
Impact of Body Reserves Dynamic on Productivity and Reproductive Performance in Fat-Tail and Thin-Tail Sheep Breeds over Contrasting Production Cycles

 , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Material and Methods
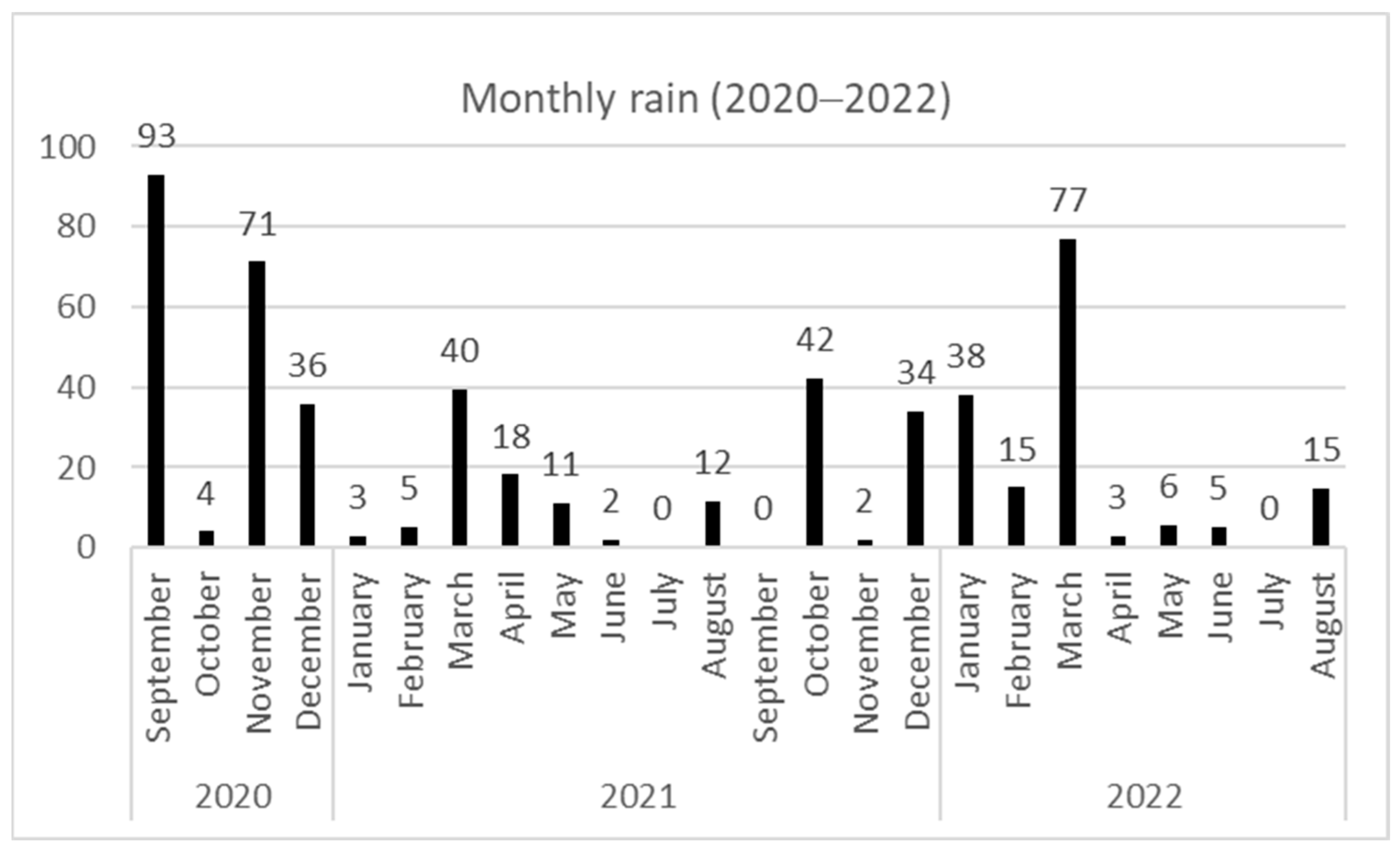
2.1. Experimental Location, Animals, and General Management
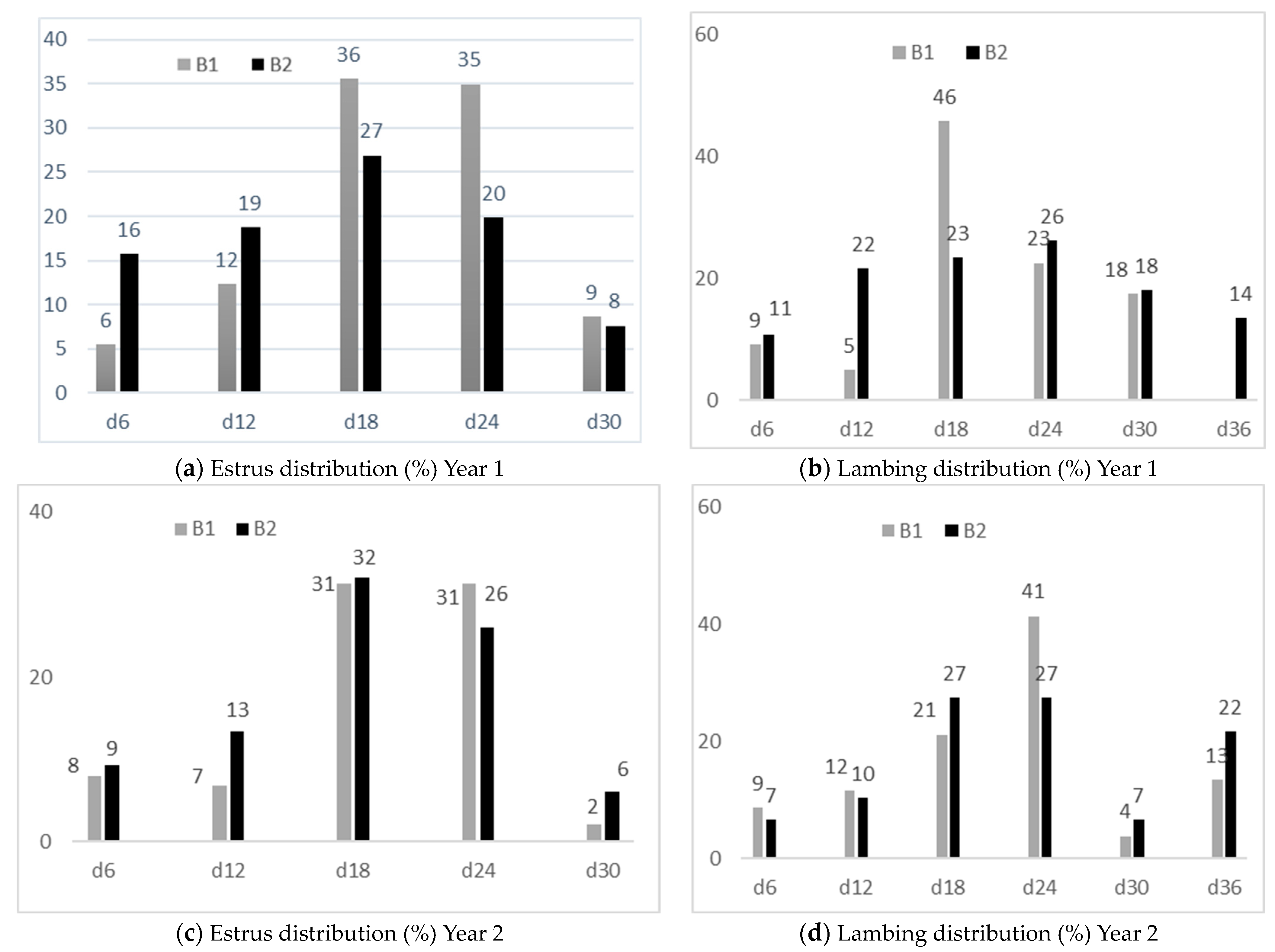
2.2. Reproductive Management
2.3. Measurements on Animals
2.4. Reproduction and Growth Parameters Calculation
2.5. Statistical Analyses
3. Results and Discussion
3.1. Body Weight and Body Condition Score Evolution during Two Consecutive Production Cycles
3.2. Reproductive Performances of Ewes
3.3. Offspring Growth Performances
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atti, N.; Thériez, M.; Abdennebi, L. Relationship between ewe body condition at mating and reproductive performance in the fat-tailed Barbarine breed. Anim. Res. 2001, 50, 135–144. [Google Scholar] [CrossRef]
- Friggens, N.C.; Blanc, F.; Berry, D.P.; Puillet, L. Review: Deciphering animal robustness. A synthesis to facilitate its use in livestock breeding and management. Animal 2017, 11, 2237–2251. [Google Scholar] [CrossRef] [PubMed]
- Belanche, A.; Martin-Collado, D.; Rose, G.; Yanez-Ruiz, D.R. A multi-stakeholder participatory study identifies the priorities for the sustainability of the small ruminants farming sector in Europe. Animal 2021, 15, 100131. [Google Scholar] [CrossRef]
- Macé, T.; González-García, E.; Carrière, F.; Douls, S.; Foulquié, D.; Robert-Granié, C.; Hazard, D. Intra-flock variability in the body reserve dynamics of meat sheep by analyzing BW and body condition score variations over multiple production cycles. Animal 2019, 13, 1986–1998. [Google Scholar] [CrossRef]
- Macé, T.; Hazard, D.; Carrière, F.; Douls, S.; Foulquié, D.; González-García, E. Relationships between body reserve dynamics and rearing performances in meat ewes. J. Anim. Sci. 2019, 97, 4076–4084. [Google Scholar] [CrossRef]
- Sanchez-Molano, E.; Kapsona, V.V.; Oikonomou, S.; McLaren, A.; Lambe, N.; Conington, J.; Banos, G. Breeding strategies for animal resilience to weather variation in meat sheep. BMC Gen. 2020, 21, 116–126. [Google Scholar] [CrossRef]
- Friggens, N.C.; Brun-Lafleur, L.; Faverdin, P.; Sauvant, D.; Martin, O. Advances in predicting nutrient partitioning in the dairy cow: Recognizing the central role of genotype and its expression through time. Animal 2013, 7, 89–101. [Google Scholar] [CrossRef] [PubMed]
- McLaren, A.; Lambe, N.R.; Conington, J. Genetic associations of ewe body condition score and lamb rearing performance in extensively managed meat sheep. Livest. Sci. 2023, 277, 105336. [Google Scholar] [CrossRef]
- Bergho, T.V.L.; Poppe, M.; Mulder, H.A. Opportunities to improve resilience in animal breeding programmes. Front. Genet. 2019, 9, 692. [Google Scholar] [CrossRef]
- Magsi, S.H.; Rashid, M.A.; Ahamed, N.; Akhter, M.; Shahid, M.Q. Effect of body condition score at calving on transition success in Nili Ravi buffaloes. J. Anim. Sci. Techn. 2022, 64, 1013–1023. [Google Scholar] [CrossRef]
- Harder, I.; Stamer, E.; Junge, W.; Thaller, G. Lactation curves and model evaluation for feed intake and energy balance in dairy cows. J. Dairy Sci. 2019, 102, 7204–7216. [Google Scholar] [CrossRef] [PubMed]
- Buonaiuto, G.; Lopez-Villalobos, N.; Niero, G.; Degano, L.; Dadati, E.; Formigoni, A.; Visentin, G. The application of Legendre Polynomials to model muscularity and body condition score in primiparous Italian Simmental cattle. Ital. J. Anim. Sci. 2022, 21, 350–360. [Google Scholar] [CrossRef]
- Atti, N.; Bocquier, F. Adaptation des brebis Barbarine à l’alternance sous-nutrition- réalimentation: Effets sur les tissus adipeux. Ann. Zootech. 1999, 48, 189–198. [Google Scholar] [CrossRef]
- Chilliard, Y.; Bocquier, F.; Doreau, M. Digestive and metabolic adaptations of ruminants to undernutrition, and consequences on reproduction. Reprod. Nutr. Dev. 1998, 38, 131–152. [Google Scholar] [CrossRef] [PubMed]
- Lambe, N.R.; Brotherstone, S.; Young, M.J.; Conington, J.; Simm, G. Genetic relationships between seasonal tissue levels in Scottish Blackface ewes and lamb growth traits. Anim. Sci. 2005, 81, 11–21. [Google Scholar] [CrossRef]
- González-García, E.; Gozzo de Figuereido, V.; Foulquie, D.; Jousserand, E.; Autran, P.; Camous, S.; Tesniere, A.; Bocquier, F.; Jouven, M. Circannual body reserve dynamics and metabolic profile changes in Romane ewes grazing on rangelands. Domest. Anim. Endoc. 2014, 46, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Ndlovu, T.; Chimonyo, M.; Okoh, A.I.; Muchenje, V.; Dzama, K.; Raats, J.G. Assessing the nutritional status of beef cattle: Current practices and future prospects. Afr. J. Biotechnol. 2007, 6, 2727–2734. [Google Scholar] [CrossRef]
- Vatankhah, M.; Talebi, M.A.; Zamani, F. Relationship between ewe body condition score (BCS) at mating and reproductive and productive traits in Lori-Bakhtiari sheep. Small Rum. Res. 2012, 106, 105–109. [Google Scholar] [CrossRef]
- Russel, A.J.F.; Doney, J.M.; Gunn, R.G. Subjective assessment of body fat in live sheep. J. Agric. Sci. 1969, 72, 451–454. [Google Scholar] [CrossRef]
- Aliyari, D.; Moeini, M.M.; Shahir, M.H.; Sirjani, M.A. Effect of body condition score, live weight and age on reproductive performance of Afshari ewes. Asian J. Anim. Vet. Adv. 2012, 7, 904–909. [Google Scholar] [CrossRef]
- Bocquier, F.; Thériez, M.; Prache, S.; Brelurut, A. Alimentation des ovins. In Alimentation des Bovins, des Ovins et des Caprins; Jarrige, R., Ed.; INRA Editions: Paris, France, 1988; pp. 249–281. Available online: http://www.civamad53.org/wp-content/uploads/2020/12/Tables-INRA.pdf (accessed on 18 September 2019).
- Atti, N.; Bocquier, F. Estimation in vivo des réserves corporelles de la brebis à queue grasse à partir des notes d’état corporel et des mensurations de la queue. Ann. INRAT 2007, 80, 137–154. [Google Scholar]
- SAS. Statistical Analysis System, User’s Guide. In Statistical, 7th ed.; SAS Inst. Inc.: Cary, NC, USA, 2004; pp. 25–421. [Google Scholar]
- Paganoni, B.L.; Ferguson, M.B.; Greeff, C.J.M.; Kearney, G.A.; Thompson, A.N. Mating weight and condition score are both good predictors of lambing potential for young Merino and Border Leicester Merino ewes across different environments and years in Australia. Anim. Prod. Sci. 2022, 62, 1507–1517. [Google Scholar] [CrossRef]
- Yagoubi, Y.; Atti, N. Effects of the fat-tailed ewes’ body condition scores at lambing on their metabolic profile and offspring growth. Arch. Anim. Breed. 2020, 63, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Macé, T.; González-García, E.; Pradel, J.; Parisot, S.; Carrière, F.; Douls, S.; Foulquié, D.; Hazard, D. Genetic analysis of robustness in meat sheep through body weight and body condition score changes over time. J. Anim. Sci. 2018, 96, 4501–4511. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.M.; Friggens, N.C.; Løvendahl, P.; Jensen, J.; Ingvartsen, K.L. Influence of breed, parity, and stage of lactation on lactational performance and relationship between body fatness and live weight. Livest. Prod. Sci. 2003, 79, 119–133. [Google Scholar] [CrossRef]
- Yagoubi, Y.; Atti, N. Effets de l’ablation de la queue sur les performances de reproduction des brebis de race Barbarine. J. New Sci. Agric. Biotech. 2018, 59, 3806–3815. [Google Scholar]
- Kharrat, M.; Bocquier, F. Adaptive responses at the whole lactation scale of Baladi dairy goats according to feed supply and level of body reserves in agro-pastoral feeding system. Small Rumin. Res. 2010, 90, 120–126. [Google Scholar] [CrossRef]
- Beker, A.; Gipson, T.A.; Puchala, R.; Askar, A.R.; Tesfai, K.; Detweiler, G.D.; Asmare, A.; Goestsch, A.L. Effects of stocking rate, breed and stage of production on energy expenditure and activity of meat goat does on pasture. J. Appl. Anim. Res. 2009, 36, 159–174. [Google Scholar] [CrossRef]
- Rekik, M.; Ben Salem, I.; Ben Hamouda, M.; Aloulou, R.; Ben Sassi, M. Sources of variation and genetic analysis of fertility of maiden ewes in an out-of-season management system. Livest. Res. Rural. Dev. 2011, 23, 18. [Google Scholar]
- Nechifor, I.; Florea, M.A.; Radu-Rusu, R.M.; Pascal, C. Influence of supplemental feeding on body condition score and reproductive performance dynamics in Botosani Karakul sheep. Agriculture 2022, 12, 2006. [Google Scholar] [CrossRef]
- Corner-Thomas, R.A.; Hickson, R.E.; Morris, S.T.; Back, P.J.; Ridler, A.L.; Stafford, K.J.; Kenyon, P.R. Effects of body condition score and nutrition in lactation on twin-bearing ewe and lamb performance to weaning. N. Z. J. Agric. Res. 2015, 58, 156–169. [Google Scholar] [CrossRef]
- Yilmaz, M.; Altin, T.; Karaca, O.; Cemal, I.; Bardakcioglu, H.E.; Yilmaz, O.; Taskin, T. Effect of body condition score at mating on the reproductive performance of Kivircik sheep under an extensive production system. Trop. Anim. Health Prod. 2011, 43, 1555–1560. [Google Scholar] [CrossRef] [PubMed]
- Walkom, S.F.; Brown, D.J. Genetic evaluation of adult ewe body weight and condition: Relationship with lamb growth, reproduction, carcass and wool production. Anim. Prod. Sci. 2017, 57, 20–32. [Google Scholar] [CrossRef]
- Thimonier, J.; Cognie, Y.; Lassoued, N.; Khaldi, G. L’effet mâle chez les ovins: Une technique actuelle de maîtrise de la reproduction. INRAE Prod. Anim. 2000, 13, 223–231. [Google Scholar] [CrossRef]
- Parraguez, V.H.; Sales, F.; Peralta, O.A.; Narbona, E.; Lira, R.; De los Reyes, M.; González-Bulnes, A. Supplementation of underfed twin-bearing ewes with herbal vitamins C and E: Impacts on birth weight, postnatal growth, and pre-weaning survival of the lambs. Animals 2020, 10, 652. [Google Scholar] [CrossRef] [PubMed]
- Koyuncu, M. Reproductive Performance of Kivircik Ewes on Accelerated Lambing Management. Pakist. J. Bio. Sci. 2005, 8, 1499–1502. [Google Scholar] [CrossRef]
- Ben Khlil, Z.; Khnissi, S.; Rekik, M.; Lassoued, N. Feed supplementation improves estrus response and increases fertility of sheep induced to breed out of season. Trop. Anim. Health Prod. 2017, 49, 607–612. [Google Scholar] [CrossRef]
- Rekik, M.; Ben Salem, I.; Diallo, H.; Ammar, H.; Aloulou, R. Productivité numérique et pondérale des brebis produites du croisement entre la D’man et la race locale Queue fine de l’Ouest. Rev. Elev. Méd. Vét. Pays Trop. 2005, 58, 81–88. [Google Scholar] [CrossRef][Green Version]
- Gardner, D.S.; Buttery, P.J.; Daniel, Z.; Symonds, M.E. Factors affecting birth weight in sheep: Maternal Environ. Reproduction 2007, 133, 297–307. [Google Scholar] [CrossRef]
- Karakuş, F.; Atmaca, M. The effect of ewe body condition at lambing on growth of lambs and colostral specific gravity. Arch. Anim. Breed. 2016, 59, 107–112. [Google Scholar] [CrossRef]
- Gronqvist, G.V.; Corner-Thomas, R.A.; Kenyon, P.R.; Stafford, K.J.; Morris, S.T.; Hickson, R.E. The effect of nutrition and body condition of triplet-bearing ewes during late pregnancy on the behaviour of ewes and lambs. Asian-Austr. J. Anim. Sci. 2018, 31, 1991–2000. [Google Scholar] [CrossRef] [PubMed]
- Mathias-Davis, H.C.; Shackell, G.H.; Greer, G.J.; Bryant, A.I.; Everett-Hincks, J.M. Ewe body condition score and the effect on lamb growth rate. Proc. N. Z. Soc. Anim. Prod. 2013, 73, 131–135. [Google Scholar]
- Jalilian, M.T.; Moeini, M.M. The effect of body condition score and body weight of Sanjabi ewes on immune system, productive and reproductive performance. Acta Agric. Slov. 2013, 102, 99–106. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | Year | |||||
|---|---|---|---|---|---|---|
| BB | QFO | p-Value | 1 | 2 | p-Value | |
| BWM | 44.4 | 45.4 | 0.1 | 44.6 | 44.9 | 0.34 |
| BCSM | 1.92 | 1.86 | 0.17 | 2.01 a | 1.78 b | 0.001 |
| BWL | 43.6 | 43.1 | 0.06 | 42.9 | 42.7 | 0.1 |
| BCSL | 1.66 | 1.76 | 0.57 | 1.78 | 1.59 | 0.24 |
| BWW | 41.4 | 41.9 | 0.54 | 41.6 a | 40.0 b | 0.04 |
| BCSW | 1.58 a | 1.42 b | 0.001 | 1.61 a | 1.49 b | 0.001 |
| dBW1 | −0.8 b | −2.3 a | 0.01 | −1.7 b | −2.2 a | 0.03 |
| dBCS1 | −0.26 a | −0.1 b | 0.03 | −0.23 | −0.19 | 0.08 |
| dBW2 | −2.2 a | −1.2 b | 0.01 | −1.3 b | −2.7 a | 0.02 |
| dBCS2 | −0.08 b | −0.34 a | 0.001 | −0.17 | −0.1 | 0.1 |
| Fertility (%) | Prolificacy (%) | ||
|---|---|---|---|
| Breed | BB | 74 | 110 |
| QFO | 75 | 113 | |
| p-value | 0.43 | 0.41 | |
| Year | 1 | 76 | 111 |
| 2 | 72 | 111 | |
| p-value | 0.53 | 0.64 | |
| BW—Mating | BW ≤ 40 (23.4%) | 65 b | 106 |
| 40 < BW < 50 (59.3%) | 76 a | 111 | |
| BW ≥ 50 (17.3%) | 80 a | 115 | |
| p-value | 0.002 | 0.08 | |
| BCS—Mating | BCS < 1.5 (18.7%) | 58 b | 112 |
| 1.5 ≤ BCS < 2 (43.7%) | 77 a | 111 | |
| 2 ≤ BCS < 2.5 (23.9%) | 80 a | 108 | |
| BCS ≥ 2.5 (13.7%) | 75 a | 115 | |
| p-value | 0.001 | 0.5 |
| BWM | BCSM | BWL | BCSL | dBW | dBCS | Fertility | Prolificacy | |
|---|---|---|---|---|---|---|---|---|
| BWM | 1.0 | 0.31 | 0.62 | 0.38 | −0.32 | −0.10 | 0.22 | 0.09 |
| p | <0.0001 | <0.001 | <0.0001 | <0.0001 | 0.006 | <0.0001 | 0.01 | |
| BCSM | 1.0 | −0.28 | 0.21 | 0.01 | 0.07 | 0.97 | −0.11 | |
| p | <0.0001 | <0.0001 | 0.69 | 0.04 | <0.0001 | 0.23 | ||
| BWL | 1.0 | 0.41 | 0.48 | 0.07 | −0.16 | −0.04 | ||
| p | <0.0001 | <0.0001 | 0.04 | <0.0001 | 0.29 | |||
| BCSL | 1.0 | 0.13 | 0.21 | −0.08 | −0.04 | |||
| p | 0.0003 | <0.0001 | 0.01 | 0.28 | ||||
| dBW | 1.0 | 0.29 | 0.05 | 0.04 | ||||
| p | <0.0001 | 0.17 | 0.31 | |||||
| dBCS | 1.0 | 0.37 | 0.40 | |||||
| p | <0.0001 | <0.0001 | ||||||
| Fertility | 1.0 | 0.17 | ||||||
| p | 0.001 | |||||||
| Prolificacy | 1.0 |
| Bi-W | W30 | W70 | ADG0–30 | ADG30–70 | ||
|---|---|---|---|---|---|---|
| Breed | BB | 3.78 | 9.33 | 15.4 | 166 | 176 |
| QFO | 3.79 | 9.82 | 16.04 | 178 | 182 | |
| p-value | 0.62 | 0.1 | 0.3 | 0.29 | 0.35 | |
| Year | 1 | 4.07 a | 9.8 a | 17.52 a | 191 a | 193 a |
| 2 | 3.46 b | 7.9 b | 14.42 b | 148 b | 163 b | |
| p-value | 0.001 | 0.001 | 0.001 | 0.03 | 0.02 | |
| Sex | Male | 3.43 | 8.29 a | 15.13 a | 162 a | 171 a |
| Female | 3.38 | 7.85 b | 14.17 b | 149 b | 158 b | |
| p-value | 0.15 | 0.003 | 0.001 | 0.01 | 0.003 | |
| Birth type | Single | 3.57 a | 8.61 a | 15.57 a | 168 a | 174 a |
| Twin | 2.72 b | 5.84 b | 10.76 b | 104 b | 123 b | |
| p-value | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | |
| BCS-Lambing | BCS < 1.5 (46.6) | 3.79 | 8.41 b | 15.21 b | 154 b | 170 b |
| 1.5 < BCS < 2 (28.2) | 3.73 | 8.86 b | 15.94 b | 171 ab | 177 b | |
| 2 ≤ BCS < 2.5 (16.4) | 3.78 | 9.75 a | 17.55 a | 199 a | 195 a | |
| BCS > 2.5 (8.8) | 3.95 | 9.65 a | 17.21 a | 190 a | 189 a | |
| p-value | 0.07 | 0.001 | 0.001 | 0.01 | 0.01 | |
| BCS-Weaning | BCS < 1.5 (56.1) | 3.74 b | 8.57 b | 15.45 b | 161 b | 172 b |
| 1.5 ≤ BCS < 2 (29.8) | 3.83 b | 9.29 a | 16.61 ab | 182 a | 183 b | |
| 2 < BCS < 2.5 (8.2) | 3.86 b | 9.62 a | 17.62 a | 192 a | 200 a | |
| BCS ≥ 2.5 (5.9) | 4.1 a | 9.35 a | 16.0 ab | 175 b | 165 b | |
| p-value | 0.01 | 0.001 | 0.001 | 0.001 | 0.03 |
| BCSL | BWW | BCSW | dBW | dBCS | Lamb Bi-W | W30 | W70 | ADG0–30 | ADG30–70 | |
|---|---|---|---|---|---|---|---|---|---|---|
| BWL | −0.16 | 0.11 | 0.16 | −0.49 | −0.29 | −0.17 | 0.01 | −0.13 | 0.04 | −0.32 |
| p | 0.0001 | 0.005 | 0.0001 | <0.0001 | <0.0001 | 0.0006 | 0.87 | 0.01 | 0.40 | <0.0001 |
| BCSL | 1.0 | 0.25 | 0.14 | −0.37 | −0.33 | −0.13 | −0.17 | −0.01 | −0.05 | −0.20 |
| p | <0.0001 | 0.0004 | <0.0001 | <0.0001 | 0.008 | 0.002 | 0.78 | 0.29 | <0.0001 | |
| BWW | 1.0 | 0.43 | −0.31 | −0.22 | −0.04 | −0.16 | −0.15 | −0.67 | −0.78 | |
| p | <0.0001 | <0.0001 | <0.0001 | 0.43 | 0.002 | 0.005 | <0.0001 | <0.0001 | ||
| BCSW | 1.0 | −0.12 | 0.09 | 0.07 | 0.18 | 0.10 | 0.26 | −0.91 | ||
| p | 0.005 | 0.04 | 0.18 | 0.001 | 0.04 | <0.0001 | 0.0001 | |||
| dBW | 1.0 | 0.46 | 0.55 | 0.50 | 0.11 | 0.31 | 0.23 | |||
| p | <0.0001 | <0.0001 | <0.0001 | 0.05 | <0.0001 | <0.0001 | ||||
| dBCS | 1.0 | 0.34 | 0.86 | 0.03 | 0.02 | −0.09 | ||||
| p | <0.0001 | <0.0001 | 0.53 | 0.77 | 0.10 | |||||
| Lamb Bi-W | 1.0 | 0.89 | −0.002 | −0.013 | −0.05 | |||||
| p | <0.0001 | 0.96 | 0.80 | 0.38 | ||||||
| W30 | 1.0 | 0.10 | −0.08 | −0.25 | ||||||
| p | 0.07 | 0.12 | <0.0001 | |||||||
| W70 | 1.0 | 0.26 | −0.17 | |||||||
| p | <0.0001 | 0.001 | ||||||||
| ADG0–30 | 1.0 | 0.007 | ||||||||
| p | 0.89 | |||||||||
| ADG30–70 | 1.0 |
| Variable | Coefficients | Standard Error | p-Value |
|---|---|---|---|
| W30; R2 = 0.91 | |||
| Intercept | −20.4 | 5.65 | 0.001 |
| BWL | 1.67 | 0.12 | 0.0001 |
| BCSL | 3.89 | 1.32 | 0.004 |
| BWW | −1.19 | 0.06 | 0.0001 |
| BCSW | 1.52 | 0.04 | 0.0001 |
| dBW | 1.21 | 0.018 | 0.0001 |
| dBCS | 6.87 | 0.58 | 0.0001 |
| W70; R2 = 0.78 | |||
| Intercept | −73.0 | 20.03 | 0.0003 |
| BWL | 0.07 | 0.45 | 0.86 |
| BCSL | 21.0 | 4.81 | 0.0001 |
| BWW | 1.28 | 0.25 | 0.0001 |
| BCSW | −2.012 | 0.15 | 0.0001 |
| dBW | −1.17 | 0.67 | 0.08 |
| dBCS | 28.0 | 2.05 | 0.0001 |
| ADG0–30; R2 = 0.98 | |||
| Intercept | 797.5 | 517.1 | 0.12 |
| BWL | 484.0 | 11.8 | 0.0001 |
| BCSL | 139.5 | 123.3 | 0.25 |
| BWW | −521.27 | 6.4 | 0.0001 |
| BCSW | 487.7 | 3.9 | 0.0001 |
| dBW | 494.2 | 17.0 | 0.0001 |
| dBCS | 42.8 | 53.8 | 0.42 |
| ADG30–70; R2 = 0.95 | |||
| Intercept | 30,450 | 2849.8 | 0.0001 |
| BWL | −8999.2 | 702.7 | 0.0001 |
| BCSL | 2235.4 | 732.4 | 0.003 |
| BWW | 8577.2 | 722.5 | 0.0001 |
| BCSW | −8896.6 | 690.1 | 0.0001 |
| dBW | −8707.3 | 707.7 | 0.0001 |
| dBCS | 2488.9 | 397.6 | 0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yagoubi, Y.; Smeti, S.; Mahouachi, M.; Nasraoui, M.; Ben Saïd, S.; Mohamed-Brahmi, A.; Atti, N. Impact of Body Reserves Dynamic on Productivity and Reproductive Performance in Fat-Tail and Thin-Tail Sheep Breeds over Contrasting Production Cycles. Animals 2024, 14, 891. https://doi.org/10.3390/ani14060891
Yagoubi Y, Smeti S, Mahouachi M, Nasraoui M, Ben Saïd S, Mohamed-Brahmi A, Atti N. Impact of Body Reserves Dynamic on Productivity and Reproductive Performance in Fat-Tail and Thin-Tail Sheep Breeds over Contrasting Production Cycles. Animals. 2024; 14(6):891. https://doi.org/10.3390/ani14060891
Chicago/Turabian StyleYagoubi, Yathreb, Samir Smeti, Mokhtar Mahouachi, Massara Nasraoui, Samia Ben Saïd, Aziza Mohamed-Brahmi, and Naziha Atti. 2024. "Impact of Body Reserves Dynamic on Productivity and Reproductive Performance in Fat-Tail and Thin-Tail Sheep Breeds over Contrasting Production Cycles" Animals 14, no. 6: 891. https://doi.org/10.3390/ani14060891
APA StyleYagoubi, Y., Smeti, S., Mahouachi, M., Nasraoui, M., Ben Saïd, S., Mohamed-Brahmi, A., & Atti, N. (2024). Impact of Body Reserves Dynamic on Productivity and Reproductive Performance in Fat-Tail and Thin-Tail Sheep Breeds over Contrasting Production Cycles. Animals, 14(6), 891. https://doi.org/10.3390/ani14060891