
AhR Activation Ameliorates Intestinal Barrier Damage in Immunostressed Piglets by Regulating Intestinal Flora and Its Metabolism

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design, Diet, Sample Collection
2.2. Intestinal Morphology
2.3. Immunofluorescence
2.4. Real-Time Quantitative PCR Analysis
2.5. Western Blotting Analysis
2.6. Gut Microbiota Analysis
2.7. Short-Chain Fatty Acid (SCFA) Analysis
2.8. Ultra-High-Performance Liquid Chromatography Tandem Quadrupole Time-of-Flight Mass Spectrometry (UHPLC-QTOFMS) Analysis
2.9. Statistical Analysis
3. Results
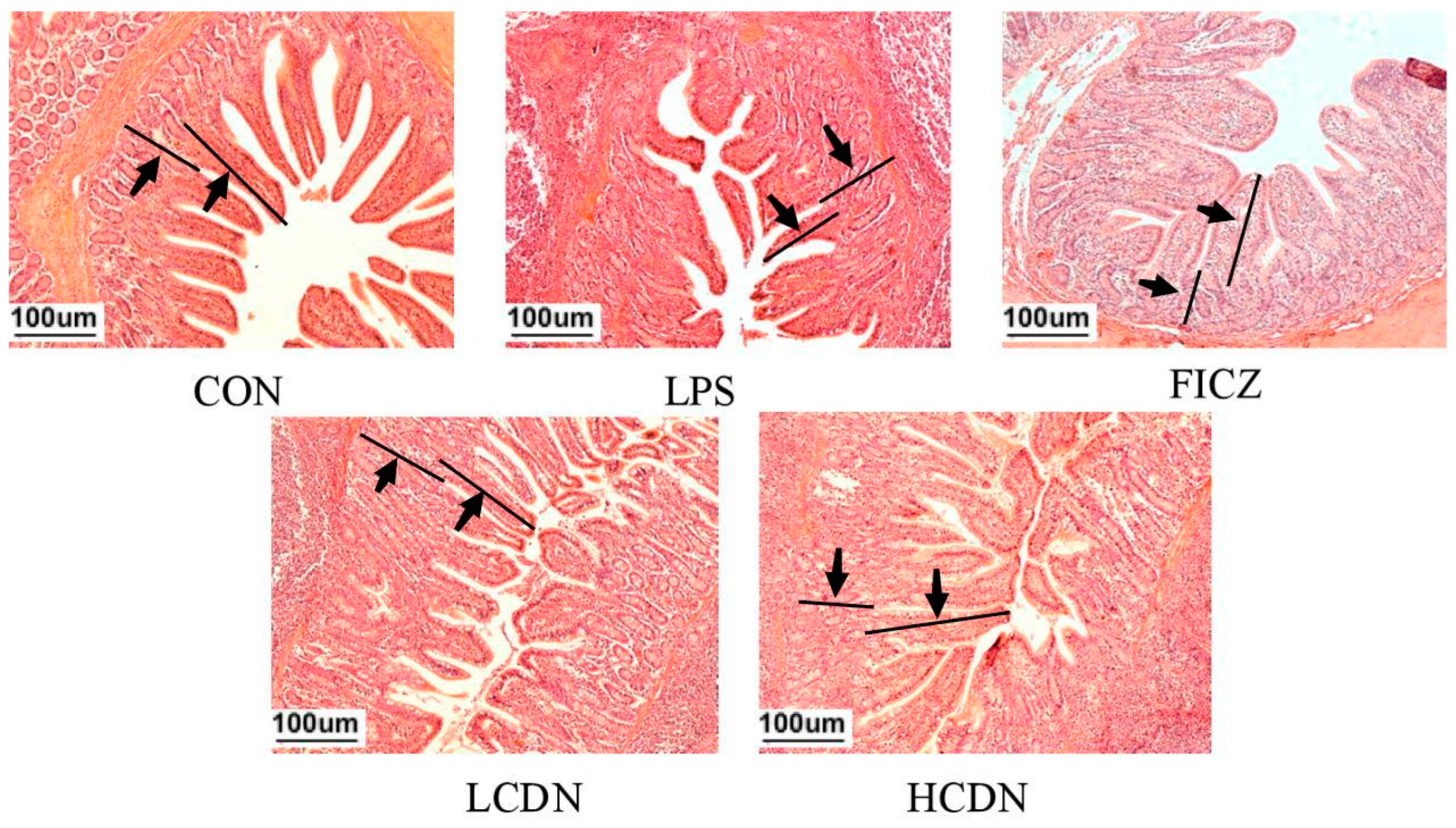
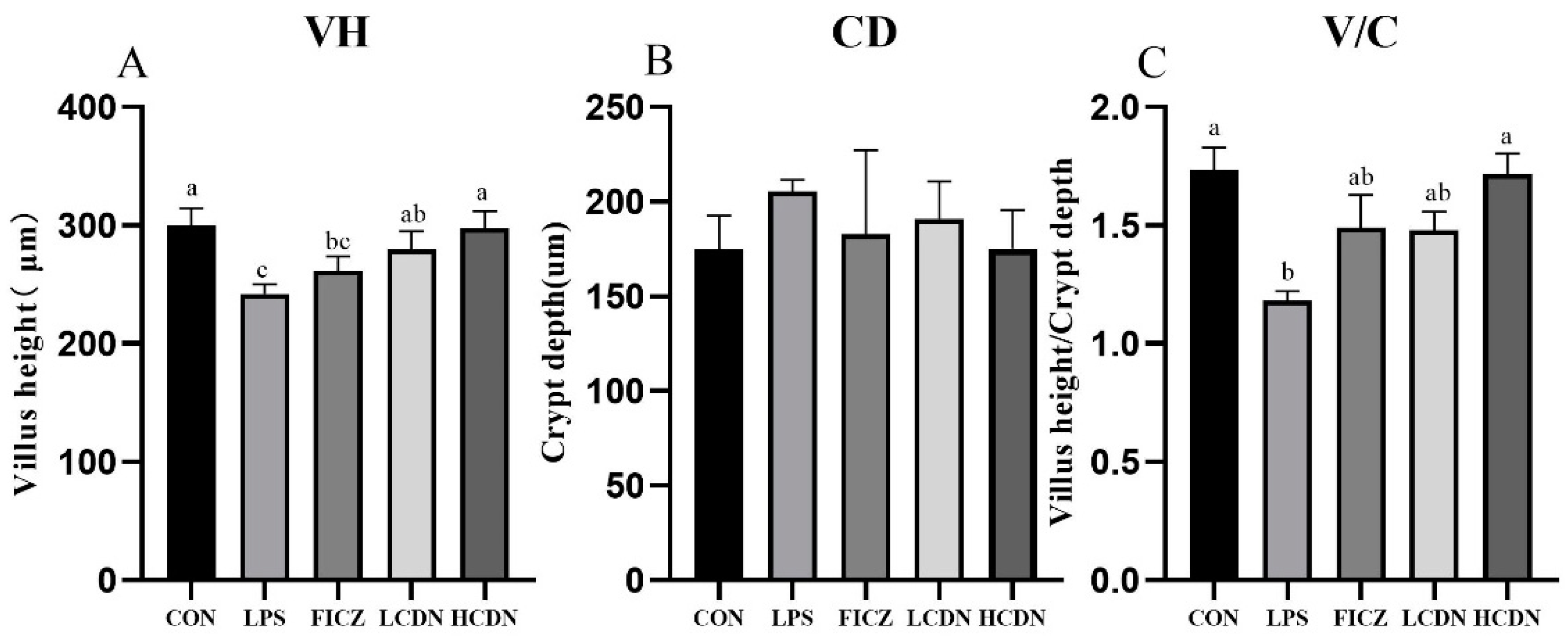
3.1. Ileal Mucosa Morphology
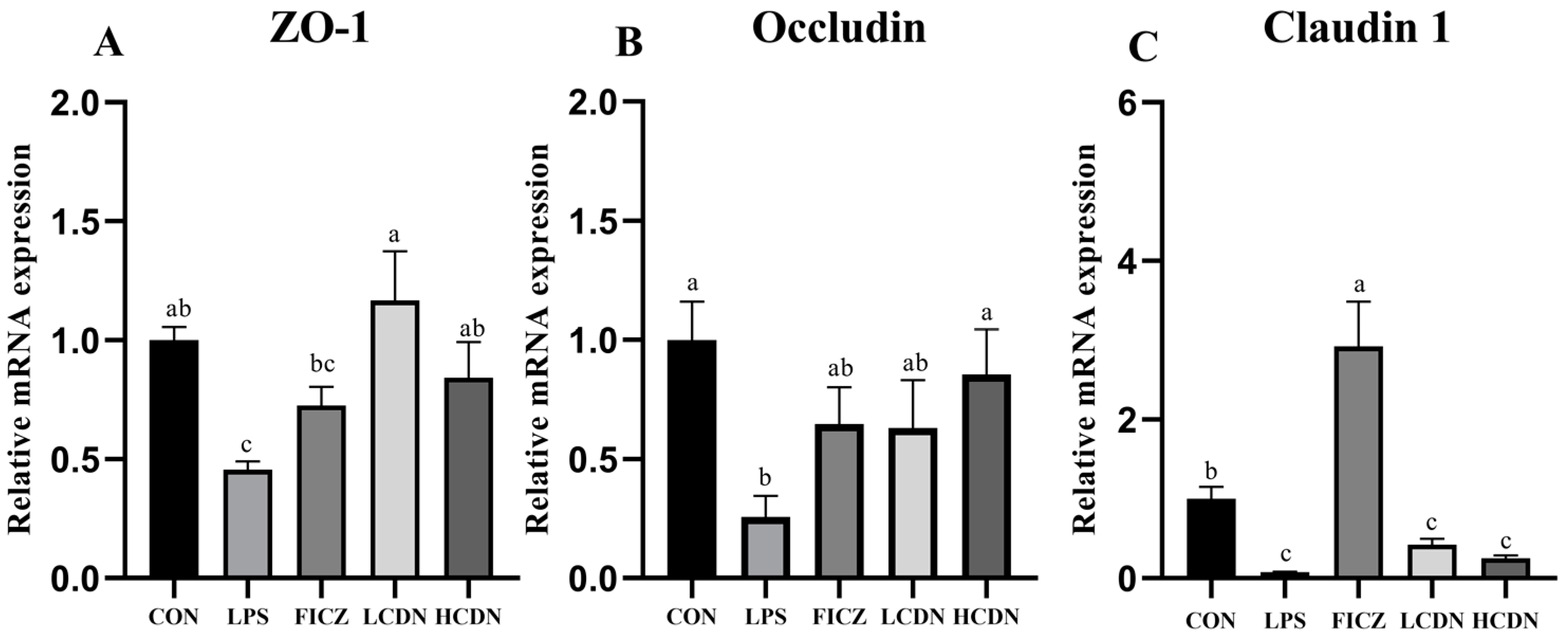
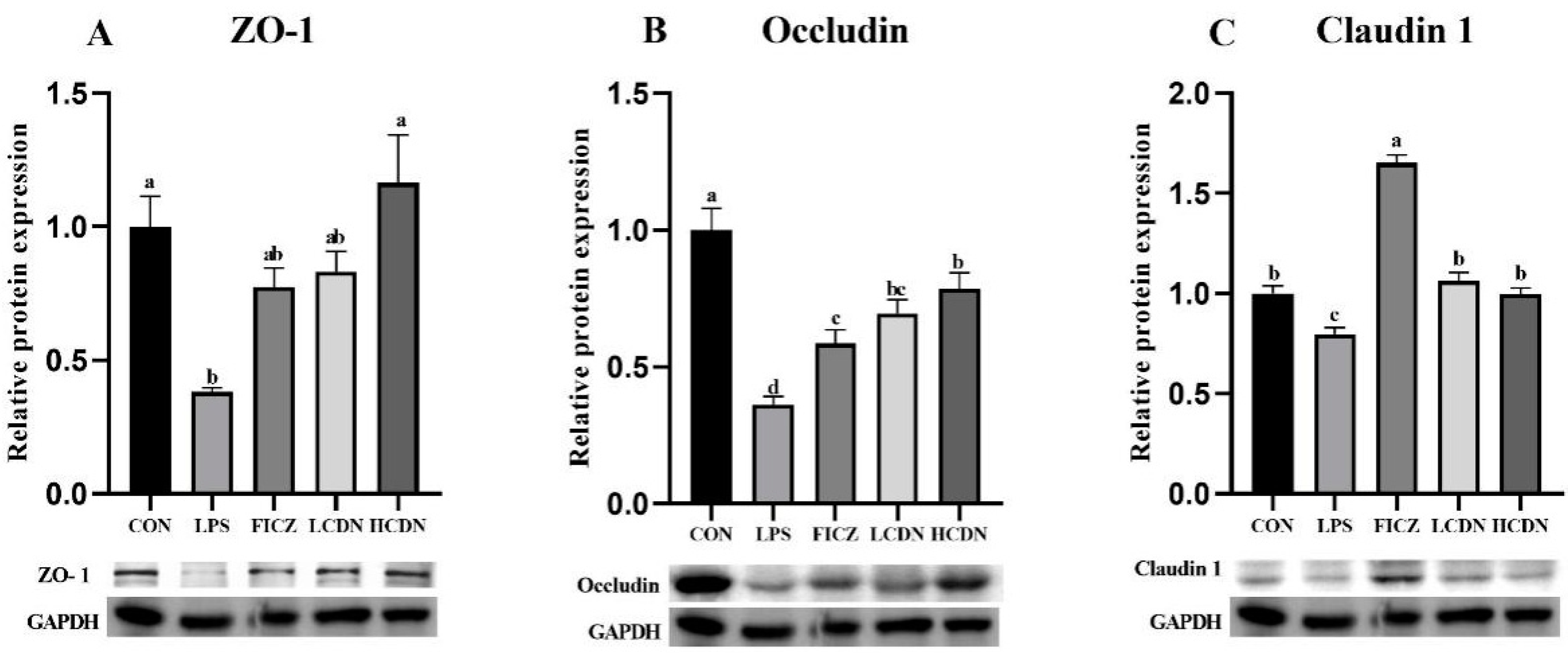
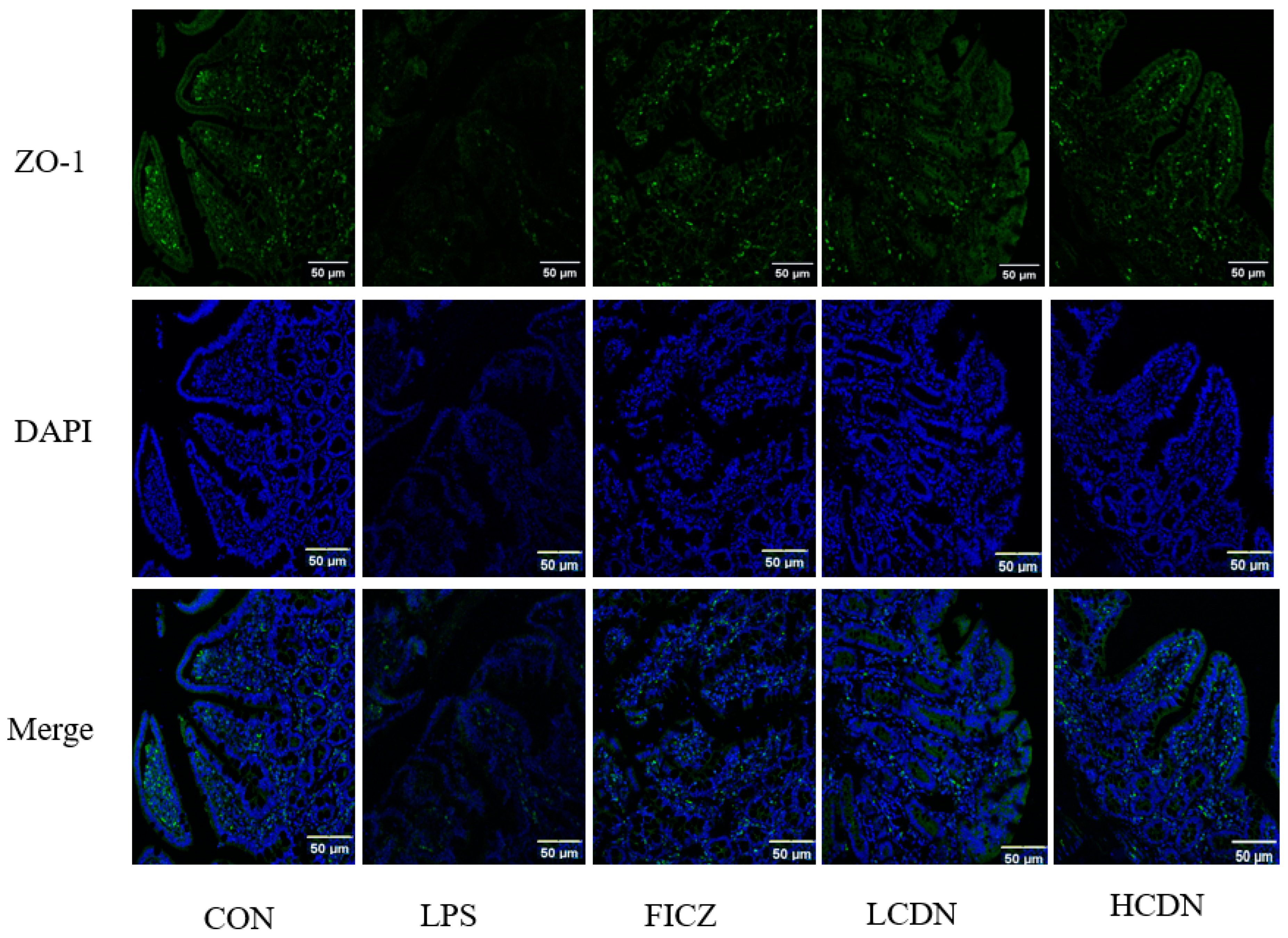
3.2. Intestinal Barrier Analysis
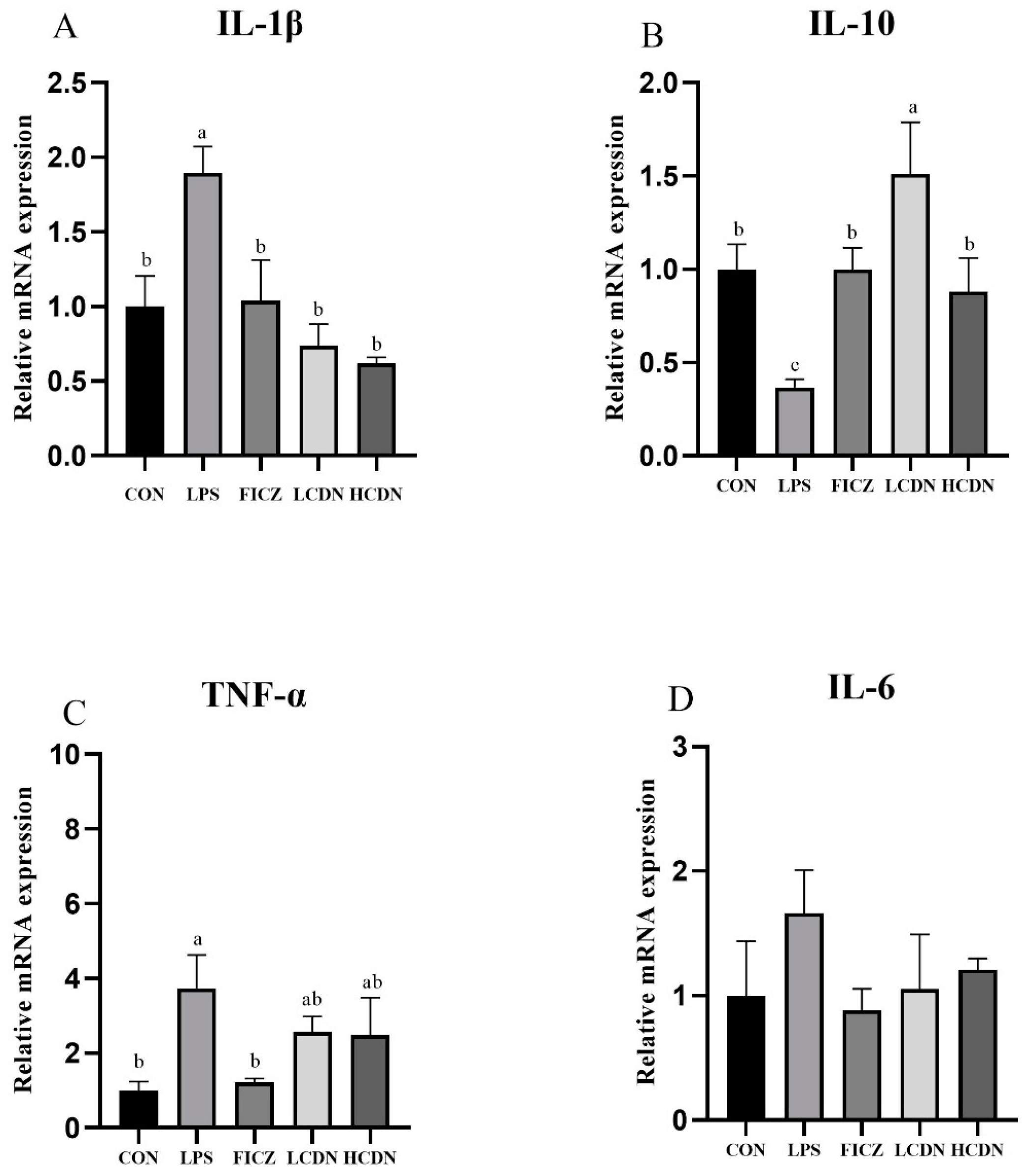
3.3. mRNA Expression of Cytokine
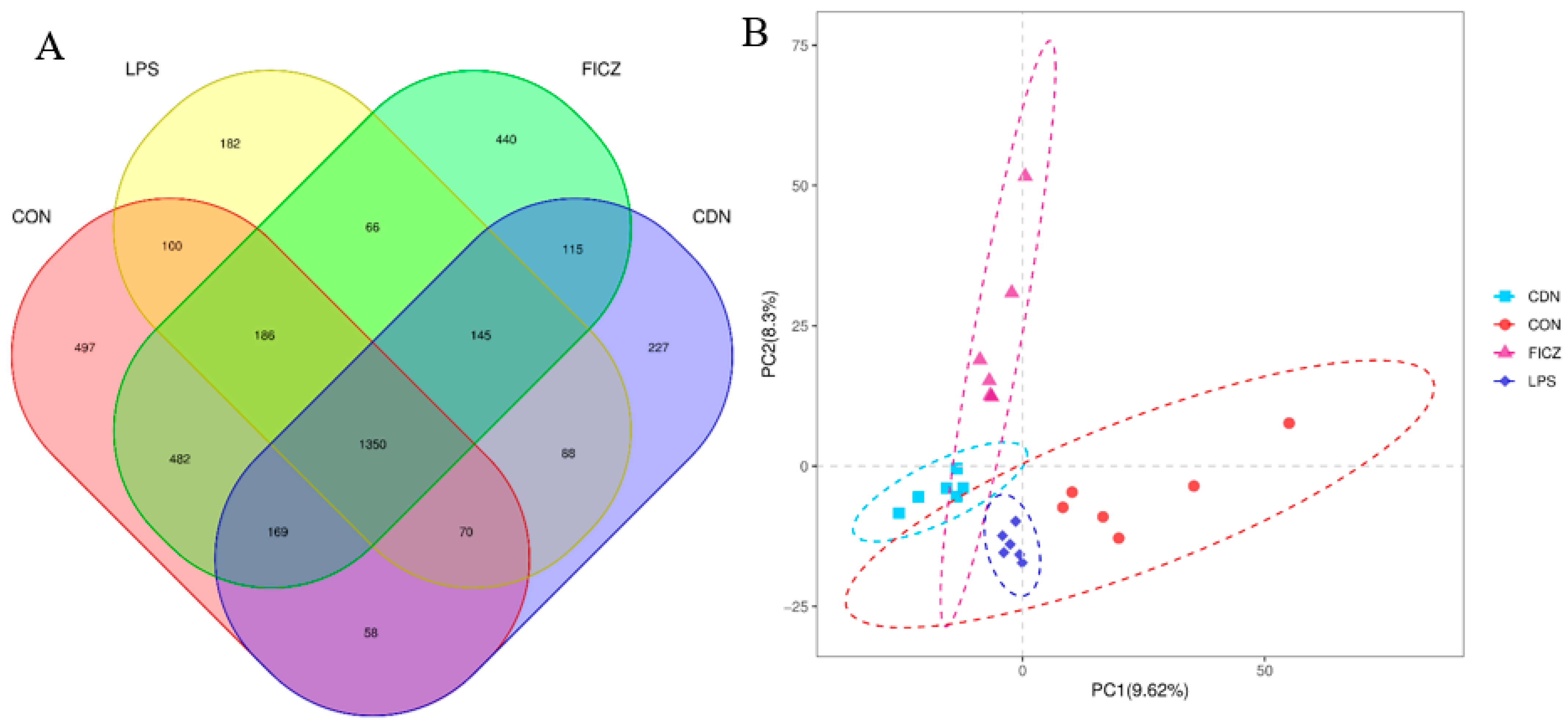
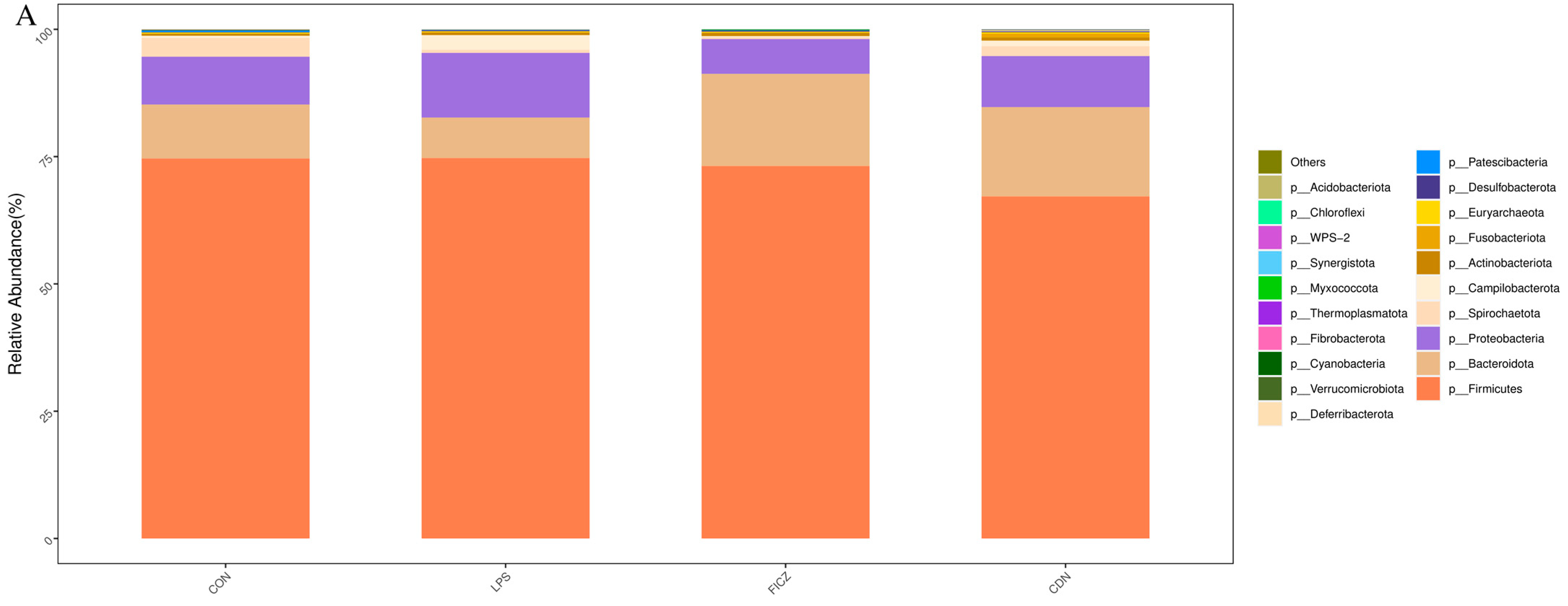
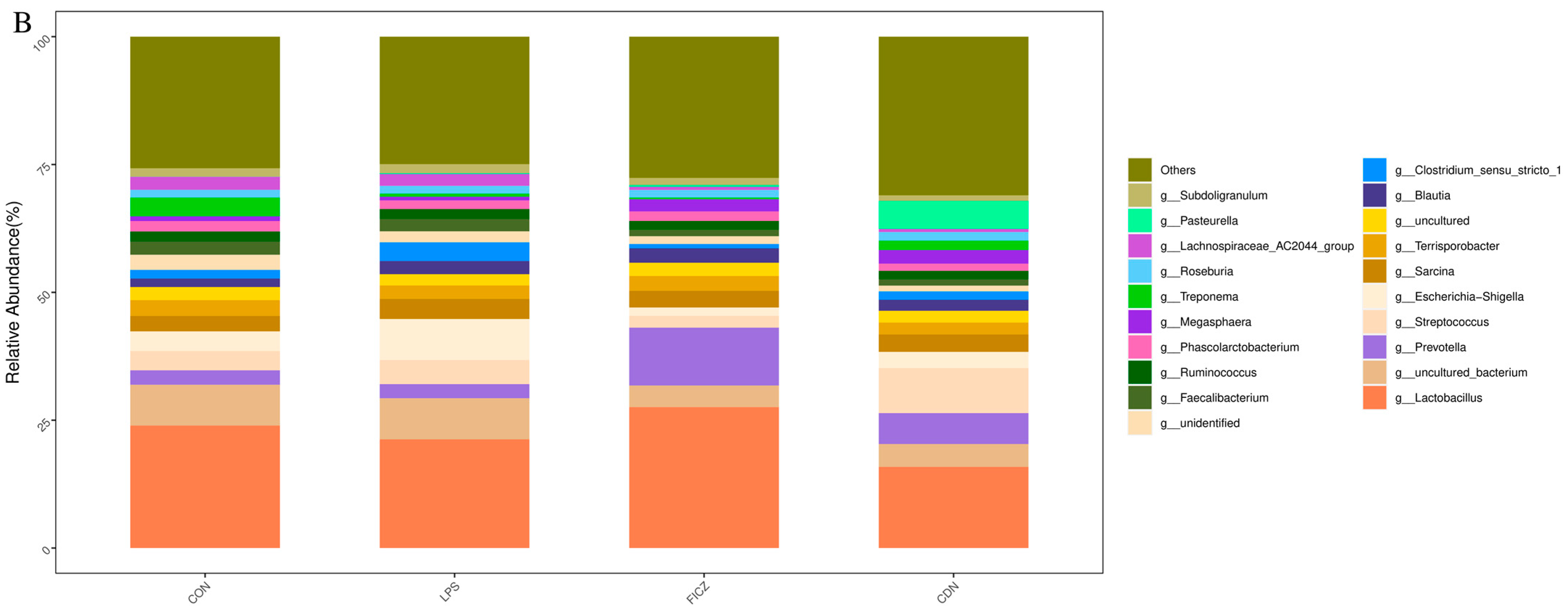
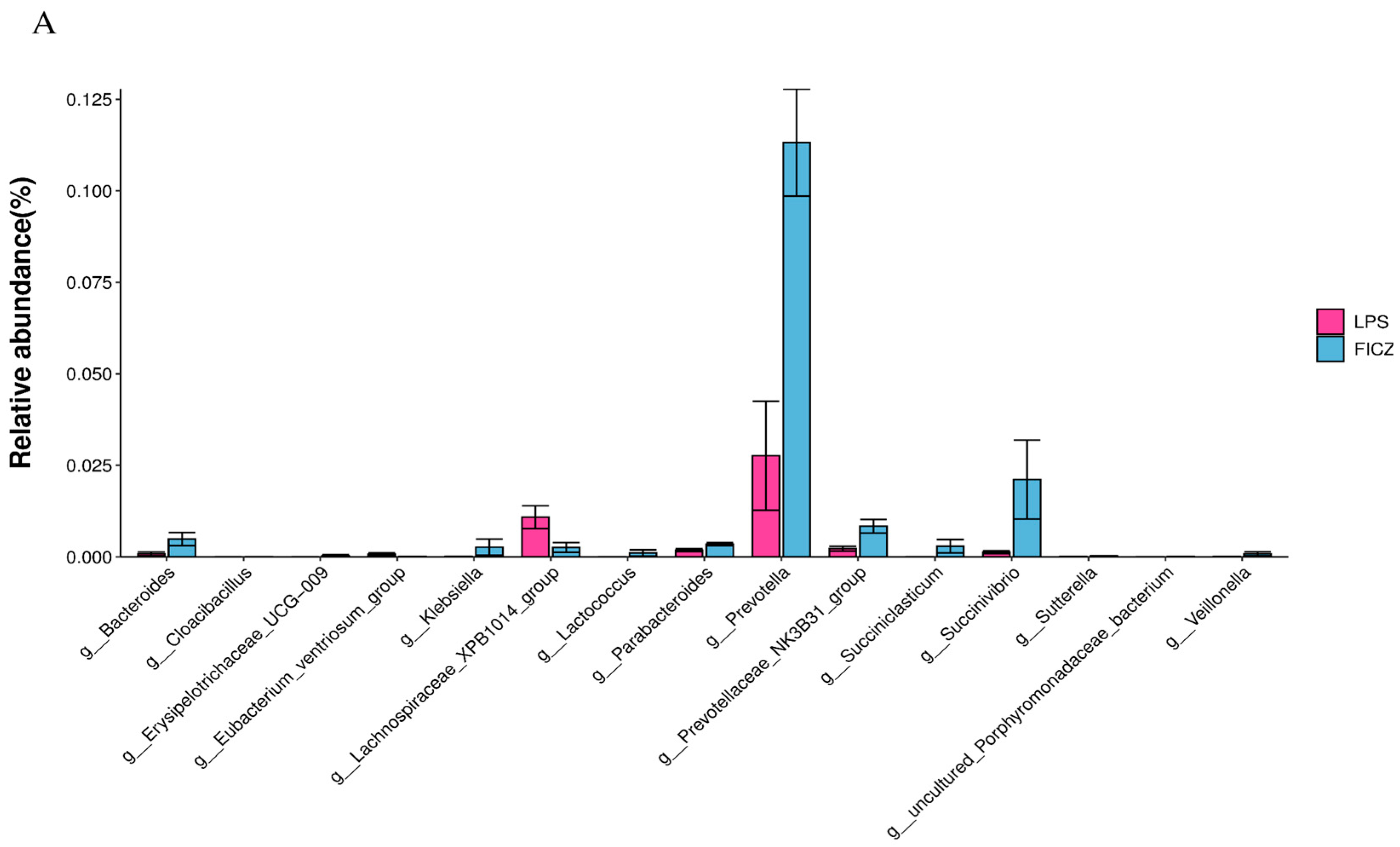
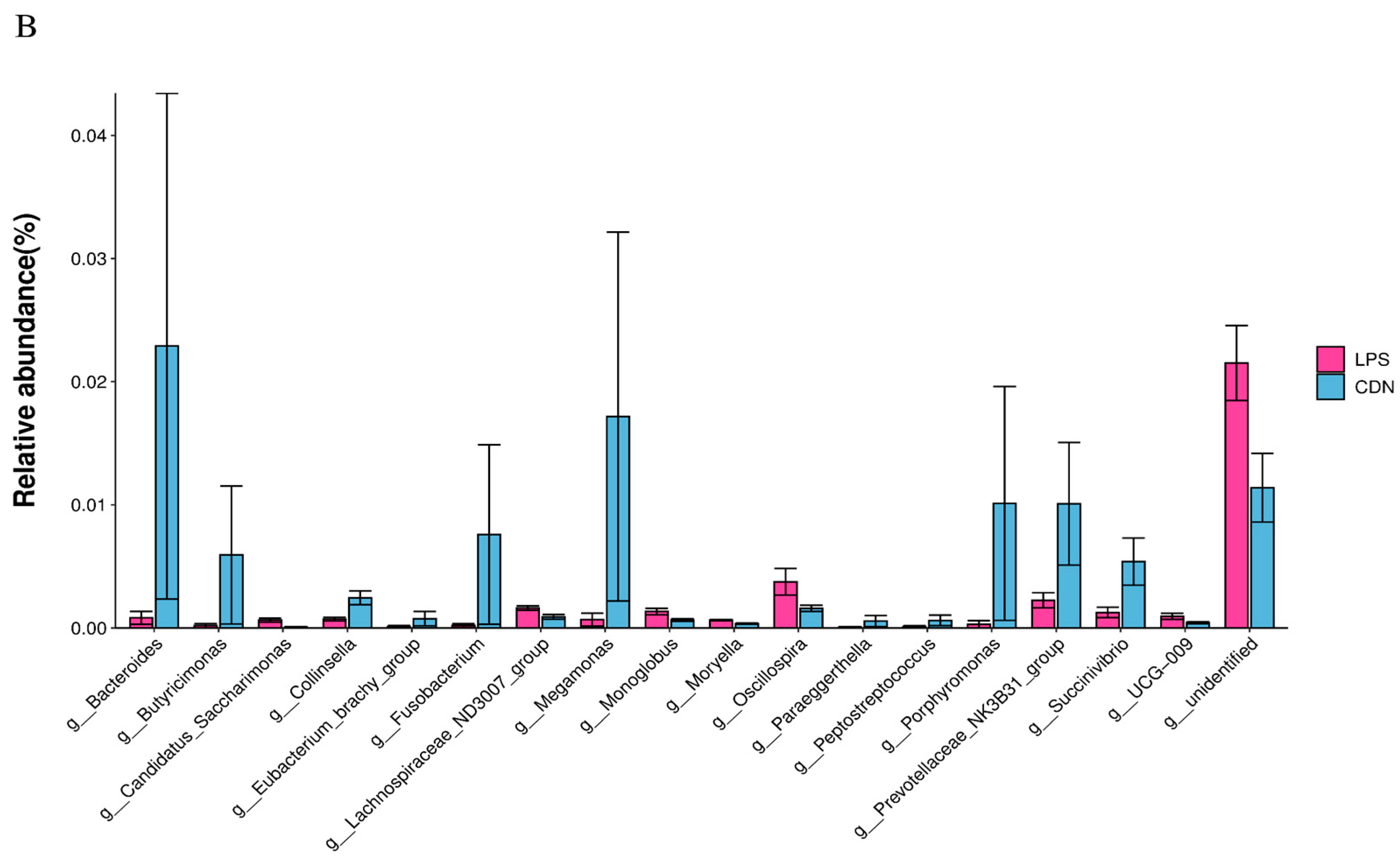
3.4. Gut Microbiota Analysis
3.4.1. Structure and Diversity of Gut Flora
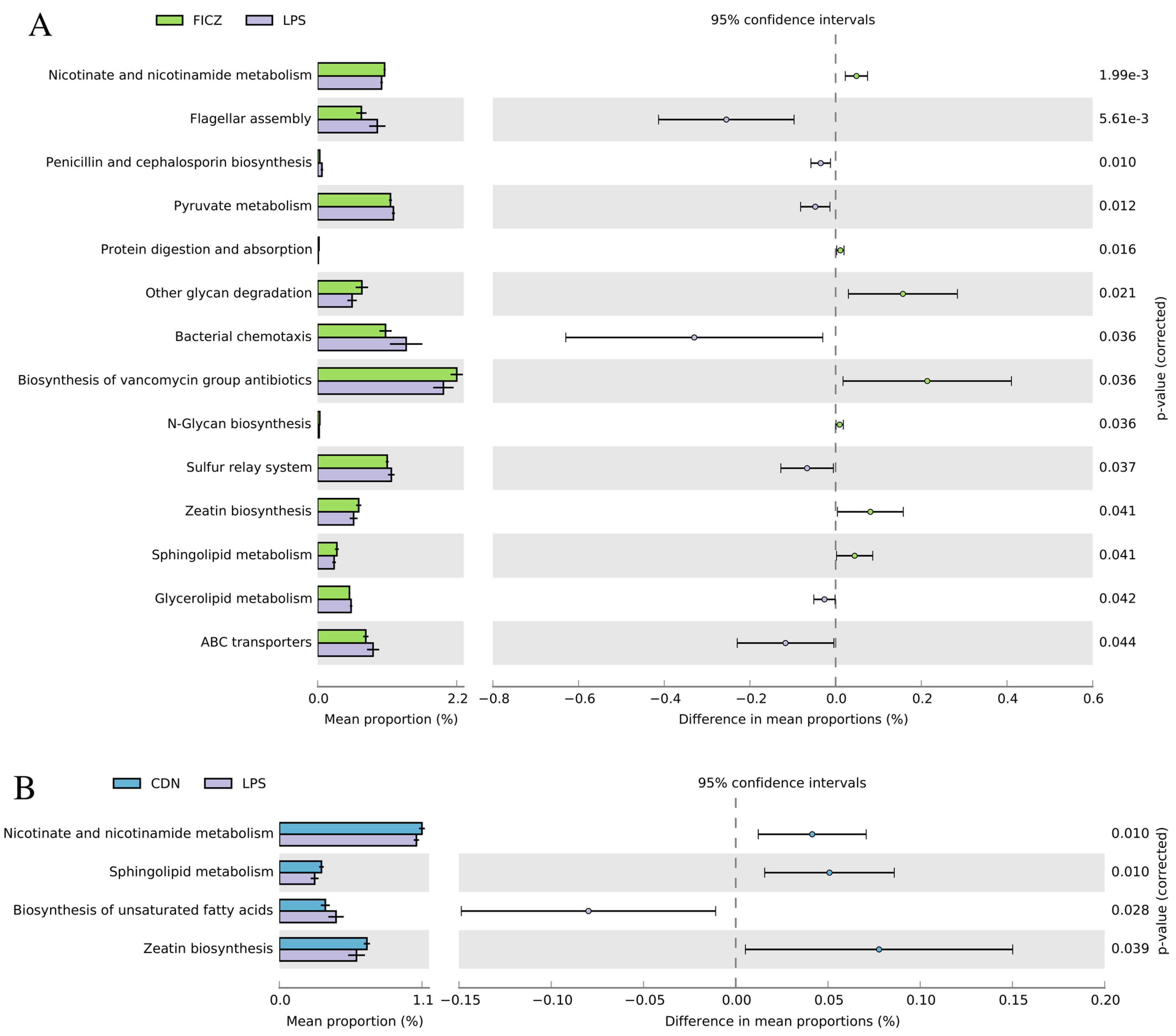
3.4.2. PICRUSt2-Predicted Functional Pathway Analysis
3.5. Short-Chain Fatty Acid (SCFA) Analysis
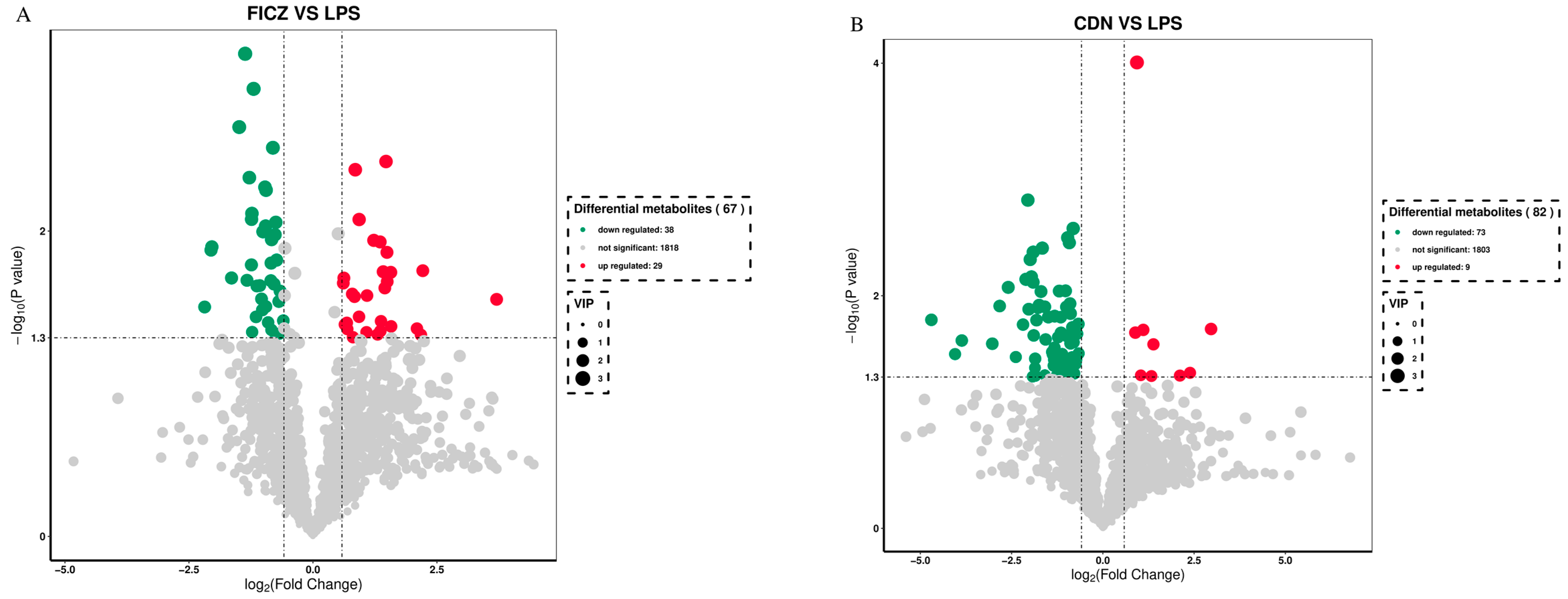
3.6. Metabolomics Analyses
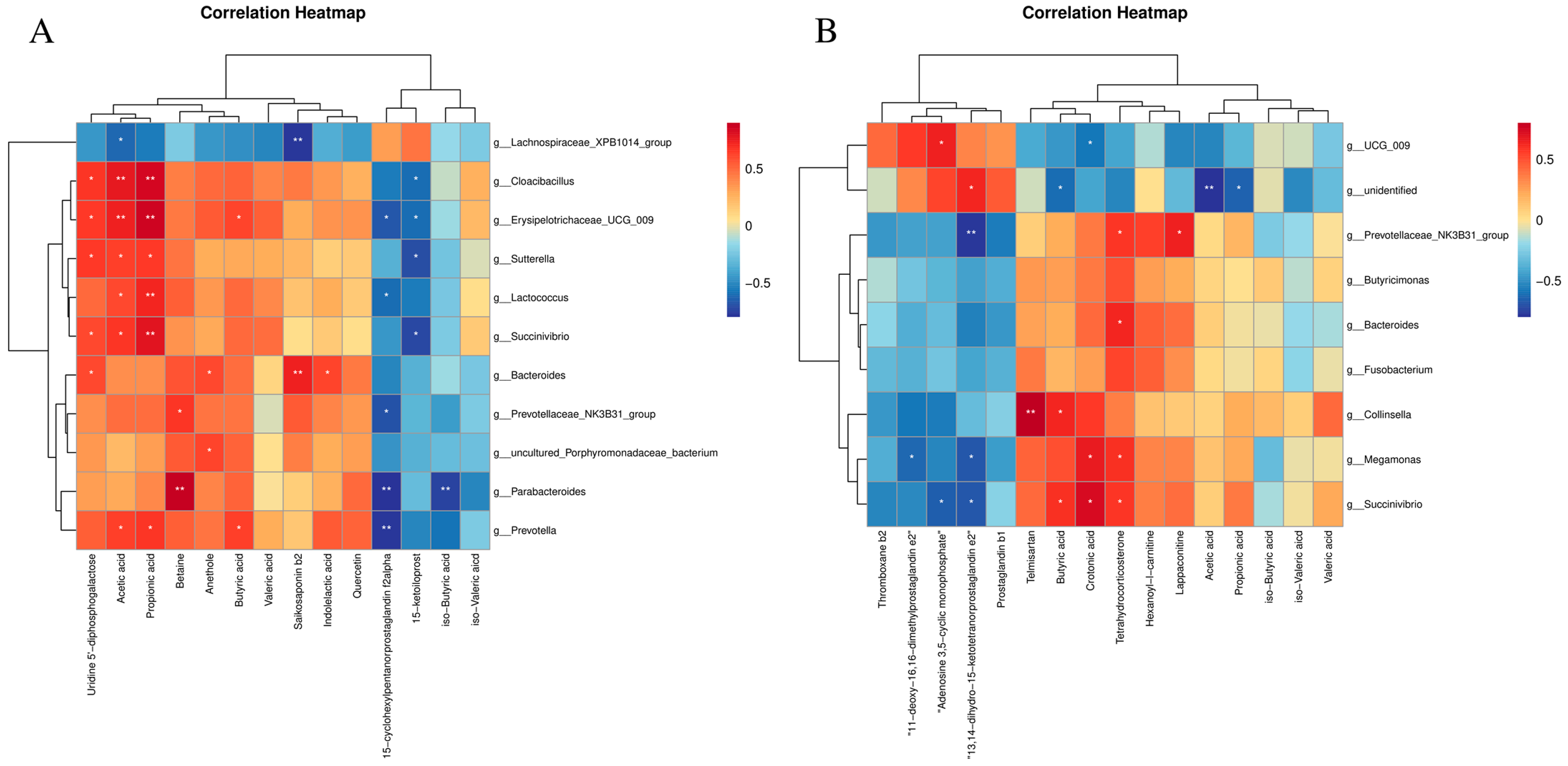
3.7. Correlation Analysis
4. Discussion
4.1. AhR Activation Improves Ileal Mucosa Morphology in Immune-Stressed Piglets
4.2. AhR Activation Reduces Intestinal Barrier Damage in Immune-Stressed Piglets
4.3. AhR Activation Reduces mRNA Expression of Cytokine in Immune-Stressed Piglets
4.4. AhR Activation Improved Intestinal Microbial Diversity and Short-Chain Fatty Acid Content in Piglets under Immune Stress
4.5. AhR Activation Regulates the Metabolome in Immune-Stressed Piglets
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Su, W.; Gong, T.; Jiang, Z.; Lu, Z.; Wang, Y. The Role of Probiotics in Alleviating Postweaning Diarrhea in Piglets from the Perspective of Intestinal Barriers. Front. Cell. Infect. Microbiol. 2022, 12, 883107. [Google Scholar] [CrossRef]
- Tang, X.; Xiong, K.; Fang, R.; Li, M. Weaning stress and intestinal health of piglets: A review. Front. Immunol. 2022, 13, 1042778. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Li, X.G.; Kong, X.; Huang, R.; Ruan, Z.; Yao, K.; Deng, Z.; Xie, M.; Shinzato, I.; Yin, Y.; et al. Dietary L-arginine supplementation enhances the immune status in early-weaned piglets. Amino Acids 2009, 37, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Modina, S.C.; Polito, U.; Rossi, R.; Corino, C.; Di Giancamillo, A. Nutritional Regulation of Gut Barrier Integrity in Weaning Piglets. Anim. Open Access J. 2019, 9, 1045. [Google Scholar] [CrossRef]
- Freudenberg, M.A.; Tchaptchet, S.; Keck, S.; Fejer, G.; Huber, M.; Schütze, N.; Beutler, B.; Galanos, C. Lipopolysaccharide sensing an important factor in the innate immune response to Gram-negative bacterial infections: Benefits and hazards of LPS hypersensitivity. Immunobiology 2008, 213, 193–203. [Google Scholar] [CrossRef]
- Han, Y.; Tang, C.; Zhao, Q.; Fan, S.; Yang, P.; Zhang, J. Butyrate Mitigates Lipopolysaccharide-Induced Intestinal Morphological Changes in Weanling Piglets by Regulating the Microbiota and Energy Metabolism, and Alleviating Inflammation and Apoptosis. Microorganisms 2022, 10, 2001. [Google Scholar] [CrossRef] [PubMed]
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Stockinger, B.; Shah, K.; Wincent, E. AHR in the intestinal microenvironment: Safeguarding barrier function. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 559–570. [Google Scholar] [CrossRef]
- Dinallo, V.; Marafini, I.; Di Fusco, D.; Di Grazia, A.; Laudisi, F.; Dwairi, R.; Paoluzi, O.A.; Monteleone, G.; Monteleone, I. Protective Effects of Aryl Hydrocarbon Receptor Signaling in Celiac Disease Mucosa and in Poly I:C-Induced Small Intestinal Atrophy Mouse Model. Front. Immunol. 2019, 10, 91. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, Q.; Yu, K.; Fan, X.; Xiao, W.; Cai, Y.; Xu, P.; Yu, M.; Yang, H. 6-Formylindolo(3,2-b)carbazole induced aryl hydrocarbon receptor activation prevents intestinal barrier dysfunction through regulation of claudin-2 expression. Chem.-Biol. Interact. 2018, 288, 83–90. [Google Scholar] [CrossRef]
- Yu, M.; Wang, Q.; Ma, Y.; Li, L.; Yu, K.; Zhang, Z.; Chen, G.; Li, X.; Xiao, W.; Xu, P.; et al. Aryl Hydrocarbon Receptor Activation Modulates Intestinal Epithelial Barrier Function by Maintaining Tight Junction Integrity. Int. J. Biol. Sci. 2018, 14, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Yang, K.; Zhou, C.; Xu, P.; Xiao, W.; Yang, H. Aryl hydrocarbon receptor activation alleviates dextran sodium sulfate-induced colitis through enhancing the differentiation of goblet cells. Biochem. Biophys. Res. Commun. 2019, 514, 180–186. [Google Scholar] [CrossRef]
- Liu, Z.; Li, L.; Chen, W.; Wang, Q.; Xiao, W.; Ma, Y.; Sheng, B.; Li, X.; Sun, L.; Yu, M.; et al. Aryl hydrocarbon receptor activation maintained the intestinal epithelial barrier function through Notch1 dependent signaling pathway. Int. J. Mol. Med. 2018, 41, 1560–1572. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Lv, Q.; Miao, Y.M.; Qiao, S.M.; Dai, Y.; Wei, Z.F. Cardamonin, a natural flavone, alleviates inflammatory bowel disease by the inhibition of NLRP3 inflammasome activation via an AhR/Nrf2/NQO1 pathway. Biochem. Pharmacol. 2018, 155, 494–509. [Google Scholar] [CrossRef]
- Hou, S.; Yuan, Q.; Yu, N.; Liu, B.; Huang, G.; Yuan, X. Cardamonin attenuates chronic inflammation and tumorigenesis in colon. Cell Cycle 2019, 18, 3275–3287. [Google Scholar] [CrossRef]
- Barber, K.; Mendonca, P.; Soliman, K.F.A. The Neuroprotective Effects and Therapeutic Potential of the Chalcone Cardamonin for Alzheimer’s Disease. Brain Sci. 2023, 13, 145. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xiao, K.; Yu, C.; Wang, L.; Liang, T.; Zhu, H.; Xu, X.; Liu, Y. Xylooligosaccharide attenuates lipopolysaccharide-induced intestinal injury in piglets via suppressing inflammation and modulating cecal microbial communities. Anim. Nutr. 2021, 7, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, H.; Wang, S.; Tu, Z.; Zhang, L.; Wang, X.; Hou, Y.; Wang, C.; Chen, J.; Liu, Y. Flaxseed Oil Attenuates Intestinal Damage and Inflammation by Regulating Necroptosis and TLR4/NOD Signaling Pathways Following Lipopolysaccharide Challenge in a Piglet Model. Mol. Nutr. Food Res. 2018, 62, e1700814. [Google Scholar] [CrossRef]
- Schiering, C.; Wincent, E.; Metidji, A.; Iseppon, A.; Li, Y.; Potocnik, A.J.; Omenetti, S.; Henderson, C.J.; Wolf, C.R.; Nebert, D.W.; et al. Feedback control of AHR signalling regulates intestinal immunity. Nature 2017, 542, 242–245. [Google Scholar] [CrossRef]
- Shen, J.; Yang, L.; You, K.; Chen, T.; Su, Z.; Cui, Z.; Wang, M.; Zhang, W.; Liu, B.; Zhou, K.; et al. Indole-3-Acetic Acid Alters Intestinal Microbiota and Alleviates Ankylosing Spondylitis in Mice. Front. Immunol. 2022, 13, 762580. [Google Scholar] [CrossRef]
- Li, X.; Akhtar, S.; Choudhry, M.A. Alteration in intestine tight junction protein phosphorylation and apoptosis is associated with increase in IL-18 levels following alcohol intoxication and burn injury. Biochim. Biophys. Acta 2012, 1822, 196–203. [Google Scholar] [CrossRef]
- Schulzke, J.D.; Ploeger, S.; Amasheh, M.; Fromm, A.; Zeissig, S.; Troeger, H.; Richter, J.; Bojarski, C.; Schumann, M.; Fromm, M. Epithelial tight junctions in intestinal inflammation. Ann. N. Y. Acad. Sci. 2009, 1165, 294–300. [Google Scholar] [CrossRef]
- Vindigni, S.M.; Zisman, T.L.; Suskind, D.L.; Damman, C.J. The intestinal microbiome, barrier function, and immune system in inflammatory bowel disease: A tripartite pathophysiological circuit with implications for new therapeutic directions. Ther. Adv. Gastroenterol. 2016, 9, 606–625. [Google Scholar] [CrossRef]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M.; et al. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef]
- Wang, X.; Yong, C.C.; Oh, S. Metabolites of Latilactobacillus curvatus BYB3 and Indole Activate Aryl Hydrocarbon Receptor to Attenuate Lipopolysaccharide-Induced Intestinal Barrier Dysfunction. Food Sci. Anim. Resour. 2022, 42, 1046–1060. [Google Scholar] [CrossRef]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, inflammation, and metabolic disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Zenewicz, L.A.; Flavell, R.A. IL-22 and inflammation: Leukin’ through a glass onion. Eur. J. Immunol. 2008, 38, 3265–3268. [Google Scholar] [CrossRef] [PubMed]
- Jacobse, J.; Li, J.; Rings, E.; Samsom, J.N.; Goettel, J.A. Intestinal Regulatory T Cells as Specialized Tissue-Restricted Immune Cells in Intestinal Immune Homeostasis and Disease. Front. Immunol. 2021, 12, 716499. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, J.; Rasul, A.; Shah, M.A.; Hussain, G.; Riaz, A.; Sarfraz, I.; Zafar, S.; Adnan, M.; Khan, A.H.; Selamoglu, Z. Cardamonin: A new player to fight cancer via multiple cancer signaling pathways. Life Sci. 2020, 250, 117591. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Cheng, Y.; Fu, J.; Lu, Z.; Wang, F.; Jin, M.; Zong, X.; Wang, Y. Gut Immunity and Microbiota Dysbiosis Are Associated with Altered Bile Acid Metabolism in LPS-Challenged Piglets. Oxidative Med. Cell. Longev. 2021, 2021, 6634821. [Google Scholar] [CrossRef]
- Geng, S.; Cheng, S.; Li, Y.; Wen, Z.; Ma, X.; Jiang, X.; Wang, Y.; Han, X. Faecal Microbiota Transplantation Reduces Susceptibility to Epithelial Injury and Modulates Tryptophan Metabolism of the Microbial Community in a Piglet Model. J. Crohn’s Colitis 2018, 12, 1359–1374. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Zhong, R.; Dang, G.; Xia, B.; Wu, W.; Tang, S.; Tang, L.; Liu, L.; Liu, Z.; Chen, L.; et al. Pectin supplementation ameliorates intestinal epithelial barrier function damage by modulating intestinal microbiota in lipopolysaccharide-challenged piglets. J. Nutr. Biochem. 2022, 109, 109107. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Li, P.; Hao, S.; Zhang, Y.; Kim, S.W.; Li, H.; Ma, X.; Gao, S.; He, L.; Wu, W.; et al. Dynamic distribution of the gut microbiota and the relationship with apparent crude fiber digestibility and growth stages in pigs. Sci. Rep. 2015, 5, 9938. [Google Scholar] [CrossRef] [PubMed]
- Panasevich, M.R.; Wankhade, U.D.; Chintapalli, S.V.; Shankar, K.; Rector, R.S. Cecal versus fecal microbiota in Ossabaw swine and implications for obesity. Physiol. Genom. 2018, 50, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Yu, B.; Yu, J.; Zheng, P.; Huang, Z.; Luo, J.; Mao, X.; He, J.; Yan, H.; Wu, J.; et al. Lower abundance of Bacteroides and metabolic dysfunction are highly associated with the post-weaning diarrhea in piglets. Sci. China Life Sci. 2022, 65, 2062–2075. [Google Scholar] [CrossRef] [PubMed]
- Zafar, H.; Saier, M.H., Jr. Gut Bacteroides species in health and disease. Gut Microbes 2021, 13, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Jiang, A.; Wang, X.; Zhou, Y.; Tang, W.; Ren, C.; Qian, X.; Zhou, Z.; Gong, A. NMN Maintains Intestinal Homeostasis by Regulating the Gut Microbiota. Front. Nutr. 2021, 8, 714604. [Google Scholar] [CrossRef]
- Xun, W.; Ji, M.; Ma, Z.; Deng, T.; Yang, W.; Hou, G.; Shi, L.; Cao, T. Dietary emodin alleviates lipopolysaccharide-induced intestinal mucosal barrier injury by regulating gut microbiota in piglets. Anim. Nutr. 2023, 14, 152–162. [Google Scholar] [CrossRef]
- Lin, T.Y.; Wu, P.H.; Lin, Y.T.; Hung, S.C. Gut dysbiosis and mortality in hemodialysis patients. NPJ Biofilms Microbiomes 2021, 7, 20. [Google Scholar] [CrossRef]
- Modoux, M.; Rolhion, N.; Lefevre, J.H.; Oeuvray, C.; Nádvorník, P.; Illes, P.; Emond, P.; Parc, Y.; Mani, S.; Dvorak, Z.; et al. Butyrate acts through HDAC inhibition to enhance aryl hydrocarbon receptor activation by gut microbiota-derived ligands. Gut Microbes 2022, 14, 2105637. [Google Scholar] [CrossRef]
- Ma, Y.; Bao, Y.; Wang, S.; Li, T.; Chang, X.; Yang, G.; Meng, X. Anti-Inflammation Effects and Potential Mechanism of Saikosaponins by Regulating Nicotinate and Nicotinamide Metabolism and Arachidonic Acid Metabolism. Inflammation 2016, 39, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. Adv. Exp. Med. Biol. 2010, 688, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Lappas, C.M. The plant hormone zeatin riboside inhibits T lymphocyte activity via adenosine A2A receptor activation. Cell. Mol. Immunol. 2015, 12, 107–112. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Flaveny, C.A.; Murray, I.A.; Chiaro, C.R.; Perdew, G.H. Ligand selectivity and gene regulation by the human aryl hydrocarbon receptor in transgenic mice. Mol. Pharmacol. 2009, 75, 1412–1420. [Google Scholar] [CrossRef] [PubMed]
- Riemschneider, S.; Hoffmann, M.; Slanina, U.; Weber, K.; Hauschildt, S.; Lehmann, J. Indol-3-Carbinol and Quercetin Ameliorate Chronic DSS-Induced Colitis in C57BL/6 Mice by AhR-Mediated Anti-Inflammatory Mechanisms. Int. J. Environ. Res. Public Health 2021, 18, 2262. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.S.; Im, H.T.; Lee, K.T. Saikosaponin B2 Suppresses Inflammatory Responses Through IKK/IκBα/NF-κB Signaling Inactivation in LPS-Induced RAW 264.7 Macrophages. Inflammation 2019, 42, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.N.; Zhang, X.X.; Shen, P.; Huang, C.S.; Deng, H.F.; Zhou, L.; Yue, L.X.; Shen, B.Y.; Zhou, W.; Gao, Y. Pinelliae rhizoma alleviated acute lung injury induced by lipopolysaccharide via suppressing endoplasmic reticulum stress-mediated NLRP3 inflammasome. Front. Pharmacol. 2022, 13, 883865. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Qin, X.J.; Li, W.P.; Ma, R.; Wang, T.; Li, Z.Q. Effects of Shu Gan Jian Pi formula on rats with carbon tetrachloride-induced liver fibrosis using serum metabonomics based on gas chromatography-time of flight mass spectrometry. Mol. Med. Rep. 2017, 16, 3901–3909. [Google Scholar] [CrossRef]
- Pang, L.; Liu, C.Y.; Gong, G.H.; Quan, Z.S. Synthesis, in vitro and in vivo biological evaluation of novel lappaconitine derivatives as potential anti-inflammatory agents. Acta Pharm. Sin. B 2020, 10, 628–645. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, S.; Liu, J.; Jin, Y.; Yu, S.; An, R. Telmisartan inhibits oxalate and calcium oxalate crystal-induced epithelial-mesenchymal transformation via PPAR-γ-AKT/STAT3/p38 MAPK-Snail pathway. Life Sci. 2020, 241, 117108. [Google Scholar] [CrossRef]
- Larigot, L.; Juricek, L.; Dairou, J.; Coumoul, X. AhR signaling pathways and regulatory functions. Biochim. Open 2018, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Seidel, S.D.; Winters, G.M.; Rogers, W.J.; Ziccardi, M.H.; Li, V.; Keser, B.; Denison, M.S. Activation of the Ah receptor signaling pathway by prostaglandins. J. Biochem. Mol. Toxicol. 2001, 15, 187–196. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items mg/g | Groups | ||||
|---|---|---|---|---|---|
| CON | LPS | FICZ | LCDN | HCDN | |
| Acetic acid | 3.26 ± 0.16 bc | 2.59 ± 0.11 c | 7.73 ± 0.15 a | 4.35 ± 0.23 bc | 4.98 ± 0.11 b |
| Propionic acid | 2.24 ± 0.15 bc | 1.74 ± 0.07 c | 5.96 ± 0.09 a | 2.86 ± 0.25 b | 2.95 ± 0.16 b |
| iso-Butyric acid | 1.31 ± 0.06 a | 0.15 ± 0.02 b | 0.13 ± 0.02 b | 0.20 ± 0.03 b | 0.19 ± 0.07 b |
| Butyric acid | 1.26 ± 0.03 bc | 0.83 ± 0.04 c | 2.84 ± 0.04 a | 1.44 ± 0.10 bc | 1.78 ± 0.04 b |
| iso-Valeric aicd | 0.14 ± 0.002 b | 0.33 ± 0.06 a | 0.29 ± 0.02 a | 0.42 ± 0.04 a | 0.33 ± 0.01 a |
| Valeric acid | 0.37 ± 0.03 b | 0.28 ± 0.03 b | 1.32 ± 0.04 a | 0.43 ± 0.07 b | 0.41 ± 0.06 b |
| MS2 Metabolite | VIP | p-Value | Regulated | LOG-FOLDCHANGE |
|---|---|---|---|---|
| Dihydrocapsaicin | 2.0112 | 0.021934 | up | 0.61289 |
| Confertifoline | 1.8944 | 0.040915 | up | 0.64403 |
| “6,7,4-trihydroxyisoflavone” | 1.8556 | 0.039896 | up | 0.68289 |
| Chaulmoogric acid | 1.9898 | 0.025845 | up | 0.79199 |
| Oleic acid methyl ester | 1.8356 | 0.049591 | up | 0.80598 |
| “D-lysergic acid n,n-diethylamide” | 1.9823 | 0.026908 | up | 0.83793 |
| Pachymic acid | 2.2856 | 0.0083937 | up | 0.93002 |
| Cholestenone | 1.9460 | 0.026444 | up | 1.0893 |
| Arg-Ile | 1.8466 | 0.047509 | up | 1.3036 |
| Saikosaponin b2 | 2.1335 | 0.011783 | up | 1.355 |
| Nicotinate d-ribonucleotide | 1.8537 | 0.038902 | up | 1.3711 |
| Betaine | 1.9961 | 0.023602 | up | 1.4485 |
| D-ribose 1-phosphate | 2.1018 | 0.013782 | up | 1.4909 |
| Quercetin | 2.1244 | 0.018647 | up | 1.5692 |
| 2-(n-ethyl-n-m-toluidino)ethanol | 1.8608 | 0.043479 | up | 2.0975 |
| Uridine 5-diphosphogalactose | 1.8457 | 0.047818 | up | 2.1773 |
| D-fructose 6-phosphate | 2.0381 | 0.018179 | up | 2.2142 |
| Anethole | 2.0317 | 0.028003 | up | 3.7029 |
| 4-imidazoleacrylic acid | 2.5018 | 0.002083 | down | −1.4865 |
| Phenol | 2.6445 | 0.00069024 | down | −1.3677 |
| 15-ketoiloprost | 2.2770 | 0.010094 | down | −1.0058 |
| 15-cyclohexylpentanorprostaglandin f2alpha | 2.3686 | 0.0051695 | down | −0.96624 |
| 4-imidazoleacrylic acid | 2.5018 | 0.002083 | down | −1.4865 |
| MS2 Metabolite | VIP | p-Value | Regulated | LOG-FOLDCHANGE |
|---|---|---|---|---|
| Pantetheine | 2.0731 | 0.0161 | down | −4.6970 |
| “13,14-dihydro-15-ketotetranorprostaglandin e2” | 1.8806 | 0.0317 | down | −4.0492 |
| Dihydrofolic acid | 2.0272 | 0.0177 | down | −2.1853 |
| “Adenosine 3,5-cyclic monophosphate” | 2.3800 | 0.0072 | down | −2.1068 |
| “2,7,8-trimethyl-2-(beta-carboxyethyl)-6-hydroxychroman” | 2.4622 | 0.0015 | down | −2.0551 |
| Thromboxane b2 | 1.8902 | 0.0349 | down | −1.1719 |
| Dapsone | 1.8302 | 0.0459 | down | −1.1707 |
| “11-deoxy-16,16-dimethylprostaglandin e2” | 2.2233 | 0.0091 | down | −1.0219 |
| 1-palmitoyl-2-lauroyl-sn-glycero-3-phosphorylcholine | 2.4830 | 0.0026 | down | −0.8113 |
| Prostaglandin b1 | 1.9438 | 0.0212 | down | −0.7051 |
| Tetrahydrocorticosterone | 2.0868 | 0.0207 | up | 0.8899 |
| “1-(5-fluoropentyl)-1h-indazole-3-carboxylic acid, 1-naphthalenyl ester” | 2.8076 | 0.0001 | up | 0.9329 |
| Asn-Trp-Arg | 2.0571 | 0.0196 | up | 1.1073 |
| Glu-Asn-Arg | 1.8234 | 0.0491 | up | 1.3299 |
| Lappaconitine | 1.9794 | 0.0262 | up | 1.3863 |
| Hexanoyl-l-carnitine | 1.8998 | 0.0486 | up | 2.1091 |
| Crotonic acid | 1.8925 | 0.0460 | up | 2.3880 |
| Telmisartan | 2.0992 | 0.0193 | up | 2.9631 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Zhang, Y.; Ji, M.; Yang, W.; Deng, T.; Hou, G.; Shi, L.; Xun, W. AhR Activation Ameliorates Intestinal Barrier Damage in Immunostressed Piglets by Regulating Intestinal Flora and Its Metabolism. Animals 2024, 14, 794. https://doi.org/10.3390/ani14050794
Wu X, Zhang Y, Ji M, Yang W, Deng T, Hou G, Shi L, Xun W. AhR Activation Ameliorates Intestinal Barrier Damage in Immunostressed Piglets by Regulating Intestinal Flora and Its Metabolism. Animals. 2024; 14(5):794. https://doi.org/10.3390/ani14050794
Chicago/Turabian StyleWu, Xiaomei, Yalei Zhang, Mengyao Ji, Wen Yang, Tanjie Deng, Guanyu Hou, Liguang Shi, and Wenjuan Xun. 2024. "AhR Activation Ameliorates Intestinal Barrier Damage in Immunostressed Piglets by Regulating Intestinal Flora and Its Metabolism" Animals 14, no. 5: 794. https://doi.org/10.3390/ani14050794
APA StyleWu, X., Zhang, Y., Ji, M., Yang, W., Deng, T., Hou, G., Shi, L., & Xun, W. (2024). AhR Activation Ameliorates Intestinal Barrier Damage in Immunostressed Piglets by Regulating Intestinal Flora and Its Metabolism. Animals, 14(5), 794. https://doi.org/10.3390/ani14050794