
Molecular Mechanisms of circRNA–miRNA–mRNA Interactions in the Regulation of Goose Liver Development

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Animals and Sampling
2.3. RNA Extraction and Sequencing
2.4. Sequencing Data Analysis
2.5. Construction of the RNA Regulation Network
3. Results
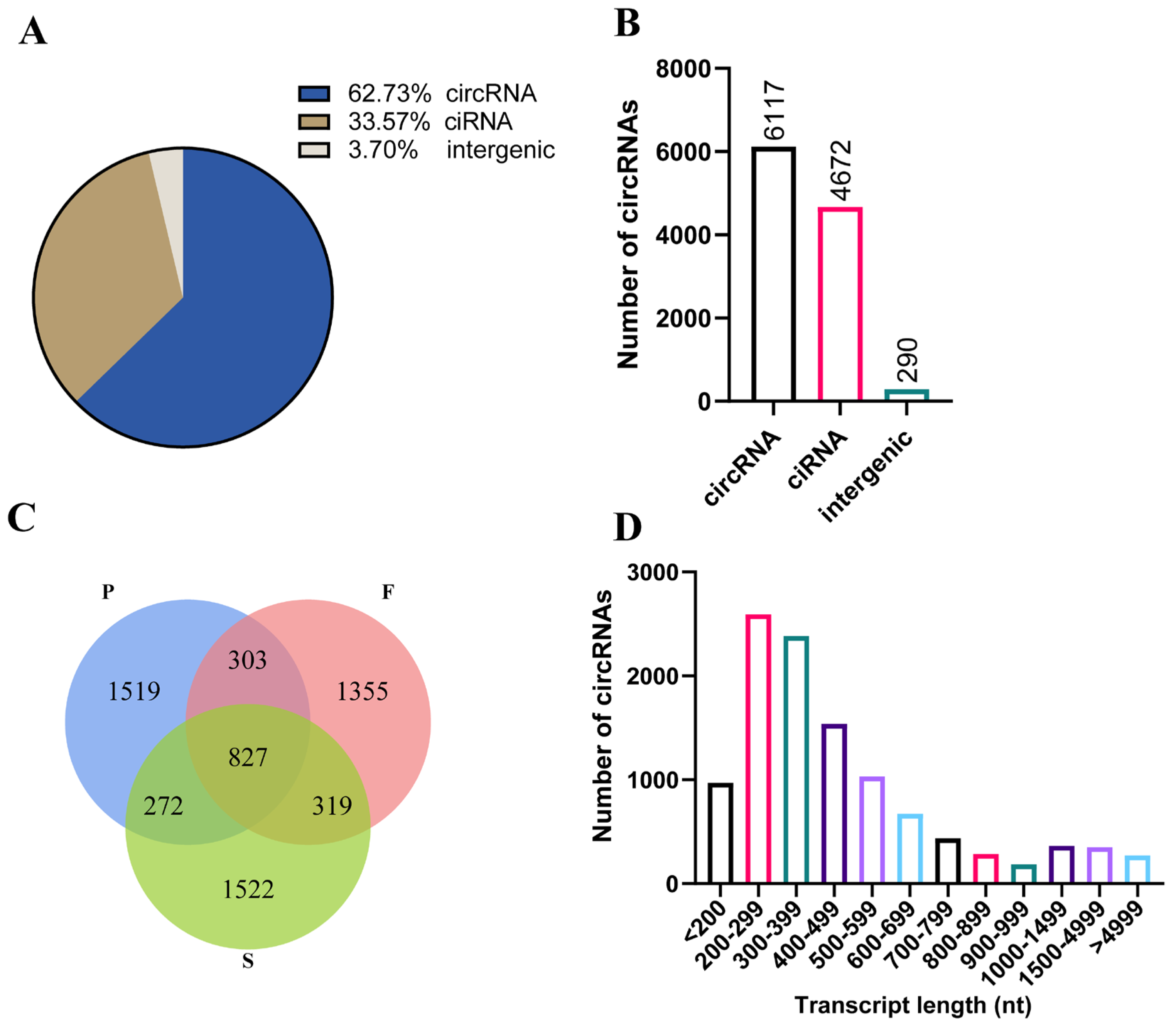
3.1. Identification and Characteristics of circRNAs during Goose Liver Development
3.2. Identification of DEcircRNAs during Goose Liver Development
3.3. Enrichment Analysis of DEcircRNAs during Goose Liver Development
3.4. Expression Profiling and Differential Expression of miRNAs during Goose Liver Development
3.5. Enrichment Analysis of the Target Genes of DEmiRNAs during Goose Liver Development
3.6. Construction of the ceRNA Interaction Regulatory Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turk, D.E. The Anatomy of the Avian Digestive Tract as Related to Feed Utilization. Poult. Sci. 1982, 61, 1225–1244. [Google Scholar] [CrossRef]
- Li, H.; Wang, T.; Xu, C.; Wang, D.; Ren, J.; Li, Y.; Tian, Y.; Wang, Y.; Jiao, Y.; Kang, X.; et al. Transcriptome Profile of Liver at Different Physiological Stages Reveals Potential Mode for Lipid Metabolism in Laying Hens. BMC Genom. 2015, 16, 763. [Google Scholar] [CrossRef]
- Grijalva, J.; Vakili, K. Neonatal Liver Physiology. Semin. Pediatr. Surg. 2013, 22, 185–189. [Google Scholar] [CrossRef]
- Chen, R.; Jiang, X.; Sun, D.; Han, G.; Wang, F.; Ye, M.; Wang, L.; Zou, H. Glycoproteomics Analysis of Human Liver Tissue by Combination of Multiple Enzyme Digestion and Hydrazide Chemistry. J. Proteome Res. 2009, 8, 651–661. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, X.; Huang, J.; Zeng, Y.; Liu, W.; Geng, C.; Li, K.W.; Yang, D.; Wu, S.; Wei, H.; et al. Relationships between Hematopoiesis and Hepatogenesis in the Midtrimester Fetal Liver Characterized by Dynamic Transcriptomic and Proteomic Profiles. PLoS ONE 2009, 4, e7641. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Tao, C.; Xu, J.; Ruan, J.; Xia, J.; Zhu, W.; Xin, L.; Ye, H.; Xie, N.; Xia, B.; et al. CD8+ T Cells Involved in Metabolic Inflammation in Visceral Adipose Tissue and Liver of Transgenic Pigs. Front. Immunol. 2021, 12, 690069. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Jin, L.; Li, Y.; Tang, Q.; Hu, S.; Xu, H.; Gill, C.A.; Li, M.; Wang, J. Transcriptomic Analysis between Normal and High-Intake Feeding Geese Provides Insight into Adipose Deposition and Susceptibility to Fatty Liver in Migratory Birds. BMC Genom. 2019, 20, 372. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Chen, Y.; Wang, Z.; Li, X.; Chen, W.; Tao, Z.; Shen, J.; Tian, Y.; Wang, D.; Li, G.; et al. The Goose Genome Sequence Leads to Insights into the Evolution of Waterfowl and Susceptibility to Fatty Liver. Genome Biol. 2015, 16, 89. [Google Scholar] [CrossRef] [PubMed]
- Davail, S.; Guy, G.; André, J.; Hermier, D.; Hoo-Paris, R. Metabolism in Two Breeds of Geese with Moderate or Large Overfeeding Induced Liver-Steatosis. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2000, 126, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Ning, R.; Han, C.; Wei, S.; Teng, Y.; Li, L.; Liu, H.; Hu, S.; Kang, B.; Xu, H. Lipidomics Analysis Reveals New Insights into the Goose Fatty Liver Formation. Poult. Sci. 2023, 102, 102428. [Google Scholar] [CrossRef] [PubMed]
- Xu, E.; Zhang, L.; Yang, H.; Shen, L.; Feng, Y.; Ren, M.; Xiao, Y. Transcriptome Profiling of the Liver among the Prenatal and Postnatal Stages in Chickens. Poult. Sci. 2019, 98, 7030–7040. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs Are a Large Class of Animal RNAs with Regulatory Potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef]
- Chen, L.-L. The Biogenesis and Emerging Roles of Circular RNAs. Nat. Rev. Mol. Cell Biol. 2016, 17, 205–211. [Google Scholar] [CrossRef]
- Dong, R.; Zhang, X.-O.; Zhang, Y.; Ma, X.-K.; Chen, L.-L.; Yang, L. CircRNA-Derived Pseudogenes. Cell Res. 2016, 26, 747–750. [Google Scholar] [CrossRef]
- Lin, Z.; Xie, F.; He, X.; Wang, J.; Luo, J.; Chen, T.; Jiang, Q.; Xi, Q.; Zhang, Y.; Sun, J. A Novel Protein Encoded by circKANSL1L Regulates Skeletal Myogenesis via the Akt-FoxO3 Signaling Axis. Int. J. Biol. Macromol. 2024, 257, 128609. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wei, X.; Song, M.; Jiang, R.; Huang, K.; Deng, Y.; Liu, Q.; Shi, D.; Li, H. Circular RNA circMYBPC1 Promotes Skeletal Muscle Differentiation by Targeting MyHC. Mol. Ther. Nucleic Acids 2021, 24, 352–368. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Sun, T.; Yang, Z.; Zou, L.; Deng, J.; Yang, X. Whole-Transcriptome RNA Sequencing Reveals the Global Molecular Responses and circRNA/lncRNA-miRNA-mRNA ceRNA Regulatory Network in Chicken Fat Deposition. Poult. Sci. 2022, 101, 102121. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Ma, H.; Li, B.; Yang, F.; Xiao, Y.; Gong, Y.; Li, Z.; Li, T.; Zeng, Q.; Xu, K.; et al. Spatiotemporal Regulation of Circular RNA Expression during Liver Development of Chinese Indigenous Ningxiang Pigs. Genes 2022, 13, 746. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xiong, T.; Zhou, M.; Wan, L.; Xi, S.; Liu, Q.; Chen, Y.; Mao, H.; Liu, S.; Chen, B. Characterization of microRNAs during Embryonic Skeletal Muscle Development in the Shan Ma Duck. Animals 2020, 10, 1417. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yan, P.; Liu, L.; Luo, Y.; Zhao, L.; Liu, H.; Tang, Q.; Long, K.; Jin, L.; Ma, J.; et al. MicroRNA Expression Profiling Reveals Potential Roles for microRNA in the Liver during Pigeon (Columba livia) Development. Poult. Sci. 2020, 99, 6378–6389. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wu, Y.; Zhang, Y.; Tang, S.; Han, S.; Cui, C.; Tan, B.; Yu, J.; Kang, H.; Chen, G.; et al. miR-27b-5p Regulates Chicken Liver Disease via Targeting IRS2 to Suppress the PI3K/AKT Signal Pathway. J. Integr. Agric. 2023, 22, 3500–3516. [Google Scholar] [CrossRef]
- Wei, R.; Han, C.; Wei, S.; Teng, Y.; Li, L.; Liu, H.; Hu, S.; Kang, B.; Xu, H. Integrative Analysis of Transcriptome and Lipidome Reveals Fructose Pro-Steatosis Mechanism in Goose Fatty Liver. Front. Nutr. 2022, 9, 1052600. [Google Scholar] [CrossRef]
- Chen, Z.; Xing, Y.; Fan, X.; Liu, T.; Zhao, M.; Liu, L.; Hu, X.; Cui, H.; Geng, T.; Gong, D. Fasting and Refeeding Affect the Goose Liver Transcriptome Mainly Through the PPAR Signaling Pathway. J. Poult. Sci. 2021, 58, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Q.; Hu, S.; Tang, B.; Hu, B.; Hu, J.; He, H.; Li, L.; Wang, J. Comparative Transcriptome Analysis Provides Novel Insights into the Effect of Lipid Metabolism on Laying of Geese. Animals 2022, 12, 1775. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, S.; Zhang, W.; Hu, X.; Mao, H.; Liu, S.; Chen, B. Transcriptome RNA Sequencing Reveals That Circular RNAs Are Abundantly Expressed in Embryonic Breast Muscle of Duck. Vet. Sci. 2023, 10, 75. [Google Scholar] [CrossRef]
- Kechin, A.; Boyarskikh, U.; Kel, A.; Filipenko, M. cutPrimers: A New Tool for Accurate Cutting of Primers from Reads of Targeted Next Generation Sequencing. J. Comput. Biol. 2017, 24, 1138–1143. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate Alignment of Transcriptomes in the Presence of Insertions, Deletions and Gene Fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef]
- Kim, D.; Salzberg, S.L. TopHat-Fusion: An Algorithm for Discovery of Novel Fusion Transcripts. Genome Biol. 2011, 12, R72. [Google Scholar] [CrossRef]
- Zhang, X.-O.; Wang, H.-B.; Zhang, Y.; Lu, X.; Chen, L.-L.; Yang, L. Complementary Sequence-Mediated Exon Circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef]
- Zhang, X.-O.; Dong, R.; Zhang, Y.; Zhang, J.-L.; Luo, Z.; Zhang, J.; Chen, L.-L.; Yang, L. Diverse Alternative Back-Splicing and Alternative Splicing Landscape of Circular RNAs. Genome Res. 2016, 26, 1277–1287. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, J.; Zhao, F. CIRI: An Efficient and Unbiased Algorithm for de Novo Circular RNA Identification. Genome Biol. 2015, 16, 4. [Google Scholar] [CrossRef]
- Nam, J.-W.; Rissland, O.S.; Koppstein, D.; Abreu-Goodger, C.; Jan, C.H.; Agarwal, V.; Yildirim, M.A.; Rodriguez, A.; Bartel, D.P. Global Analyses of the Effect of Different Cellular Contexts on microRNA Targeting. Mol. Cell 2014, 53, 1031–1043. [Google Scholar] [CrossRef]
- Betel, D.; Koppal, A.; Agius, P.; Sander, C.; Leslie, C. Comprehensive Modeling of microRNA Targets Predicts Functional Non-Conserved and Non-Canonical Sites. Genome Biol. 2010, 11, R90. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Zhou, R.; Wu, Y.; Wang, W.; Su, W.; Liu, Y.; Wang, Y.; Fan, C.; Li, X.; Li, G.; Li, Y.; et al. Circular RNAs (circRNAs) in Cancer. Cancer Lett. 2018, 425, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Liang, M.; Liu, H.; Huang, J.; Li, P.; Wang, C.; Zhang, Y.; Lin, Y.; Jiang, X. CircRNA hsa_circRNA_104348 Promotes Hepatocellular Carcinoma Progression through Modulating miR-187-3p/RTKN2 Axis and Activating Wnt/β-Catenin Pathway. Cell Death Dis. 2020, 11, 1065. [Google Scholar] [CrossRef]
- Kang, B.; Jiang, D.M.; Bai, L.; He, H.; Ma, R. Molecular Characterisation and Expression Profiling of the ENO1 Gene in the Ovarian Follicle of the Sichuan White Goose. Mol. Biol. Rep. 2014, 41, 1927–1935. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, Y.-Y.; Chen, P.-P.; Xu, H.; Xie, S.-J.; Xu, S.-J.; Li, B.; Li, J.-H.; Liu, S.; Yang, J.-H.; et al. Genome-Wide Identification of microRNA Targets Reveals Positive Regulation of the Hippo Pathway by miR-122 during Liver Development. Cell Death Dis. 2021, 12, 1161. [Google Scholar] [CrossRef]
- Chen, Y.; Verfaillie, C.M. MicroRNAs: The Fine Modulators of Liver Development and Function. Liver Int. 2014, 34, 976–990. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Gong, W.; Pan, X.; Cai, J.; Jiang, Y.; He, M.; Zhao, S.; Li, Y.; Yuan, X.; Li, J. Comprehensive Analysis of CircRNA Expression Profiles in Multiple Tissues of Pigs. Int. J. Mol. Sci. 2023, 24, 16205. [Google Scholar] [CrossRef]
- Gu, Y.; Wang, Y.; He, L.; Zhang, J.; Zhu, X.; Liu, N.; Wang, J.; Lu, T.; He, L.; Tian, Y.; et al. Circular RNA circIPO11 Drives Self-Renewal of Liver Cancer Initiating Cells via Hedgehog Signaling. Mol. Cancer 2021, 20, 132. [Google Scholar] [CrossRef]
- Ouyang, H.; Chen, X.; Wang, Z.; Yu, J.; Jia, X.; Li, Z.; Luo, W.; Abdalla, B.A.; Jebessa, E.; Nie, Q.; et al. Circular RNAs Are Abundant and Dynamically Expressed during Embryonic Muscle Development in Chickens. DNA Res. 2018, 25, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Xiao, H.; Pi, J.; Zhang, H.; Pan, A.; Pu, Y.; Liang, Z.; Shen, J.; Du, J. The Circular RNA Aplacirc_13267 Upregulates Duck Granulosa Cell Apoptosis by the Apla-miR-1-13/THBS1 Signaling Pathway. J. Cell. Physiol. 2020, 235, 5750–5763. [Google Scholar] [CrossRef]
- Qi, X.; Hu, Q.; Elghobashi-Meinhardt, N.; Long, T.; Chen, H.; Li, X. Molecular Basis of Wnt Biogenesis, Secretion, and Wnt7-Specific Signaling. Cell 2023, 186, 5028–5040.e14. [Google Scholar] [CrossRef]
- Chai, G.; Szenker-Ravi, E.; Chung, C.; Li, Z.; Wang, L.; Khatoo, M.; Marshall, T.; Jiang, N.; Yang, X.; McEvoy-Venneri, J.; et al. A Human Pleiotropic Multiorgan Condition Caused by Deficient Wnt Secretion. N. Engl. J. Med. 2021, 385, 1292–1301. [Google Scholar] [CrossRef] [PubMed]
- Leibing, T.; Géraud, C.; Augustin, I.; Boutros, M.; Augustin, H.G.; Okun, J.G.; Langhans, C.-D.; Zierow, J.; Wohlfeil, S.A.; Olsavszky, V.; et al. Angiocrine Wnt Signaling Controls Liver Growth and Metabolic Maturation in Mice. Hepatology 2018, 68, 707–722. [Google Scholar] [CrossRef]
- Wang, W.; Xu, L.; Liu, P.; Jairam, K.; Yin, Y.; Chen, K.; Sprengers, D.; Peppelenbosch, M.P.; Pan, Q.; Smits, R. Blocking Wnt Secretion Reduces Growth of Hepatocellular Carcinoma Cell Lines Mostly Independent of β-Catenin Signaling. Neoplasia 2016, 18, 711–723. [Google Scholar] [CrossRef]
- Lu, W.; Liu, Y.; Gao, Y.; Geng, Q.; Gurbani, D.; Li, L.; Ficarro, S.B.; Meyer, C.J.; Sinha, D.; You, I.; et al. Development of a Covalent Inhibitor of C-Jun N-Terminal Protein Kinase (JNK) 2/3 with Selectivity over JNK1. J. Med. Chem. 2023, 66, 3356–3371. [Google Scholar] [CrossRef]
- Bost, F.; Aouadi, M.; Caron, L.; Binétruy, B. The Role of MAPKs in Adipocyte Differentiation and Obesity. Biochimie 2005, 87, 51–56. [Google Scholar] [CrossRef]
- Treisman, R. Regulation of Transcription by MAP Kinase Cascades. Curr. Opin. Cell Biol. 1996, 8, 205–215. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, J.; Liu, Y.; Li, J.; Chang, C.; Xu, C. Rat Hepatocytes Weighted Gene Co-Expression Network Analysis Identifies Specific Modules and Hub Genes Related to Liver Regeneration after Partial Hepatectomy. PLoS ONE 2014, 9, e94868. [Google Scholar] [CrossRef]
- Roach, P.J.; Depaoli-Roach, A.A.; Hurley, T.D.; Tagliabracci, V.S. Glycogen and Its Metabolism: Some New Developments and Old Themes. Biochem. J. 2012, 441, 763–787. [Google Scholar] [CrossRef]
- Li, Y.; Zhuang, H.; Zhang, X.; Li, Y.; Liu, Y.; Yi, X.; Qin, G.; Wei, W.; Chen, R. Multiomics Integration Reveals the Landscape of Prometastasis Metabolism in Hepatocellular Carcinoma. Mol. Cell Proteom. 2018, 17, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Thakral, S.; Ghoshal, K. miR-122 Is a Unique Molecule with Great Potential in Diagnosis, Prognosis of Liver Disease, and Therapy Both as miRNA Mimic and Antimir. Curr. Gene Ther. 2015, 15, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Q.; Zhao, X.; Wang, L.; Wang, X.; Wang, J.; Dong, B.; Gong, D. MicroRNA-122 Targets Genes Related to Goose Fatty Liver. Poult. Sci. 2018, 97, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-H.; Jiang, W.; Zeng, Q.; Wu, D.; Li, H.; Zhou, L.; Bai, L.; Tang, H. CircRNA-PI4KB Induces Hepatic Lipid Deposition in Non-Alcoholic Fatty Liver Disease by Transporting miRNA-122 to Extra-Hepatocytes. Int. J. Mol. Sci. 2023, 24, 1297. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.-H.; Liu, Z.-Y.; Zhang, D.-Y.; Zhang, S.; Tang, W.-Q.; Li, D.-P.; Zhang, F.; Chen, R.-X.; Weng, S.-Q.; Xue, R.-Y.; et al. circRanGAP1/miR-27b-3p/NRAS Axis May Promote the Progression of Hepatocellular Carcinoma. Exp. Hematol. Oncol. 2022, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Jiang, G.; Shao, J.; Wang, M.; Zhang, X.; Xia, S.; Sun, W.; Jia, X.; Wang, J.; Lai, S. lncRNA MSTRG4710 Promotes the Proliferation and Differentiation of Preadipocytes through miR-29b-3p/IGF1 Axis. Int. J. Mol. Sci. 2023, 24, 15715. [Google Scholar] [CrossRef] [PubMed]
- Dalgaard, L.T.; Sørensen, A.E.; Hardikar, A.A.; Joglekar, M.V. The microRNA-29 Family: Role in Metabolism and Metabolic Disease. Am. J. Physiol. Cell Physiol. 2022, 323, C367–C377. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, C.L.; Peck, B.C.E.; Fannin, E.E.; Beysen, C.; Miao, J.; Landstreet, S.R.; Ding, S.; Turaga, V.; Lund, P.K.; Turner, S.; et al. MicroRNA-29 Fine-Tunes the Expression of Key FOXA2-Activated Lipid Metabolism Genes and Is Dysregulated in Animal Models of Insulin Resistance and Diabetes. Diabetes 2014, 63, 3141–3148. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Liu, C.; Qiao, A.; Cui, Y.; Zhang, H.; Cui, A.; Zhang, S.; Yang, Y.; Xiao, X.; Chen, Y.; et al. MicroRNA-29a-c Decrease Fasting Blood Glucose Levels by Negatively Regulating Hepatic Gluconeogenesis. J. Hepatol. 2013, 58, 535–542. [Google Scholar] [CrossRef]
- Pandey, A.K.; Verma, G.; Vig, S.; Srivastava, S.; Srivastava, A.K.; Datta, M. miR-29a Levels Are Elevated in the Db/Db Mice Liver and Its Overexpression Leads to Attenuation of Insulin Action on PEPCK Gene Expression in HepG2 Cells. Mol. Cell. Endocrinol. 2011, 332, 125–133. [Google Scholar] [CrossRef]
- Zhang, J.; Xing, Y.; Li, F.; Mu, J.; Liu, T.; Ge, J.; Zhao, M.; Liu, L.; Gong, D.; Geng, T. Study on the Mechanism of MC5R Participating in Energy Metabolism of Goose Liver. Int. J. Mol. Sci. 2023, 24, 8648. [Google Scholar] [CrossRef]
- Trefts, E.; Gannon, M.; Wasserman, D.H. The Liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef]
- Han, H.-S.; Kang, G.; Kim, J.S.; Choi, B.H.; Koo, S.-H. Regulation of Glucose Metabolism from a Liver-Centric Perspective. Exp. Mol. Med. 2016, 48, e218. [Google Scholar] [CrossRef]
- Jones, J.G. Hepatic Glucose and Lipid Metabolism. Diabetologia 2016, 59, 1098–1103. [Google Scholar] [CrossRef]
- Upadhya, D.; Ogata, M.; Reneker, L.W. MAPK1 Is Required for Establishing the Pattern of Cell Proliferation and for Cell Survival during Lens Development. Development 2013, 140, 1573–1582. [Google Scholar] [CrossRef]
- Wang, H.; Ke, J.; Guo, Q.; Barnabo Nampoukime, K.-P.; Yang, P.; Ma, K. Long Non-Coding RNA CRNDE Promotes the Proliferation, Migration and Invasion of Hepatocellular Carcinoma Cells through miR-217/MAPK1 Axis. J. Cell Mol. Med. 2018, 22, 5862–5876. [Google Scholar] [CrossRef]
- Hussain, S.; Sun, M.; Guo, Y.; Mushtaq, N.; Zhao, Y.; Yuan, Y.; Hussain, N.; Osoro, E.; Suleiman, A.; Sadiq, M.; et al. SFMBT2 Positively Regulates SOX9 and Chondrocyte Proliferation. Int. J. Mol. Med. 2018, 42, 3503–3512. [Google Scholar] [CrossRef]
- Dommann, N.; Gavini, J.; Sánchez-Taltavull, D.; Baier, F.A.; Birrer, F.; Loforese, G.; Candinas, D.; Stroka, D. LIM Protein Ajuba Promotes Liver Cell Proliferation through Its Involvement in DNA Replication and DNA Damage Control. FEBS Lett. 2022, 596, 1746–1764. [Google Scholar] [CrossRef]
- Wang, B.; Kaufmann, B.; Engleitner, T.; Lu, M.; Mogler, C.; Olsavszky, V.; Öllinger, R.; Zhong, S.; Geraud, C.; Cheng, Z.; et al. Brg1 Promotes Liver Regeneration after Partial Hepatectomy via Regulation of Cell Cycle. Sci. Rep. 2019, 9, 2320. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Deng, D.; Teng, Y.; Lu, C.; Luo, Z.; Abdulai, M.; Liu, H.; Xu, H.; Li, L.; Hu, S.; et al. Study on the Effect of Different Types of Sugar on Lipid Deposition in Goose Fatty Liver. Poult. Sci. 2022, 101, 101729. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Dong, H.H. FoxO Integration of Insulin Signaling with Glucose and Lipid Metabolism. J. Endocrinol. 2017, 233, R67–R79. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identification Number | Read | Q30% | Valid Reads | Mapped Reads | Back-Spliced Junctions Reads | Number of circRNA-Hosting Genes | Number of circRNAs |
|---|---|---|---|---|---|---|---|
| P1 | 182,770,542 | 96.02 | 94,846,238 | 63,808,104 (67.28%) | 920,007 (0.97%) | 1573 | 2385 |
| P2 | 194,923,376 | 95.70 | 89,396,850 | 59,130,157 (66.14%) | 855,279 (0.96%) | 1356 | 1943 |
| P3 | 167,873,330 | 96.29 | 96,477,610 | 69,367,040 (71.90%) | 844,581 (0.88%) | 1458 | 2120 |
| F1 | 130,003,024 | 96.80 | 81,026,888 | 60,809,218 (75.05%) | 813,446 (1.00%) | 1486 | 2176 |
| F2 | 134,426,502 | 96.67 | 91,167,744 | 67,387,484 (73.92%) | 1,268,977 (1.39%) | 1315 | 1882 |
| F3 | 151,546,164 | 96.65 | 86,646,924 | 64,557,846 (74.51%) | 830,104 (0.96%) | 1351 | 1975 |
| S1 | 151,401,712 | 96.35 | 88,253,086 | 63,009,145 (71.40%) | 843,406 (0.96%) | 1497 | 2180 |
| S2 | 149,935,366 | 96.66 | 88,416,132 | 64,772,234 (73.26%) | 782,180 (0.88%) | 1689 | 2529 |
| S3 | 127,097,270 | 96.75 | 67,356,910 | 51,322,862 (76.20%) | 637,003 (0.95%) | 1237 | 1733 |
| Identification Number | Raw Reads | Clean Reads | Clean Ratio, % | Valid Reads | Valid Ratio, % |
|---|---|---|---|---|---|
| P1 | 10,696,086 | 8,876,088 | 82.98 | 7,316,506 | 68.40 |
| P2 | 11,729,357 | 9,302,214 | 79.31 | 7,922,998 | 67.55 |
| P3 | 11,681,199 | 9,556,620 | 81.82 | 8,077,834 | 69.15 |
| F1 | 10,880,983 | 9,258,042 | 85.08 | 7,994,024 | 73.47 |
| F2 | 12,183,931 | 10,423,708 | 85.55 | 8,759,628 | 71.89 |
| F3 | 10,290,009 | 8,546,903 | 83.06 | 7,324,075 | 71.18 |
| S1 | 11,750,317 | 9,597,940 | 81.69 | 7,928,524 | 67.47 |
| S2 | 11,457,809 | 9,158,427 | 79.93 | 7,615,894 | 66.47 |
| S3 | 11,728,045 | 8,123,358 | 69.27 | 6,952,832 | 59.28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Li, C.; Hu, X.; Mao, H.; Liu, S.; Chen, B. Molecular Mechanisms of circRNA–miRNA–mRNA Interactions in the Regulation of Goose Liver Development. Animals 2024, 14, 839. https://doi.org/10.3390/ani14060839
Liu S, Li C, Hu X, Mao H, Liu S, Chen B. Molecular Mechanisms of circRNA–miRNA–mRNA Interactions in the Regulation of Goose Liver Development. Animals. 2024; 14(6):839. https://doi.org/10.3390/ani14060839
Chicago/Turabian StyleLiu, Shuibing, Chuan Li, Xiaolong Hu, Huirong Mao, Sanfeng Liu, and Biao Chen. 2024. "Molecular Mechanisms of circRNA–miRNA–mRNA Interactions in the Regulation of Goose Liver Development" Animals 14, no. 6: 839. https://doi.org/10.3390/ani14060839
APA StyleLiu, S., Li, C., Hu, X., Mao, H., Liu, S., & Chen, B. (2024). Molecular Mechanisms of circRNA–miRNA–mRNA Interactions in the Regulation of Goose Liver Development. Animals, 14(6), 839. https://doi.org/10.3390/ani14060839