic Toxins from the Sea Anemone Actinia equina L. Toxicon 1988, 26, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins During the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Rodrigo, A.P.; Lopes, A.; Pereira, R.; Anjo, S.I.; Manadas, B.; Grosso, A.R.; Baptista, P.V.; Fernandes, A.R.; Costa, P.M. Endogenous Fluorescent Proteins in the Mucus of an Intertidal Polychaeta: Clues for Biotechnology. Mar. Drugs 2022, 20, 224. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Metand for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ihaka, R.; Gentleman, R. R: A Language for Data Analysis and Graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar] [CrossRef]
- Chintiroglou, C.; Koukouras, A. The Feeding Habits of Three Mediterranean Sea Anemone Species, Anemonia viridis (Forskål), Actinia equina (Linnaeus) and Cereus pedunculatus (Pennant). Helgoländer Meeresunters. 1992, 46, 53–68. [Google Scholar] [CrossRef]
- Stabili, L.; Schirosi, R.; Parisi, M.; Piraino, S.; Cammarata, M. The Mucus of Actinia equina (Anthozoa, Cnidaria): An Unexplored Resource for Potential Applicative Purposes. Mar. Drugs 2015, 13, 5276–5296. [Google Scholar] [CrossRef]
- Sahyoun, C.; Krezel, W.; Mattei, C.; Sabatier, J.-M.; Legros, C.; Fajloun, Z.; Rima, M. Neuro- and cardiovascular activities of Montivipera bornmuelleri Snake Venom. Biology 2022, 11, 888. [Google Scholar] [CrossRef]
- Bordon, K.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar] [CrossRef] [PubMed]
- Amreen Nisa, S.; Vinu, D.; Krupakar, P.; Govindaraju, K.; Sharma, D.; Vivek, R. Jellyfish Venom Proteins and their Pharmacological Potentials: A Review. Int. J. Biol. Macromol. 2021, 176, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.Y.; Ishida, M.; Nagashima, Y.; Shiomi, K. A Polypeptide Toxin in The Sea Anemone Actinia equina Homologous with Other Sea Anemone Sodium Channel Toxins: Isolation and Amino Acid Sequence. Toxicon 1996, 34, 57–65. [Google Scholar] [CrossRef]
- Oshiro, N.; Kobayashi, C.; Iwanaga, S.; Nozaki, M.; Namikoshi, M.; Spring, J.; Nagai, H. A New Membrane-Attack Complex/Perforin (MACPF) Domain Lethal Toxin from the Nematocyst Venom of the Okinawan Sea Anemone Actineria villosa. Toxicon 2004, 43, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Oshiro, N.; Takuwa-Kuroda, K.; Iwanaga, S.; Nozaki, M.; Nakajima, T. Novel Proteinaceous Toxins from the Nematocyst Venom of the Okinawan Sea Anemone Phyllodiscus semoni kwietniewski. Biochem. Biophys. Res. Commun. 2002, 294, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Ferlan, I.; Lebez, D. Equinatoxin, A Lethal Protein from Actinia equina—I Purification and Characterization. Toxicon 1974, 12, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Carreto, S.; Miranda-Zaragoza, B.; Rodríguez-Almazán, C. Actinoporins: From the Structure and Function to the Generation of Biotechnological and Therapeutic Tools. Biomolecules 2020, 10, 539. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Accession | Species | Number of Peptides | % Coverage | ||
|---|---|---|---|---|---|---|
| Red | Green | Red | Green | |||
| Delta-actitoxin-Aeq2a | Q9NJQ2 | Actinia equina | 9 | 11 | 63.41 | 65.85 |
| PI-actitoxin-Aeq3c | P0DMJ2 | Actinia equina | 8 | 6 | 87.72 | 87.72 |
| PI-actitoxin-Aeq3a | P0DMW6 | Actinia equina | 5 | 5 | 77.97 | 77.97 |
| U-actitoxin-Avd3n-like | A0A6P8IZ51 | Actinia tenebrosa | 1 | 2 | 12.5 | 12.5 |
| PI-actitoxin-Aeq3a-like | A0A6P8HBL9 | Actinia tenebrosa | 4 | 4 | 62.85 | 62.82 |
| U-actitoxin-Bgr3d | G0W2I1 | Bunodosoma granuliferum | 1 | 1 | 11.39 | 11.39 |
| Protein Name | Accession | Species | Number of Peptides | % Coverage |
|---|---|---|---|---|
| Delta-actitoxin-Aeq2b | B1NWU4 | Actinia equina | 3 | 47.56 |
| Delta-alicitoxin-Pse2a | P58911 | Phyllodiscus semoni | 1 | 1.80 |
| Delta-alicitoxin-Pse2b | P58912 | Phyllodiscus semoni | 1 | 3.07 |
| U-actitoxin-Avd9c | P0DN02 | Anemonia viridis | 1 | 21.25 |
| Potassium channel toxin alpha-KTx 6.2 | P80719 | Scorpius palmatus | 1 | 35.29 |
| U16-lycotoxin-Ls1a | B6DD52 | Lycosa singoriensis | 1 | 8.54 |
| Protein Name | Accession | Species | Number of Peptides | % Coverage |
|---|---|---|---|---|
| U-actitoxin-Avd3s (Fragment) | P0DN20 | Anemonia viridis | 4 | 41.25 |
| Delta-actitoxin-Aeq2b 2 | B1NWU3 | Actinia equina | 3 | 45.12 |
| KappaPI-actitoxin-Ael3a | P86862 | Anthopleura elegantissima | 2 | 13.85 |
| Type III potassium channel toxin protein | A0A0S1M193 | Anemonia sulcata | 2 | 18.67 |
| Delta-thalatoxin-Avl2a | Q76DT2 | Actineria villosa | 2 | 5.42 |
| Phospholipase A2 A2-actitoxin-Ucs2a | A7LCJ2 | Urticina crassicornis | 1 | 6.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alcaide, M.; Moutinho Cabral, I.; Carvalho, L.; Mendes, V.M.; Alves de Matos, A.P.; Manadas, B.; Saúde, L.; D’Ambrosio, M.; Costa, P.M. A Comparative Analysis of the Venom System between Two Morphotypes of the Sea Anemone Actinia equina. Animals 2024, 14, 981. https://doi.org/10.3390/ani14060981
Alcaide M, Moutinho Cabral I, Carvalho L, Mendes VM, Alves de Matos AP, Manadas B, Saúde L, D’Ambrosio M, Costa PM. A Comparative Analysis of the Venom System between Two Morphotypes of the Sea Anemone Actinia equina. Animals. 2024; 14(6):981. https://doi.org/10.3390/ani14060981
Chicago/Turabian StyleAlcaide, Maria, Inês Moutinho Cabral, Lara Carvalho, Vera M. Mendes, António P. Alves de Matos, Bruno Manadas, Leonor Saúde, Mariaelena D’Ambrosio, and Pedro M. Costa. 2024. "A Comparative Analysis of the Venom System between Two Morphotypes of the Sea Anemone Actinia equina" Animals 14, no. 6: 981. https://doi.org/10.3390/ani14060981
APA StyleAlcaide, M., Moutinho Cabral, I., Carvalho, L., Mendes, V. M., Alves de Matos, A. P., Manadas, B., Saúde, L., D’Ambrosio, M., & Costa, P. M. (2024). A Comparative Analysis of the Venom System between Two Morphotypes of the Sea Anemone Actinia equina. Animals, 14(6), 981. https://doi.org/10.3390/ani14060981