Simple Summary
Consumer perception related to health and environmental issues associated with ruminant products, such as greenhouse gas emissions, has led to a paradigm shift aimed at mitigating the potential harmful effects of ruminant production. This study consolidated the current body of research on dietary rumen manipulating strategies with an aim to quantify their impact on rumen fermentation, enteric methane (CH4) emission and productivity by creating a global database on in vivo evaluation studies. A meta-analytical approach was used to achieve the study’s aim and the result showed that nitrate, saponin, oils, biochar and 3-nitroxypropanol (3-NOP) were effective dietary rumen modulating strategies to mitigate enteric CH4 emission. Of these effective strategies, oils and 3-NOP provided a co-benefit in terms of improving productivity in ruminant livestock. Concentrate feeding equally improved production without any significant effect on enteric methane emissions.
Abstract
A meta-analysis was conducted with an aim to quantify the beneficial effects of nine different dietary rumen modulating strategies which includes: the use of plant-based bioactive compounds (saponin, tannins, oils, and ether extract), feed additives (nitrate, biochar, seaweed, and 3-nitroxy propanol), and diet manipulation (concentrate feeding) on rumen fermentation, enteric methane (CH4) production (g/day), CH4 yield (g/kg dry matter intake) and CH4 emission intensity (g/kg meat or milk), and production performance parameters (the average daily gain, milk yield and milk quality) of ruminant livestock. The dataset was constructed by compiling global data from 110 refereed publications on in vivo studies conducted in ruminants from 2005 to 2023 and anlayzed using a meta-analytical approach.. Of these dietary rumen manipulation strategies, saponin and biochar reduced CH4 production on average by 21%. Equally, CH4 yield was reduced by 15% on average in response to nitrate, oils, and 3-nitroxy propanol (3-NOP). In dairy ruminants, nitrate, oils, and 3-NOP reduced the intensity of CH4 emission (CH4 in g/kg milk) on average by 28.7%. Tannins and 3-NOP increased on average ruminal propionate and butyrate while reducing the acetate:propionate (A:P) ratio by 12%, 13.5% and 13%, respectively. Oils increased propionate by 2% while reducing butyrate and the A:P ratio by 2.9% and 3.8%, respectively. Use of 3-NOP increased the production of milk fat (g/kg DMI) by 15% whereas oils improved the yield of milk fat and protein (kg/d) by 16% and 20%, respectively. On the other hand, concentrate feeding improved dry matter intake and milk yield (g/kg DMI) by 23.4% and 19%, respectively. However, feed efficiency was not affected by any of the dietary rumen modulating strategies. Generally, the use of nitrate, saponin, oils, biochar and 3-NOP were effective as CH4 mitigating strategies, and specifically oils and 3-NOP provided a co-benefit of improving production parameters in ruminant livestock. Equally concentrate feeding improved production parameters in ruminant livestock without any significant effect on enteric methane emission. Therefore, it is advisable to refine further these strategies through life cycle assessment or modelling approaches to accurately capture their influence on farm-scale production, profitability and net greenhouse gas emissions. The adoption of the most viable, region-specific strategies should be based on factors such as the availability and cost of the strategy in the region, the specific goals to be achieved, and the cost–benefit ratio associated with implementing these strategies in ruminant livestock production systems.
1. Introduction
Livestock contribute to approximately 14.5% of global agricultural-related greenhouse gas (GHG) emissions [1], with ruminants being responsible for about 80% of these emissions [2]. Enteric methane (CH4) emitted by ruminants from ruminal fermentation accounts for 2–12% of the energy loss from the diets they consume [3]. In order to improve the energy utilisation efficiency of ruminant diets and reduce their CH4 emissions, it is necessary to adopt sustainable dietary strategies that are specifically designed to mitigate CH4 emissions. Several dietary strategies to reduce CH4 emissions in ruminants have been extensively studied and qualitatively reviewed [4,5]. However, the results of these studies have been variable. Therefore, a meta-analytical approach could provide quantified effects of these strategies on enteric CH4 emissions and productivity in ruminants, improving overall understanding. Most of the available meta-analytical studies have only focused on individual strategies at a time [6,7], whereas studies evaluating multiple rumen-modulating strategies were based on observations from a sole ruminant species [8]. However, van Gastelen et al. [9] quantitatively reviewed the effects of several enteric CH4 mitigating strategies on how they behave across different ruminant species and conclusively reported no difference between species when the mode of action directly approaches a methanogenesis pathway. Hence, there is a need to complement this approach by consolidating data from global studies conducted to evaluate different dietary rumen manipulating strategies in ruminants using percentage change data rather than using absolute values, as the approach by previous authors ruled out animal species as an explanatory variable when quantifying effects of methane reducing strategies in relation to the biological influence of methane-related pathways. Moreover, Congio et al. [10] compared different rumen-modulating strategies for ruminant livestock, but their study was limited to the Latin America and Caribbean regions. Thus, there is a need to conduct a quantitative review at a global level in order to identify the dietary rumen modulating strategies that reduce the enteric methane emission and/or those that increase productivity across ruminant livestock systems. Recently, Arndt et al. [11] examined the quantified effects of sizeable dietary CH4 mitigating strategies without accounting for rumen fermentation parameters or the quality of milk produced in response to these strategies. Such limitations hinder the translation of results from these studies due to the need to determine the feasibility of effective CH4 mitigation strategies and their impact on rumen fermentation, feed efficiency and production parameters like milk yield and milk quality. Therefore, the current meta-analysis study was conducted using global in vivo experiments to quantify different dietary rumen modulating strategies such as the use of plant-based bioactive compounds (saponin, tannins, oils, and ether extract), feed additives (nitrate, biochar, seaweed, and 3-nitroxy propanol), and diet manipulation (concentrate feedings) on enteric methane emissions (CH4 production, CH4 yield and CH4 intensity) and the productivity of ruminant livestock.
2. Materials and Methods
2.1. Dataset
To comparatively examine quantified effects of multiple rumen-modulating strategies on enteric methane (CH4) emission metrics such as CH4 production (g/day), CH4 yield (g/kg dry matter intake) and CH4 intensity (g/kg meat or milk), dry matter intake, total tract digestibility, ruminal fermentation parameters, milk yield, liveweight gain, and feed efficiency of ruminants, we firstly identified strategies with modes of action associated with methane formation-related pathways. A key rationale behind this approach stems from the notion that if the mode of action of a dietary methane mitigation strategy is closely tied to methanogenesis-related fermentation pathways, it is likely to yield consistent outcomes across diverse ruminant types [9]. Thereafter, the dataset was created by searching online internet sources (i.e., google scholar, web of science, semantic scholar and science direct) for peer-reviewed articles, conferences and theses using the search terms: in vivo, rumen modulating strategies (tannin, saponin, ether extract, nitrate, concentrate feeding, oils, biochar, seaweed, and 3-nitroxy propanol), ruminants (cattle: dairy or beef, buffaloes, goats, sheep, and deer), methane production or emission, and production performance (dry matter intake, total tract digestibility, average daily gain, milk production and yield, and feed conversion ratio) in different combinations. It is important to note that the data on average daily gain and feed conversion ratio (an index of feed efficiency) were gathered from studies reporting the growth performance of beef cattle, buffaloes, deer, sheep, and goats, excluding dairy ruminants. Similarly, feed efficiency in dairy ruminants, measured in grams of milk produced per kilogram of dry matter intake (g/kg DMI), was derived from studies specifically conducted to evaluate milk production data in dairy ruminants. Additionally, ether extract (EE) is the crude fat containing other compounds extracted by diethyl or petroleum ether prior to hydrolysis. The addition of EE as an explanatory variable in mathematical models that predict CH4 emission from ruminants improved the accuracy of the predictions [12]. Hence, EE was evaluated as a separate strategy from the oil addition strategy in this current study. The titles and abstracts of the publications were used as the first screening approach to identify the suitability of the publications for inclusion in the database. Studies were further assessed if they reported experiments conducted using ruminants, evaluated the CH4 mitigating strategies in question, and whether the treatments within each study were clearly defined for inclusion in the database. Publications included in the final dataset as depicted in, met the following criteria: (1) studies should have been written in English; (2) studies should have reported in vivo experiments; (3) methane emissions data reported were measured (e.g., sf6: sulphur hexafluoride) and not estimated; (4) production performance data were reported; (5) for any variable of interest, studies had to have reported the mean and the measure of variability such as the standard error of the mean or the reported measure of variability allows the calculation of the standard error of the mean; (6) studies should have reported the chemical composition per treatment (Figure 1). The dataset included 107 studies published from 2005 to 2023, conducted in 27 countries across six continents viz. Africa, Asia, Australia, Europe, North America, and South America. Studies including more than one methane mitigating strategy as treatments (e.g., factorial experiments) were counted as many times as the variation shown for rumen modulating strategy classification criteria. For instance, studies that compared different strategies and their combinations (interaction) were counted twice, and the data from the interaction effects were not included in the dataset. Sources of saponins encompassed tea saponin, Linum usitatissimum, Medicago sativa, Garcinia mongostana (peel powder), Macrotyloma uniflorum, Camellia sinensis, Gliricidia sepium, Enterolobium cyclocarpum, Pyrus salicifolia, Yucca schidigera, and Quillaja saponaria. Tannin sources included Varchelia tortilis, Terminalia chebula, Allium sativa, Azodirahta indica, Artcarpus heterophyllus, Ficus benghalens, Corylus avellana, Kobe lespedeza, Garcinia mangostana, Cymbopogon citratus, Matricaria chamomilla, Sercea lespedeza, Cosmos bipinnatus, hydrolysable tannin extract, Camellia sinensis, condensed tannin extract, Gliricidia sepium, Enterolobium cyclocarpum, and Schinopsis lorentzii. Oils were derived from Zea mays, Allium sativa, Helianthus annus, Linum usitatissimum, Brassica napus, Menthae piperitae aetheroleumm and Trachyspermum ammi. Ether extract was from crude and extruded Linum usitatissimum, Oryza sativa straw, Garcinia mangostana peel, Camellia sinensis, Digitaria eriantha, Punica granatum, Madhuca longifolia, Tecomella undulata, Elaeis oleifera meal, and Cuminum cyminum. Biochar sources included Pinus Sylvetris, Gallus gallus domesticus, (droppings), Bambusa vulgaris, Oryza sativa (husks), Pistachia vera (by-product), Junglus (shell). Seaweed was derived from Ascophyllum nodosum, Asparagopsis taxiformis, Sargassum wightii, and Rhodopyta.
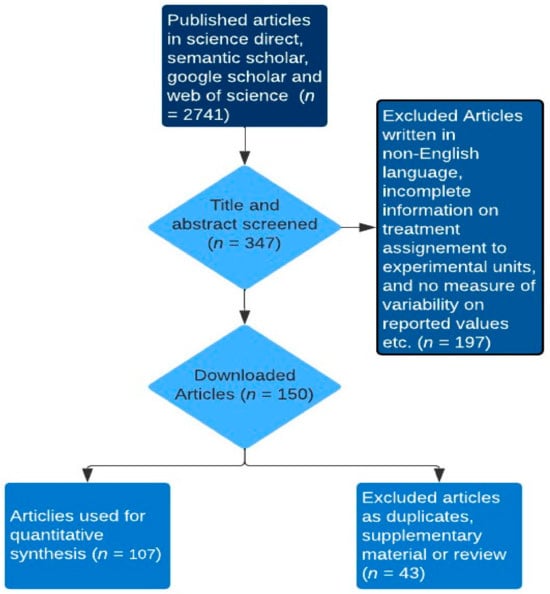
Figure 1.
Flowchart of article selection.
2.2. Calculations and Statistical Analysis
The level of the response variables varied greatly across studies using different types of ruminants, making comparisons on absolute values across studies impractical. Comparison using the mean effect size approach promoted feasibility and facilitated the examination of dietary rumen modulating strategies using data from different types of ruminants. The influence (mean effect size, %) of the dietary rumen modulating strategies on any response variable was calculated as the difference between the rumen modulating strategy (mitigating treatment) and the respective control treatment or baseline, divided by the control treatment, and multiplied by hundred (100) for percentage representation [13], as follows:
The dataset was subjected to the mixed model procedures of SAS (9.4 SAS Inc., Cary, NC, USA) and different studies were considered as random effects. The initial body weight of animals and day of lactation in dairy ruminants were considered as covariates. The dietary rumen modulating strategies, namely saponin, essential oils, tannins, nitrate, concentrate feeding and ether extract, seaweed, and 3-nitroxypropanol, were considered as fixed effects. To account for the differences arising from studies used in the current evaluation, the inverse of the squared standard error of the mean was used as the weighting factor in the “weight” statement of the model as outlined by St-Pierre [14]. The statistical significance was declared at p < 0.05.
3. Results
Table 1 presents summary statistics of the refined complete dataset on dietary rumen modulating strategies, initial body weight, dietary composition, intake, digestibility, average daily gain, milk parameters, methane and fermentation parameters. The skewness and kurtosis represent the degree of symmetry and distribution around the means; either the peakedness or flatness and distribution of data points in comparison to the normal distribution of the refined dataset [15,16]. The data presented in Table 1 failed to achieve homogeneity of variance and none of the data transformation approaches were employed because the mean effect size and weighting approaches described under Section 2.2 are robust against the violation of this assumption [17]. It is worth mentioning that some studies did not report all the variables of interest for the present study, making the number of observations on the variables presented to be inconsistent (Table 1). Some of the dietary rumen modulating strategies evaluated in the current study either reduced enteric methane (CH4) emissions while maintaining or improving productivity of ruminant livestock as reported in Table 2 and Table 3. The mean effect size (%) of evaluated dietary rumen modulating strategies on enteric CH4 production, CH4 yield and CH4 emission intensity, and production performance parameters (average daily gain, milk yield and milk quality) in ruminant livestock is presented in Table 4 and Table 5, respectively, to facilitate the interpretation of results. The information on studies used in the current analysis is reported in Appendix A. Tannins reduced (p < 0.05) total tract digestibility (−12%) and CH4 production (g/day) and CH4 yield (g/kg DMI) by 37% and 40%, respectively. Concentrate feeding improved (p < 0.05) dry matter intake (kg/day) and milk yield (g/kg DMI) on average by 23.41% and 20%, respectively, in ruminant livestock. On average, nitrate, saponin, oils, ether extract and biochar had no effect (p > 0.05) on dry matter intake (DMI), milk yield (MY), and average daily gain (ADG: beef cattle, buffaloes, deer, sheep, and goats). Equally, nitrate and ether extract had no effect (p > 0.05) on CH4liveweight (g/kg liveweight gain) and CH4 production. Seaweed induced a decrease in ADG (p < 0.05) by 3.75%. Methane yield (g/kg DMI) was reduced (p < 0.05) in response to nitrate (−10.11%), tannin (−37%), oils (−7.12-%), seaweed (−35.34%), and 3-nitroxypropanol (3-NOP: −27.36%). Equally, methane production (g/day) was reduced (p < 0.05) in response to the use of saponin (−37.27%), tannins (−40%), biochar (−5.45%), and seaweed (−21.80%) in ruminant livestock. Nitrate, oils and 3-NOP reduced CH4milk (g/kg milk) by 16.63%, 38.96% and 30.46%, respectively. Rumen fermentation parameters (i.e., propionate, butyrate, and A:P ratio) were influenced by the presence of tannins, oils and 3-NOP in diets of ruminant livestock. Tannins increased ruminal propionate (+10.90%) and butyrate (+9.80%) while reducing the A:P ratio (−5.70%). Similarly, 3-NOP increased propionate (+13.10%) and butyrate (+17.26%) with a reduction in the A:P ratio (−20.26%). In contrast, oils increased propionate (+2%) and reduced butyrate (−2.9%) and the A:P ratio (−3.80%) in rumen fermentation parameters. The production of milk fat (g/kg DMI) was increased in response to 3-NOP by 15%, whereas oils improved the yield of milk fat and protein (kg/d) by 16% and 20%, respectively.

Table 1.
Summary statistics of the refined complete dataset on dietary rumen modulating strategies, initial body weight, dietary composition, intake, total tract digestibility, average daily gain, milk parameters, feed conversion ratio, methane and fermentation parameters of ruminant livestock.
Table 2.
Effect of dietary rumen modulating strategies on enteric methane and rumen fermentation parameters from ruminant livestock.
Table 3.
Effect of dietary rumen modulating strategies on production performance and milk quality parameters of ruminant livestock.
Table 4.
Mean effect size (%) of potential dietary rumen modulating strategies on enteric methane emission and rumen fermentation parameters in ruminant livestock.
Table 5.
Mean effect size (%) of potential dietary rumen modulating strategies on production performance parameters in ruminant livestock systems.
4. Discussion
The enormous research interest in adopting a variety of strategies aimed to curb enteric CH4 emissions to reduce the environmental impact of ruminant livestock has yielded dynamic yet attainable results. However, these strategies have been reported to be effective in most in vitro trials and when verified in vivo, few have proved to impact CH4 emission. Also, the vast array of strategies awarding success of mitigating enteric CH4 emissions in ruminants are allied with detrimental impacts on the productivity of animals [4]. Thus, it is of prime importance to collectively examine the existing strategies with the aim to better understand their mode of action and facilitate the creation or meticulous attainment of feasible solutions towards circumventing CH4 emission while improving productivity and feed efficiency in ruminants. The current analysis identified strategies with the mode of action associated with methane formation-related pathways to single out mechanisms to explain the percentage mean effect size of different strategies on CH4 emission, rather than utilising a multi-faceted approach of potential drivers to discuss the results. This is due to their mode of action being similar across different types of ruminant species [9], facilitating the discussion of current findings using quantitative comparative examination.
4.1. Nitrate
Nitrate supply to ruminants in a meta-analysis study by Almedia et al. [18] revealed a 15% reduction in methane yield (g/kg DMI) without impairing total tract digestibility. Congruently, our findings revealed that nitrate reduced methane yield by 10.11% with no adverse effect on dry matter intake and total tract digestibility in ruminants. The reduction in CH4 emission intensity (g/kg milk) was 16% in our meta-analysis which is comparable to the study by Almedia et al. [18], who reported a range of reduction from 10.7% to 18.7%. The mechanism by which nitrate may reduce enteric CH4 emission is via outcompeting methanogenesis pathways for metabolic hydrogen (H2) resulting from the enteric microbial fermentation. This is due to the high affinity of nitrate with H2 as compared to carbon dioxide in the rumen [19]. Nitrate does not adversely affect dry matter intake and total tract digestibility, while it provides (to a minor extent) non-protein nitrogen (N) to the microbiota in the rumen [20]. Also, in developing countries, livestock are mostly maintained in roughage-based diets with little or no dietary non-protein nitrogen supplementation [21]. Therefore, nitrate supply can be a beneficial strategy to reduce CH4 emissions and provide a non-protein nitrogen source in livestock systems that rely on pastures that are inherently low in nitrogen content, especially in the tropics during the dry season.
4.2. Saponin
Although there were no effects of saponins on production performance parameters and other CH4 metrics (i.e., CH4 yield and CH4 per liveweight gain), our results revealed a 37% reduction in CH4 production due to saponins in ruminant livestock. In agreement with our results, a previous study by Abarghuei et al. [22] reported a similar reduction (37%) in CH4 production. Also, when Samane saman (containing saponins) was supplemented to steers fed a basal diet of rice treated with urea, there was a reduction in CH4 production of 50% [23]. However, Samane saman also contains tannins, therefore, its CH4 mitigating effect could be attributed to some unknown extent to other components rather than solely saponins. The mechanism of action in saponins to reduce CH4 production is through the detergent-like action interfering with the integrity of methanogenic archaea cell membranes [24]. In this view, the population and ability of methanogenic archaea to produce CH4 may be compromised and enteric fermentation pathways altered by redirecting more hydrogen towards alternative sinks. The literature reporting the impact of saponin associated protozoa influencing methanogens is intriguing, yet limited studies hinder a conclusive understanding of the phenomenon.
4.3. Tannin
The study conducted by Goel and Makkar [21] compared the effectiveness of tannins and saponins on reducing CH4 production. Their findings indicated that tannins were less effective than saponins, which is contrary to the results obtained in our study (−40% vs. −37%). However, our analysis demonstrated that tannins highly (>30%) reduce both CH4 production and CH4 yield by 40% and 37%, respectively, despite the 12% decrease in total tract digestibility. Similarly, Arndt et al. [11] reported a 12% decrease in both CH4 production and total tract digestibility. Goel and Makkar [21] noted that although tannins do reduce CH4 production in vivo, there is a drawback in terms of impaired animal productivity in view of the reduction in total tract digestibility. Additionally, in this study, fermentation parameters such as propionate and butyrate increased by 10.9% and 9.8%, respectively, while the acetate to propionate ratio decreased by 5.7%. The mechanism by which tannins affect methanogenesis can be partially explained by their direct toxic effect on methanogens and protozoa, as well as their ability to bind to polysaccharides, thereby adversely affecting fibre degradation in the rumen and increasing propionate production [25,26,27]. The reduction in the amount of dry matter degraded in the rumen leads to a decrease in the production of metabolic hydrogen, which is necessary for methanogenesis. Moreover, tannins directly reduce the population of methanogens through their toxic effect and by increasing rumen washout rates, resulting in a reduction of CH4 formation in the rumen [28]. To optimize feed utilization while using condensed tannins in ruminant diets, careful attention must be paid to protein concentration, ensuring it does not compromise real apparent total digestibility. Additionally, research suggests that the effectiveness of tannins varies with their origin, as some rumen microbes develop resistance mechanisms over time [29]. Tannins also form complexes with proteins under physiological ruminal pH conditions, which reduces the degradability of protein in the rumen, thereby improving bypass protein [21].
Although it is not evaluated in the current analysis, it is important to note that the shift of nitrogen excretion from urine to faeces attributed to tannin supply is significant and cannot be overlooked, despite the induced low dry matter digestibility. Therefore, it is necessary to evaluate the long-term effects of tannins on the overall productivity of ruminants [5]. This evaluation should be accompanied by determining the threshold dosage and source of tannins supplied in separate pulse doses, using a range of carriers in addition to the general approach of including it in a total mixed ration. These carriers should be tailored to the specific feeding systems prevalent in different regions; for instance, grazing is the most common system of rearing ruminants in sub-Saharan Africa [30]. Therefore, the carriers should be situation specific or should cater for conditions prevalent in a specific area, or, if possible, a range of areas.
4.4. Oils and Ether Extract
Both oils and ether extract contain polyunsaturated fatty acids that have been found to exhibit anti-methanogenic properties. Therefore, the combination of oils and ether extract facilitate the discussion of the results according to their mode of action in reducing CH4 emissions from ruminants. The findings of the current study revealed several interesting results regarding the effects of oils on ruminants and their milk production. The supply of oils to ruminants led to a reduction of 7.13% and 38.96% in CH4 yield and CH4 intensity (g/kg milk), respectively. Additionally, there was an increase of 16% and 20% in milk fat and protein production (kg) per day, respectively. These results are consistent with a study conducted by Madhavi et al. [31], which reported similar increases of 9.09% and 9.38% in milk fat and protein production, respectively, in response to a supply of oil in the diets of dairy cows. The reduction in CH4 intensity is due to an increase in the production of milk, thereby lowering the quantity of CH4 per unit of milk produced. This is attributed to the biohydrogenation process triggered by the rise in dietary energy from oils in the rumen, where hydrogens are captured and, in the case of dairy ruminants, utilised for the synthesis of milk fat. However, the mechanism behind the potential increase in milk protein remains not clearly known. Generally, it is believed that incorporating feedstuffs such as grains, fats, or oils in ruminant diets can stimulate milk production by providing high energy and redirecting additional energy from CH4 mitigation. The spared energy from inhibiting CH4 in livestock, even with a 25% reduction through feed additives, rarely yields significant improvements in production [32]. Theoretical calculations based on experimental data suggest a minimal and challenging-to-detect enhancement in net energy for production [32]. Despite the anticipated impact on animal energy balance, the observed gains in net energy are biologically insignificant at commonly reported inhibition rates. Methane reduction is linked to changes in organic matter digestibility and apparent digestible energy, complicating its relationship with energy metabolism. Studies reveal that targeting rumen methanogenesis does not consistently boost milk or growth productivity, with confounding factors like intake, digestibility, and rumen pH [33]. In essence, while there is theoretical premise, empirical evidence indicates limited and inconsistent effects on livestock production [34].
As expected, the findings of our study align with a meta-analysis conducted by Villar et al. [35] that examined effects of oil supplementation on dairy cows. The authors reported an 8.8% reduction in CH4 production, supporting the notion that oils can be an effective strategy for mitigating methane emissions. Benchaar et al. [36] reported a 25% reduction in the production of a gram of CH4 per kg of milk produced in lactating dairy cows, which is less than the reported 38.96% from our meta-analysis. However, Martin et al. [37] reported a slightly comparable 30% reduction in CH4 intensity in response to the oil supply in lactating dairy cows. The study of Beauchemin et al. [38] reported a similar observation of reduced CH4 intensity when cows were subjected to oils. The mechanism behind this reduction is believed to involve the disruption of methanogen cell membranes by the direct toxic effect of oils, leading to a decrease in methane production in the rumen. Briefly, polyunsaturated fatty acids in oils can undergo oxidation processes in the rumen. This oxidation can result in the generation of reactive oxygen species within the microbial cells causing oxidative stress, damaging cellular components and leading to cell death in ruminal microbes [39]. This disruption impacts their ability to carry out metabolic processes such as methane production.
On the other hand, our study did not find any significant impact of ether extract on both enteric methane production and animal performance parameters. This differs from the findings of a study by Giger-Reverdin et al. [40], which showed a reduction in CH4 yield in dairy cows fed diets with increased ether extract content. This reduction was attributed to the presence of unsaturated fatty acids (UFAs) in the ether extract, which promote the production of propionate and inhibit cellulolytic activity, thereby reducing methane production. The disparity between our results and those of Giger-Reverdin et al. [40] may be attributed to differences in the sources of ether extracts used in the respective studies, as well as variations in the average ether extract content of the diets. Our study had a lower average ether extract content than the study by Giger-Reverdin et al. [40], which could have contributed to the observed differences.
4.5. Concentrate Feeding
Dietary manipulation through the inclusion of concentrate in ruminant rations reduces the ratio of structural carbohydrates to starch. This improves diet digestibility and helps to reduce the amount of feed required per unit of animal product [41]. Some authors [42,43,44] have postulated that including concentrate in ruminant diets effectively reduces methane emissions. However, in general, including concentrate in ruminant diets at levels below 60% of the total diet would have no significant impact on methane emissions while only modestly affecting ruminal processes (Hegarty, personal communication). Nevertheless, Hristov et al. [45] reported that the use of concentrate as dietary intervention has a low (≤10%) to medium (10–30%) long-term ability to mitigate enteric methane emissions in ruminants when included at levels beyond 50%.
In the current study, our results are consistent with a grazing trial conducted by Jiao et al. [46] where feeding concentrate as a supplement feed sustained methane emission parameters while improving the dry matter intake and milk yield in Jersey dairy cows. Our analysis revealed that the supply of concentrate improved dry matter intake (kg/day) and milk yield (g/kg DMI) by 23.41% and 19%, respectively, in ruminant livestock. The increase in milk yield (+19%) observed in response to concentrate feeding in ruminants in the current study was also evident in the study of Schilde et al. [47], where dairy cows fed a high (58.2%) versus low (46.7%) concentrate diet had a higher (+14.57%) milk yield. Similarly, Angeles-Hernandez et al. [8] reported an increased milk yield in sheep fed a high concentrate diet compared to a high forage-based diet. In contrast to the findings of Schilde et al. [47], the current study did not show an increase in milk protein content. In fact, the milk protein yield decreased by 16%. Concentrates are typically more energy-dense than forages, providing a greater amount of metabolisable energy per unit of feed. This additional energy can be utilised by dairy ruminants for milk synthesis, resulting in an increased milk yield. Concentrates typically have higher protein levels and are more palatable than forages, resulting in improved dry matter intake and milk production as seen in the current study.
However, it is important to note that the reduction in milk protein content by −16% needs to be considered. The decrease in milk protein concentration can be attributed to the dilution effect caused by the increased milk yield. As milk production increases, the total amount of milk protein synthesized by the animal may still be higher despite the reduced concentration. Therefore, although the milk protein content decreases, the overall milk protein yield might not be significantly affected. Supporting this, Huhtanen and Hetta [48] reported a positive relationship between the level of concentrate intake and milk yield in a meta-analysis study. These results suggest that including concentrate in ruminant diets is an effective strategy to improve dry matter intake and milk yield. While there are no reported studies on the deleterious environmental impacts of including concentrate in ruminant diets, inclusion levels of concentrate beyond a certain threshold are associated with acidosis [49]. Moreover, an increase in dry matter intake is implicated in increasing the passage rate and reducing ruminal degradability [50], which could potentially increase the fermentable organic matter content in faecal excreta and subsequently greenhouse emissions [51]. However, the agronomic traits of the basal diet available, the amount, and the chemical composition of concentrate diets consumed by ruminants should be considered as underlying factors affecting the degree of degradability, passage rate, and ultimately the content of faecal excreta which are not reported in the current analysis.
4.6. Seaweed
Extensive research has been conducted both in the laboratory [52,53,54] and on animals such as sheep [55], beef cattle [56], and dairy cattle [57] to investigate the efficacy of seaweed (i.e., Asparagopsis) in reducing enteric CH4. The outcomes have shown a variation in CH4 reduction, likely due to a variation in diet quality (e.g., fibre content) as reported by Lean et al. [58] and the level of bromoform, a compound derived from seaweed responsible for the reduction in CH4 emissions. In alignment with the outcomes of our research, which exhibited a 35.3% decline in CH4 yield (g/kg DMI) in ruminant livestock, a meta-analysis conducted by Machado et al. [52] examining the efficacy of mainly Asparagopsis algae-based diets in beef cattle also indicated a 37% decrease in CH4 yield. Nevertheless, trials conducted on dairy cows demonstrated inconsistent effectiveness of Asparagopsis. According to Roque et al. [54] and Stefenoni et al. [57], the reported efficacy in CH4 yield was 43% and 80%, respectively, when Asparagopsis was included at levels of 1.84% DM and 0.5% DM, correspondingly. Also, the sole study of Li et al. [59] conducted in sheep reported a range of 15% to 81% CH4 yield mitigating potential when the inclusion level of Asparagopsis varied from 1.0% to 5.7% on DM basis. The meta-analysis study by Almeida et al. [18] reported a comparative higher mean reduction of 49% in CH4 yield across ruminant species subjected to variable feeding levels and diet quality. Such results could be attributed to factors such as diet quality and the inclusion level of seaweed, to name but a few. Nevertheless, certain varieties of seaweeds possess the ability to produce and enclose halogenated methane analogues within specialized gland cells. Among these seaweeds, Asparagopsis has been identified as having bromoform as the primary compound that suppresses methane emissions. Bromoform (CHBr3) and other halogenated methane analogues, such as bromochloromethane (BCM), inhibit the process of methanogenesis by binding to and isolating the prosthetic group necessary for methyl coenzyme M reductase (MCR), which is responsible for the final step in methanogenesis. Bromoform is considered toxic, and exposure to high levels of bromoform can, as a residue in milk or meat products, cause adverse health effects [53]. These effects can include central nervous system depression, liver and kidney damage, respiratory effects, and gastrointestinal disturbances [53].
4.7. Nitroxypropanol
Our study findings align with previous research conducted by Melgar et al. [60] which reported a reduction of 27% in CH4 yield with the use of 3-nitrooxypropanol (3-NOP). Similarly, our study observed a slightly higher reduction of 27.4% in CH4 yield from ruminants upon incorporating 3-NOP. This reduction in CH4 yield was accompanied by an increase in propionate (+13.1%) and butyrate (+17.3%) production, as well as a decrease in the acetate-to-propionate ratio (−20.3%) within rumen fermentation parameters. A decrease in CH4 yield achieved using 3-NOP did not negatively impact dry matter intake or milk yield. Long-term investigation conducted by Melgar et al. [60] consistently demonstrated that incorporating 3-NOP into dairy cattle diets did not result in any adverse effects on DMI or lactation performance. Similarly, Kim et al. [61] specifically focused on beef cattle and found no significant effect on DMI when 3-NOP was included. Our results revealed a 30.5% reduction in CH4 intensity (g/kg milk). Similarly, Melgar et al. [60] reported a 27.4% decrease in CH4 intensity. Melgar et al. [60] reported an increase of 4.83% in milk fat yield when 3-NOP was included in the rations of lactating dairy cows. Equally, the inclusion of 3-NOP was found to increase milk fat concentration in a meta-analysis study by Jayanegara et al. [62]. Consistent with this finding, we observed a noteworthy 15% increase in milk fat yield in response to 3-NOP. To understand the mode of action behind the reduction of CH4 emissions in response to 3-NOP, it is important to consider the normal process of archaeal-associated methane formation in the rumen. This process involves the docking of methyl-coenzyme M reductase with methyl-coenzyme M, resulting in the production of enteric CH4 [63]. In comparison to methyl-coenzyme M, 3-NOP exhibits a strong affinity for methyl-coenzyme M reductase and 3-NOP possesses a similar chemical structure to methyl-coenzyme M [38]. When administered to ruminants, 3-NOP replaces methyl-coenzyme M and prevents the formation of CH4 [38]. This occurs through the oxidation of the nickel in methyl-coenzyme M reductase by 3-NOP, preventing its binding to methyl-coenzyme M [64]. During this reaction, 3-NOP undergoes reduction in intermediate steps, including the formation of nitrate, nitrite, ultimately resulting in the production of 1,3-propanediol as 3-NOP is degraded in the rumen [64]. Our study findings, along with those of Melgar et al. [60] and Jayanegara et al. [6], highlight the significant reduction in CH4 intensity achieved through the utilization of 3-NOP in ruminant diets. This reduction is accompanied by favourable changes in rumen fermentation parameters, including increased propionate and butyrate production, and modified VFA composition. The redirection of energy towards milk fat synthesis contributes to the notable increase in milk fat yield. The consistent findings across these studies emphasize the potential of 3-NOP as a valuable tool in mitigating methane emissions from ruminants while maintaining or even enhancing animal productivity. The consistent administration of 3-NOP through its incorporation into total mixed diets fed to ruminants in confinement maximizes efficacy, while its use in grazing conditions is not yet effective. Furthermore, the mode of action of 3-NOP, involving its affinity for methyl-coenzyme M reductase and the subsequent prevention of CH4 formation, provides mechanistic insights into its effectiveness.
4.8. Biochar
Biochar is a charcoal-like product of the anaerobic combustion of vegetation under an extreme heat (pyrolysis) of between 350 °C to 1100 °C [65]. The porous nature and large Brunauer–Emmett–Teller (BET; 2–40 m2/g) surface area of biochar provides a site for ruminal microbiota allied with methanogenic–methanotrophic interaction, likely mediating CH4 oxidation [66]. The BET is the measure of the ability of a material to absorb gases [67]. The ability of biochar to trap gases in soils [68] could be inferred to the trapping of CH4 in the rumen as made evident in a study by Leng et al. [66], who reported a 22% reduction of CH4 production in “Yellow” cattle fed 0.6% of dietary biochar on a dry matter basis. Our findings demonstrate a 5.5% reduction in CH4 production (g/day) in ruminants which is in agreement with the findings of Leng et al. [66] who observed a similar reduction pattern but lower in magnitude. Conversely, Conlin [68] reported no change in CH4 production and production performance parameters of grazing beef cattle in response to an addition of 1–3% DM of biochar in their diet. Similarly, Winders et al. reported no effects of biochar on CH4 production and CH4 yield in beef steers. Likewise, supplementation of biochar in the diets of lambs had no effect on ADG or gain to feed ratio. Thus, due to the sporadic methane mitigation effect of biochars in the rumen, it is difficult to ascribe the specific mode of action of biochar associated with enteric CH4 mitigation in ruminants [69]. Regardless of this, it is postulated that if CH4 mitigation occurs in the rumen due to the inclusion of biochar in ruminant diets, the reduction could be attributed to the reduction in the rumen methanogenesis-associated protozoa population and/or role of biochar acting as an electron (e−) shuttle responsible for moving e− between ruminal microbes or chemical acceptors and ruminal microbes [63]. In this study, the exact mechanism associated with the observed reduction in CH4 production due to biochar might likely be ascribed to one of the following two notions postulated by Leng et al. [66]. Firstly, it could be stated that biofilm associated with the surface area provided by the inert biochar may have supported the population density of anaerobic methanotrophs sufficiently for CH4 oxidation. Secondly, it is possible that biochar provided a site for the improved efficiency of microbial cell production through the closer association of microbial communities, thereby improving adenosine triphosphate (ATP) production and utilization [66]. However, our findings do not corroborate the second notion suggested by Leng et al. [66] as the efficiency of ATP utilization was not reflected in the improved production performance parameters (i.e., ADG and milk production) of ruminant livestock. Therefore, in this study, it is inferred that the observed reduction in CH4 production due to the use of biochar can be partly explained by the first mode of action described by Leng et al. [66].
5. Conclusions
This study revealed that the various dietary rumen manipulating strategies found to be effective in reducing enteric methane emission differed significantly in terms of the magnitude of emission reduction, mode of emission reduction and subsequent effect on production parameters. Among the identified effective CH4 mitigating strategies that do not adversely affect the production performance of ruminants, saponins have a high effect (>30%) on the reduction of CH4 production, while oil inclusion revealed a high effect in reducing the intensity of methane emission in dairy cows (lower CH4milk); nitrate and 3-NOP showed a medium effect (10–30%) in reducing the intensity of methane emission in dairy ruminants (CH4milk). Similarly, nitrate and 3-NOP has a medium effect in reducing methane yield while oil has a low effect (<10%) in reducing methane yield (CH4yield). On the other hand, concentrate feeding has revealed a beneficial effect by improving the production performance (DMI, MY and MPp) of ruminants without any significant effect on CH4 metrics in ruminant livestock. The inclusion of oil and 3-NOP has provided a co-benefit by improving MFy and MPy in dairy ruminants. Similarly, inclusion of 3-NOP provided co-benefit by improving MFp in dairy ruminants. The observed variation in the magnitude of CH4 reduction and mode of reduction call for further research on the combined use of two or more of the effective CH4 mitigating strategies in conjunction with evaluating their impact at farm-scale levels and assessing the sustainability of their long term use by quantifying net greenhouse gas emissions, through modelling or life cycle analysis, to mediate the adoption rate. Hence, it is advisable to adopt and upscale these strategies with caution, depending on the specific goal to be achieved in ruminant livestock systems.
Author Contributions
Conceptualisation, B.N.P. and A.H.; methodology, B.N.P.; software, validation, formal analysis and investigation, B.N.P.; resources, A.H.; data curation, B.N.P.; writing original draft, B.N.P.; writing—review and editing, B.N.P., A.H. and E.H.T.; visualisation, B.N.P., A.H. and E.H.T.; supervision, A.H. and E.H.T.; project administration, A.H.; funding acquisition, A.H. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partly funded by the Regional Universities Forum for Capacity Building in Agriculture (RUFORUM) and the Global Research Alliance on Agricultural Greenhouse Gases (GRA); RUFORUM-GRA grant provided to the University of Pretoria under Grant number-RU/GRA-GRG/2023/07. Additional funding was provided by the National Research Foundation (NRF) of South Africa under grant No: 139170 and SRUG2204254606 to provide bursary for the first author and cover the article processing fee, respectively.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is unavailable due to privacy.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A
Table A1.
Studies included in the meta-analysis.
References
- Gerber, P.; Opio, C. Food and Agriculture Organization of the United Nations. Animal Production and Health Division. In Greenhouse Gas Emmission from Ruminant Supply Chains: A Global Life Cycle Assessment; FAO: Rome, Italy, 2013; ISBN 9789251079454. [Google Scholar]
- MacLeod, M.J.; Vellinga, T.; Opio, C.; Falcucci, A.; Tempio, G.; Henderson, B.; Makkar, H.; Mottet, A.; Robinson, T.; Steinfeld, H.; et al. Invited Review: A Position on the Global Livestock Environmental Assessment Model (GLEAM). Animal 2018, 12, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Kebreab, E.; Hristov, A.N.; Oh, J.; Arndt, C.; Bannink, A.; Bayat, A.R.; Brito, A.F.; Boland, T.; Casper, D.; et al. Prediction of Enteric Methane Production, Yield, and Intensity in Dairy Cattle Using an Intercontinental Database. Glob. Chang. Biol. 2018, 24, 3368–3389. [Google Scholar] [CrossRef] [PubMed]
- Tseten, T.; Sanjorjo, R.A.; Kwon, M.; Kim, S.W. Strategies to Mitigate Enteric Methane Emissions from Ruminant Animals. J. Microbiol. Biotechnol. 2022, 32, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Hristov, A.N.; Melgar, A.; Wasson, D.; Arndt, C. Symposium Review: Effective Nutritional Strategies to Mitigate Enteric Methane in Dairy Cattle. J. Dairy Sci. 2022, 105, 8543–8557. [Google Scholar] [CrossRef]
- Jayanegara, A.; Sarwono, K.A.; Kondo, M.; Matsui, H.; Ridla, M.; Laconi, E.B.; Nahrowi. Use of 3-Nitrooxypropanol as Feed Additive for Mitigating Enteric Methane Emissions from Ruminants: A Meta-Analysis. Ital. J. Anim. Sci. 2018, 17, 650–656. [Google Scholar] [CrossRef]
- Belanche, A.; Newbold, C.J.; Morgavi, D.P.; Bach, A.; Zweifel, B.; Yáñez-Ruiz, D.R. A Meta-Analysis Describing the Effects of the Essential Oils Blend Agolin Ruminant on Performance, Rumen Fermentation and Methane Emissions in Dairy Cows. Animals 2020, 10, 620. [Google Scholar] [CrossRef]
- Angeles-Hernandez, J.C.; Vieyra Alberto, R.; Kebreab, E.; Appuhamy, J.A.D.R.N.; Dougherty, H.C.; Castelan-Ortega, O.; Gonzalez-Ronquillo, M. Effect of Forage to Concentrate Ratio and Fat Supplementation on Milk Composition in Dairy Sheep: A Meta-Analysis. Livest. Sci. 2020, 238, 104069. [Google Scholar] [CrossRef]
- van Gastelen, S.; Dijkstra, J.; Bannink, A. Are Dietary Strategies to Mitigate Enteric Methane Emission Equally Effective across Dairy Cattle, Beef Cattle, and Sheep? J. Dairy Sci. 2019, 102, 6109–6130. [Google Scholar] [CrossRef]
- de Souza Congio, G.F.; Bannink, A.; Mayorga Mogollón, O.L.; Jaurena, G.; Gonda, H.; Gere, J.I.; Cerón-Cucchi, M.E.; Ortiz-Chura, A.; Tieri, M.P.; Hernández, O.; et al. Enteric Methane Mitigation Strategies for Ruminant Livestock Systems in the Latin America and Caribbean Region: A Meta-Analysis. J. Clean. Prod. 2021, 312, 127693. [Google Scholar] [CrossRef]
- Arndt, C.; Hristov, A.N.; Price, W.J.; McClelland, S.C.; Pelaez, A.M.; Cueva, S.F.; Oh, J.; Dijkstra, J.; Bannink, A.; Bayat, A.R.; et al. Full Adoption of the Most Effective Strategies to Mitigate Methane Emissions by Ruminants Can Help Meet the 1.5 °C Target by 2030 but Not 2050. Proc. Natl. Acad. Sci. USA 2023, 119, e2111294119. [Google Scholar] [CrossRef]
- Benaouda, M.; Martin, C.; Li, X.; Kebreab, E.; Hristov, A.N.; Yu, Z.; Yáñez-Ruiz, D.R.; Reynolds, C.K.; Crompton, L.A.; Dijkstra, J.; et al. Evaluation of the Performance of Existing Mathematical Models Predicting Enteric Methane Emissions from Ruminants: Animal Categories and Dietary Mitigation Strategies. Anim. Feed Sci. Technol. 2019, 255, 114207. [Google Scholar] [CrossRef]
- Congio, G.F.S.; Bannink, A.; Mayorga, O.L.; Rodrigues, J.P.P.; Bougouin, A.; Kebreab, E.; Silva, R.R.; Maurício, R.M.; da Silva, S.C.; Oliveira, P.P.A.; et al. Prediction of Enteric Methane Production and Yield in Dairy Cattle Using a Latin America and Caribbean Database. Sci. Total Environ. 2022, 825, 153982. [Google Scholar] [CrossRef]
- St-Pierre, N.R. Invited Review. Integrating Quantitative Findings from Multiple Studies Using Mixed Model Methodology. J. Dairy Sci. 2001, 84, 741–755. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Pandey, C.M.; Singh, U.; Gupta, A.; Sahu, C.; Keshri, A. Descriptive Statistics and Normality Tests for Statistical Data. Ann. Card. Anaesth. 2019, 22, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K. A Meta-Analysis of the Effect of Dietary Fat on Enteric Methane Production, Digestibility and Rumen Fermentation in Sheep, and a Comparison of These Responses between Cattle and Sheep. Livest. Sci. 2014, 162, 97–103. [Google Scholar] [CrossRef]
- Gurevitch, J.; Hedges, L.V. Statistical Issues in Ecological Meta-Analyses. Ecology 1999, 80, 1142–1149. [Google Scholar] [CrossRef]
- Almeida, A.K.; Hegarty, R.S.; Cowie, A. Meta-Analysis Quantifying the Potential of Dietary Additives and Rumen Modifiers for Methane Mitigation in Ruminant Production Systems. Anim. Nutr. 2021, 7, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Latham, E.A.; Anderson, R.C.; Pinchak, W.E.; Nisbet, D.J. Insights on Alterations to the Rumen Ecosystem by Nitrate and Nitrocompounds. Front. Microbiol. 2016, 7, 228. [Google Scholar] [CrossRef] [PubMed]
- Villar, L.; Hegarty, R.; Van Tol, M.; Godwin, I.; Nolan, J. Dietary Nitrate Metabolism and Enteric Methane Mitigation in Sheep Consuming a Protein-Deficient Diet. Anim. Prod. Sci. 2019, 60, 232–241. [Google Scholar] [CrossRef]
- Goel, G.; Makkar, H.P.S. Methane Mitigation from Ruminants Using Tannins and Saponins. Trop. Anim. Health Prod. 2012, 44, 729–739. [Google Scholar] [CrossRef]
- Abarghuei, M.J.; Rouzbehan, Y.; Salem, A.Z.M.; Zamiri, M.J. Nutrient Digestion, Ruminal Fermentation and Performance of Dairy Cows Fed Pomegranate Peel Extract. Livest. Sci. 2013, 157, 452–461. [Google Scholar] [CrossRef]
- Anantasook, N.; Wanapat, M.; Cherdthong, A. Manipulation of Ruminal Fermentation and Methane Production by Supplementation of Rain Tree Pod Meal Containing Tannins and Saponins in Growing Dairy Steers. J. Anim. Physiol. Anim. Nutr. 2014, 98, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Ma, M.P.; Diao, Q.Y.; Tu, Y. Saponin-Induced Shifts in the Rumen Microbiome and Metabolome of Young Cattle. Front. Microbiol. 2019, 10, 356. [Google Scholar] [CrossRef] [PubMed]
- Lima, P.R.; Apdini, T.; Freire, A.S.; Santana, A.S.; Moura, L.M.L.; Nascimento, J.C.S.; Rodrigues, R.T.S.; Dijkstra, J.; Garcez Neto, A.F.; Queiroz, M.A.Á.; et al. Dietary Supplementation with Tannin and Soybean Oil on Intake, Digestibility, Feeding Behavior, Ruminal Protozoa and Methane Emission in Sheep. Anim. Feed. Sci. Technol. 2019, 249, 10–17. [Google Scholar] [CrossRef]
- Silanikove, N.; Perevolotsky, A.; Provenza, F.D. Use of Tannin-Binding Chemicals to Assay for Tannins and Their Negative Postingestive Effects in Ruminants. Anim. Feed. Sci. Technol. 2001, 91, 69–81. [Google Scholar] [CrossRef]
- Woodward, S.; Waghorn, G.; Laboyrie, P. Condensed Tannins in Birdsfoot Trefoil (Lotus Corniculatus) Reduce Methane Emissions from Dairy Cows. In Proceedings of the New Zealand Society of Animal Production 64th Conference, Hamilton, New Zealand, 28 June–1 July 2004. [Google Scholar]
- Stewart, E.K.; Beauchemin, K.A.; Dai, X.; MacAdam, J.W.; Christensen, R.G.; Villalba, J.J. Effect of Tannin-Containing Hays on Enteric Methane Emissions and Nitrogen Partitioning in Beef Cattle. J. Anim. Sci. 2019, 97, 3286–3299. [Google Scholar] [CrossRef]
- Patra, A.K.; Saxena, J. Exploitation of Dietary Tannins to Improve Rumen Metabolism and Ruminant Nutrition. J. Sci. Food Agric. 2011, 91, 24–37. [Google Scholar] [CrossRef]
- Pepeta, B.N.; Moyo, M.; Adejoro, F.A.; Hassen, A.; Nsahlai, I.V. Techniques Used to Determine Botanical Composition, Intake, and Digestibility of Forages by Ruminants. Agronomy 2022, 12, 2456. [Google Scholar] [CrossRef]
- Mahdavi, A.; Mahdavi, A.; Darabighane, B.; Mead, A.; Lee, M.R.F. Effects of Soybean Oil Supplement to Diets of Lactating Dairy Cows, on Productive Performance, and Milk Fat Acids Profile: A Meta-Analysis. Ital. J. Anim. Sci. 2019, 18, 809–819. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Cantalapiedra-Hijar, G.; Eugène, M.; Martin, C.; Noziere, P.; Popova, M.; Ortigues-Marty, I.; Muñoz-Tamayo, R.; Ungerfeld, E.M. Review: Reducing Enteric Methane Emissions Improves Energy Metabolism in Livestock: Is the Tenet Right? Animal 2023, 17, 100830. [Google Scholar] [CrossRef]
- Bes, A.; Nozière, P.; Renand, G.; Rochette, Y.; Guarnido-Lopez, P.; Cantalapiedra-Hijar, G.; Martin, C. Individual Methane Emissions (and Other Gas Flows) Are Repeatable and Their Relationships with Feed Efficiency Are Similar across Two Contrasting Diets in Growing Bulls. Animal 2022, 16, 100583. [Google Scholar] [CrossRef]
- Guarnido-Lopez, P.; Ortigues-Marty, I.; Salis, L.; Chantelauze, C.; Bes, A.; Nozière, P.; Cantalapiedra-Hijar, G. Protein Metabolism, Body Composition and Oxygen Consumption in Young Bulls Divergent in Residual Feed Intake Offered Two Contrasting Forage-Based Diets. Animal 2022, 16, 100558. [Google Scholar] [CrossRef]
- Villar, M.L.; Hegarty, R.S.; Nolan, J.V.; Godwin, I.R.; McPhee, M. The Effect of Dietary Nitrate and Canola Oil Alone or in Combination on Fermentation, Digesta Kinetics and Methane Emissions from Cattle. Anim. Feed Sci. Technol. 2020, 259, 114294. [Google Scholar] [CrossRef]
- Benchaar, C.; Hassanat, F.; Petit, H.V. Dose-Response to Eugenol Supplementation to Dairy Cow Diets: Methane Production, N Excretion, Ruminal Fermentation, Nutrient Digestibility, Milk Production, and Milk Fatty Acid Profile. Anim. Feed Sci. Technol. 2015, 209, 51–59. [Google Scholar] [CrossRef]
- Martin, C.; Rouel, J.; Jouany, J.P.; Doreau, M.; Chilliard, Y. Methane Output and Diet Digestibility in Response to Feeding Dairy Cows Crude Linseed, Extruded Linseed, or Linseed Oil. J. Anim. Sci. 2008, 86, 2642–2650. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty Years of Research on Rumen Methanogenesis: Lessons Learned and Future Challenges for Mitigation. Animal 2020, 14, S2–S16. [Google Scholar] [CrossRef] [PubMed]
- Kliem, K.E.; Humphries, D.J.; Kirton, P.; Givens, D.I.; Reynolds, C.K. Differential Effects of Oilseed Supplements on Methane Production and Milk Fatty Acid Concentrations in Dairy Cows. Animal 2019, 13, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Giger-Reverdin, S.; Morand-Fehr, P.; Tran, G. Literature Survey of the Influence of Dietary Fat Composition on Methane Production in Dairy Cattle. Livest. Prod. Sci. 2003, 82, 73–79. [Google Scholar] [CrossRef]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited Review: Enteric Methane in Dairy Cattle Production: Quantifying the Opportunities and Impact of Reducing Emissions. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef]
- Boadi, D.; Benchaar, C.; Chiquette, J.; Massé, D.; Massé, J. Mitigation Strategies to Reduce Enteric Methane Emissions from Dairy Cows: Update Review. J. Microbiol. Biotechnol. 2022, 32, 269–277. [Google Scholar] [CrossRef]
- Lovett, D.; Lovell, S.; Stack, L.; Callan, J.; Finlay, M.; Conolly, J.; O’Mara, F.P. Effect of Forage/Concentrate Ratio and Dietary Coconut Oil Level on Methane Output and Performance of Finishing Beef Heifers. Livest. Prod. Sci. 2003, 84, 135–146. [Google Scholar] [CrossRef]
- van Wyngaard, J.D.V.; Meeske, R.; Erasmus, L.J. Effect of Concentrate feeding on Methane Emissions, Production Performance and Rumen Fermentation of Jersey Cows Grazing Ryegrass Pasture during Spring. Anim. Feed Sci. Technol. 2018, 241, 121–132. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Firkins, J.L.; Dijkstra, J.; Kebreab, E.; Waghorn, G.; Makkar, H.P.S.; Adesogan, A.T.; Yang, W.; Lee, C.; et al. SPECIAL TOPICS-Mitigation of Methane and Nitrous Oxide Emissions from Animal Operations: I. A Review of Enteric Methane Mitigation Options 1. J. Anim. Sci. 2013, 91, 5045–5069. [Google Scholar] [CrossRef]
- Jiao, H.P.; Dale, A.J.; Carson, A.F.; Murray, S.; Gordon, A.W.; Ferris, C.P. Effect of Concentrate Feed Level on Methane Emissions from Grazing Dairy Cows. J. Dairy Sci. 2014, 97, 7043–7053. [Google Scholar] [CrossRef] [PubMed]
- Schilde, M.; von Soosten, D.; Hüther, L.; Meyer, U.; Zeyner, A.; Dänicke, S. Effects of 3-Nitrooxypropanol and Varying Concentrate Feed Proportions in the Ration on Methane Emission, Rumen Fermentation and Performance of Periparturient Dairy Cows. Arch. Anim. Nutr. 2021, 75, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Huhtanen, P.; Hetta, M. Comparison of Feed Intake and Milk Production Responses in Continuous and Change-over Design Dairy Cow Experiments. Livest. Sci. 2012, 143, 184–194. [Google Scholar] [CrossRef]
- Maekawa, M.; Beauchemin, K.A.; Christensen, D.A. Effect of Concentrate Level and Feeding Management on Chewing Activities, Saliva Production, and Ruminal PH of Lactating Dairy Cows. J. Dairy Sci. 2002, 85, 1165–1175. [Google Scholar] [CrossRef]
- Ferraretto, L.F.; Shaver, R.D.; Bertics, S.J. Effect of Dietary Supplementation with Live-Cell Yeast at Two Dosages on Lactation Performance, Ruminal Fermentation, and Total-Tract Nutrient Digestibility in Dairy Cows. J. Dairy Sci. 2012, 95, 4017–4028. [Google Scholar] [CrossRef]
- Hristov, A.N.; Lee, C.; Cassidy, T.; Heyler, K.; Tekippe, J.A.; Varga, G.A.; Corl, B.; Brandt, R.C. Effect of Origanum Vulgare L. Leaves on Rumen Fermentation, Production, and Milk Fatty Acid Composition in Lactating Dairy Cows. J. Dairy Sci. 2013, 96, 1189–1202. [Google Scholar] [CrossRef]
- Machado, L.; Magnusson, M.; Paul, N.A.; Kinley, R.; de Nys, R.; Tomkins, N. Identification of Bioactives from the Red Seaweed Asparagopsis Taxiformis that Promote Antimethanogenic Activity In Vitro. J. Appl. Phycol. 2016, 28, 3117–3126. [Google Scholar] [CrossRef]
- Magnusson, M.; Vucko, M.J.; Neoh, T.L.; de Nys, R. Using Oil Immersion to Deliver a Naturally-Derived, Stable Bromoform Product from the Red Seaweed Asparagopsis Taxiformis. Algal Res. 2020, 51, 102065. [Google Scholar] [CrossRef]
- Roque, B.M.; Brooke, C.G.; Ladau, J.; Polley, T.; Marsh, L.J.; Najafi, N.; Pandey, P.; Singh, L.; Kinley, R.; Salwen, J.K.; et al. Effect of the Macroalgae Asparagopsis Taxiformis on Methane Production and Rumen Microbiome Assemblage. Anim. Microbiome 2019, 1, 3. [Google Scholar] [CrossRef]
- El-Waziry, A.; Al-Haidary, A.; Okab, A.; Samara, E.; Abdoun, K. Effect of Dietary Seaweed (Ulva Lactuca) Supplementation on Growth Performance of Sheep and on In Vitro Gas Production Kinetics. Turk. J. Vet. Anim. Sci. 2015, 39, 81–86. [Google Scholar] [CrossRef]
- Kinley, R.D.; Martinez-Fernandez, G.; Matthews, M.K.; de Nys, R.; Magnusson, M.; Tomkins, N.W. Mitigating the Carbon Footprint and Improving Productivity of Ruminant Livestock Agriculture Using a Red Seaweed. J. Clean. Prod. 2020, 259, 120836. [Google Scholar] [CrossRef]
- Stefenoni, H.A.; Räisänen, S.E.; Cueva, S.F.; Wasson, D.E.; Lage, C.F.A.; Melgar, A.; Fetter, M.E.; Smith, P.; Hennessy, M.; Vecchiarelli, B.; et al. Effects of the Macroalga Asparagopsis Taxiformis and Oregano Leaves on Methane Emission, Rumen Fermentation, and Lactational Performance of Dairy Cows. J. Dairy Sci. 2021, 104, 4157–4173. [Google Scholar] [CrossRef] [PubMed]
- Lean, I.J.; Golder, H.M.; Grant, T.M.D.; Moate, P.J. A Meta-Analysis of Effects of Dietary Seaweed on Beef and Dairy Cattle Performance and Methane Yield. PLoS ONE 2021, 16, e0249053. [Google Scholar] [CrossRef]
- Li, X.; Norman, H.C.; Kinley, R.D.; Laurence, M.; Wilmot, M.; Bender, H.; de Nys, R.; Tomkins, N. Asparagopsis Taxiformis Decreases Enteric Methane Production from Sheep. Anim. Prod. Sci. 2018, 58, 681–688. [Google Scholar] [CrossRef]
- Melgar, A.; Nedelkov, K.; Martins, C.M.M.R.; Welter, K.C.; Chen, X.; Räisänen, S.E.; Harper, M.T.; Oh, J.; Duval, S.; Hristov, A.N. Short Communication: Short-Term Effect of 3-Nitrooxypropanol on Feed Dry Matter Intake in Lactating Dairy Cows. J. Dairy Sci. 2020, 103, 11496–11502. [Google Scholar] [CrossRef]
- Kim, H.; Lee, H.G.; Baek, Y.C.; Lee, S.; Seo, J. The Effects of Dietary Supplementation with 3-Nitrooxypropanol on Enteric Methane Emissions, Rumen Fermentation, and Production Performance in Ruminants: A Meta-Analysis. J. Anim. Sci. Technol. 2020, 62, 31–42. [Google Scholar] [CrossRef]
- Jayanegara, A.; Leiber, F.; Kreuzer, M. Meta-Analysis of the Relationship between Dietary Tannin Level and Methane Formation in Ruminants from In Vivo and In Vitro Experiments. J. Anim. Physiol. Anim. Nutr. 2012, 96, 365–375. [Google Scholar] [CrossRef]
- Sun, X.; Cheng, L.; Jonker, A.; Munidasa, S.; Pacheco, D. A Review: Plant Carbohydrate Types—The Potential Impact on Ruminant Methane Emissions. Front. Vet. Sci. 2022, 9, 880115. [Google Scholar] [CrossRef]
- Duin, E.C.; Wagner, T.; Shima, S.; Prakash, D.; Cronin, B.; Yáñez-Ruiz, D.R.; Duval, S.; Rümbeli, R.; Stemmler, R.T.; Thauer, R.K.; et al. Mode of Action Uncovered for the Specific Reduction of Methane Emissions from Ruminants by the Small Molecule 3-Nitrooxypropanol. Proc. Natl. Acad. Sci. USA 2016, 113, 6172–6177. [Google Scholar] [CrossRef]
- Dadhich, A. Engineered Biochar as Feed Supplement and Other Husbandry Applications. In Engineered Biochar: Fundamentals, Preparation, Characterization and Applications; Ramola, S., Mohan, D., Masek, O., Méndez, A., Tsubota, T., Eds.; Springer Nature Singapore: Singapore, 2022; pp. 319–329. ISBN 978-981-19-2488-0. [Google Scholar]
- Leng, R.A.; Preston, T.R.; Inthapanya, S. Biochar Reduces Enteric Methane and Improves Growth and Feed Conversion in Local “Yellow” Cattle Fed Cassava Root Chips and Fresh Cassava Foliage. Livest. Res. Rural. Dev. 2012, 24, 1–7. [Google Scholar]
- Bartzanas, T.; Calvet, S.; Zhang, G. Technology for Environmentally Friendly Livestock Production. In Technol for Environ Friend Livest Product; Bartzanas, T., Ed.; Springer International Publishing: Cham, Switzerland, 2023; pp. 1–9. ISBN 978-3-031-19730-7. [Google Scholar]
- Conlin, E.E. Performance and Environmental Benefits from Biochar Supplementation in Beef Cattle Grazing Systems. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2021. [Google Scholar]
- Hegarty, R. An Evaluation of Evidence for Efficacy and Applicability of Methane Inhibiting Feed Additives for Livestock; New Zealand Agricultural Greenhouse Gas Research Centre: Palmerston North, New Zealand, 2021. [Google Scholar]
- Terry, S.A.; Ribeiro, G.O.; Gruninger, R.J.; Chaves, A.V.; Beauchemin, K.A.; Okine, E.; McAllister, T.A. A Pine Enhanced Biochar Does Not Decrease Enteric CH4 Emissions, but Alters the Rumen Microbiota. Front. Vet. Sci. 2019, 6, 308. [Google Scholar] [CrossRef]
- Doreau, M.; Arbre, M.; Popova, M.; Rochette, Y.; Martin, C. Linseed plus Nitrate in the Diet for Fattening Bulls: Effects on Methane Emission, Animal Health and Residues in Offal. Animal 2018, 12, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Mirheidari, A.; Torbatinejad, N.M.; Shakeri, P.; Mokhtarpour, A. Effects of Biochar Produced from Different Biomass Sources on Digestibility, Ruminal Fermentation, Microbial Protein Synthesis and Growth Performance of Male Lambs. Small Rumin. Res. 2020, 183, 106042. [Google Scholar] [CrossRef]
- Lunsin, R. Effect of Oil Palm Meal on Nutrient Utilization and Milk Production in Lactating Dairy Cows Fed with Urea-Treated Rice Straw. Agric. Nat. Resour. 2018, 52, 285–289. [Google Scholar] [CrossRef]
- Singla, A.; Singh Hundal, J.; Kumar Patra, A.; Wadhwa, M.; Nagarajappa, V.; Malhotra, P. Effect of Dietary Supplementation of Emblica officinalis Fruit Pomace on Methane Emission, Ruminal Fermentation, Nutrient Utilization, and Milk Production Performance in Buffaloes. Environ. Sci. Pollut. Res. 2021, 28, 18120–18133. [Google Scholar] [CrossRef]
- Marino, R.; Caroprese, M.; Annicchiarico, G.; Ciampi, F.; Ciliberti, M.G.; della Malva, A.; Santillo, A.; Sevi, A.; Albenzio, M. Effect of Diet Supplementation with Quinoa Seed and/or Linseed on Immune Response, Productivity and Meat Quality in Merinos Derived Lambs. Animals 2018, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.K.; Kim, Y.J. Effects of Feeding Charcoal Powder and Vitamin A on Growth Performance, Serum Profile and Carcass Characteristics of Fattening Hanwoo Steers. J. Anim. Sci. Technol. 2005, 47, 233–242. [Google Scholar]
- Ortiz-Chura, A.; Gere, J.; Marcoppido, G.; Depetris, G.; Cravero, S.; Faverín, C.; Pinares-Patiño, C.; Cataldi, A.; Cerón-Cucchi, M.E. Dynamics of the Ruminal Microbial Ecosystem, and Inhibition of Methanogenesis and Propiogenesis in Response to Nitrate Feeding to Holstein Calves. Anim. Nutr. 2021, 7, 1205–1218. [Google Scholar] [CrossRef]
- Al-Kindi, A.; Schlecht, E.; Schiborra, A.; Joergensen, R.G. Effects of Quebracho Tannin Extract (Schinopsis Balansae) and Activated Charcoal on Feed Intake and Digestibility by Goats and Their Faecal Microbial Biomass. Biol. Agric. Hortic. 2016, 32, 159–169. [Google Scholar] [CrossRef]
- Winders, T.M.; Jolly-Breithaupt, M.L.; Wilson, H.C.; MacDonald, J.C.; Erickson, G.E.; Watson, A.K. Evaluation of the Effects of Biochar on Diet Digestibility and Methane Production from Growing and Finishing Steers. Transl. Anim. Sci. 2019, 3, 775–783. [Google Scholar] [CrossRef]
- Valenti, B.; Natalello, A.; Vasta, V.; Campidonico, L.; Roscini, V.; Mattioli, S.; Pauselli, M.; Priolo, A.; Lanza, M.; Luciano, G. Effect of Different Dietary Tannin Extracts on Lamb Growth Performances and Meat Oxidative Stability: Comparison between Mimosa, Chestnut and Tara. Animal 2019, 13, 435–443. [Google Scholar] [CrossRef]
- McAvoy, D.J.; Burritt, B.; Villalba, J.J. Use of Biochar by Sheep: Impacts on Diet Selection, Digestibility, and Performance. J. Anim. Sci. 2020, 98, skaa380. [Google Scholar] [CrossRef]
- Liu, H.; Vaddella, V.; Zhou, D. Effects of Chestnut Tannins and Coconut Oil on Growth Performance, Methane Emission, Ruminal Fermentation, and Microbial Populations in Sheep. J. Dairy Sci. 2011, 94, 6069–6077. [Google Scholar] [CrossRef]
- Van, D.T.T.; Mui, N.T.; Ledin, I. Effect of Method of Processing Foliage of Acacia Mangium and Inclusion of Bamboo Charcoal in the Diet on Performance of Growing Goats. Anim. Feed. Sci. Technol. 2006, 130, 242–256. [Google Scholar] [CrossRef]
- Pen, B.; Takaura, K.; Yamaguchi, S.; Asa, R.; Takahashi, J. Effects of Yucca schidigera and Quillaja saponaria with or without β 1-4 Galacto-Oligosaccharides on Ruminal Fermentation, Methane Production and Nitrogen Utilization in Sheep. Anim. Feed. Sci. Technol. 2007, 138, 75–88. [Google Scholar] [CrossRef]
- Lind, L.; Sizmaz, S.; Weldon, D.; Dragan, D.; Jørgensen, G.M. Enteric Methane Emissions from Sheep Fed Diets Including Biochar. In Proceedings of the 28th General Meeting of the European Grassland Federation, Helsinki, Finland, 19–22 October 2020; p. 306. [Google Scholar]
- Wang, C.J.; Wang, S.P.; Zhou, H. Influences of Flavomycin, Ropadiar, and Saponin on Nutrient Digestibility, Rumen Fermentation, and Methane Emission from Sheep. Anim. Feed. Sci. Technol. 2009, 148, 157–166. [Google Scholar] [CrossRef]
- Animut, G.; Puchala, R.; Goetsch, A.L.; Patra, A.K.; Sahlu, T.; Varel, V.H.; Wells, J. Methane Emission by Goats Consuming Different Sources of Condensed Tannins. Anim. Feed. Sci. Technol. 2008, 144, 228–241. [Google Scholar] [CrossRef]
- Liu, H.; Puchala, R.; Leshure, S.; Gipson, T.A.; Flythe, M.D.; Goetsch, A.L. Effects of Lespedeza Condensed Tannins Alone or with Monensin, Soybean Oil, and Coconut Oil on Feed Intake, Growth, Digestion, Ruminal Methane Emission, and Heat Energy by Yearling Alpine Doelings. J. Anim. Sci. 2019, 97, 885–899. [Google Scholar] [CrossRef]
- Poteko, J.; Schrade, S.; Zeyer, K.; Mohn, J.; Zaehner, M.; Zeitz, J.O.; Kreuzer, M.; Schwarm, A. Methane Emissions and Milk Fatty Acid Profiles in Dairy Cows Fed Linseed, Measured at the Group Level in a Naturally Ventilated Housing and Individually in Respiration Chambers. Animals 2020, 10, 1091. [Google Scholar] [CrossRef]
- Mirzaei-Alamouti, H.; Akbari-Pabandi, K.; Mansouryar, M.; Sirjani, M.A.; Cieslak, A.; Szumacher-Strabel, M.; Patra, A.K.; Vazirigohar, M. Effects of Feeding Frequency and Oil Supplementation on Feeding Behavior, Ruminal Fermentation, Digestibility, Blood Metabolites, and Milk Performance in Late-Lactation Cows Fed a High-Forage Diet. J. Dairy Sci. 2020, 103, 11424–11438. [Google Scholar] [CrossRef] [PubMed]
- El-Essawy, A.M.; Anele, U.Y.; Abdel-Wahed, A.M.; Abdou, A.R.; Khattab, I.M. Effects of Anise, Clove and Thyme Essential Oils Supplementation on Rumen Fermentation, Blood Metabolites, Milk Yield and Milk Composition in Lactating Goats. Anim. Feed. Sci. Technol. 2021, 271, 114760. [Google Scholar] [CrossRef]
- Gerlach, K.; Pries, M.; Südekum, K.H. Effect of Condensed Tannin Supplementation on In Vivo Nutrient Digestibilities and Energy Values of Concentrates in Sheep. Small Rumin. Res. 2018, 161, 57–62. [Google Scholar] [CrossRef]
- Robles Jimenez, L.E.; Ruiz Perez, J.A.; Nicolas, D.L.; Chay Canul, A.J.; Ramirez-Rivera, J.C.; Villegas-Estrada, D.; Vargas-Bello-Pérez, E.; Gonzalez-Ronquillo, M. Productive Behavior in Growing Kid Goats and Methane Production with the Inclusion of Chokecherry Leaf (Prunus Salicifolia). Trop. Anim. Health Prod. 2020, 52, 1257–1267. [Google Scholar] [CrossRef]
- Molina-Botero, I.C.; Arroyave-Jaramillo, J.; Valencia-Salazar, S.; Barahona-Rosales, R.; Aguilar-Pérez, C.F.; Ayala Burgos, A.; Arango, J.; Ku-Vera, J.C. Effects of Tannins and Saponins Contained in Foliage of Gliricidia sepium and Pods of Enterolobium cyclocarpum on Fermentation, Methane Emissions and Rumen Microbial Population in Crossbred Heifers. Anim. Feed. Sci. Technol. 2019, 251, 1–11. [Google Scholar] [CrossRef]
- Adejoro, F.A.; Hassen, A.; Akanmu, A.M.; Morgavi, D.P. Replacing Urea with Nitrate as a Non-Protein Nitrogen Source Increases Lambs’ Growth and Reduces Methane Production, Whereas Acacia Tannin Has No Effect. Anim. Feed. Sci. Technol. 2020, 259, 114360. [Google Scholar] [CrossRef]
- Nasehi, M.; Torbatinejad, N.M.; Rezaie, M.; Ghoorchi, T. Effects of Partial Substitution of Alfalfa Hay with Green Tea Waste on Growth Performance and In Vitro Methane Emission of Fat-Tailed Lambs. Small Rumin. Res. 2018, 168, 52–59. [Google Scholar] [CrossRef]
- Puchala, R.; Leshure, S.; Gipson, T.A.; Tesfai, K.; Flythe, M.D.; Goetsch, A.L. Effects of Different Levels of Lespedeza and Supplementation with Monensin, Coconut Oil, or Soybean Oil on Ruminal Methane Emission by Mature Boer Goat Wethers after Different Lengths of Feeding. J. Appl. Anim. Res. 2018, 46, 1127–1136. [Google Scholar] [CrossRef]
- Zhang, F.; Li, B.; Ban, Z.; Liang, H.; Li, L.; Zhao, W.; Yan, X. Evaluation of Origanum Oil, Hydrolysable Tannins and Tea Saponin in Mitigating Ruminant Methane: In Vitro and In Vivo Methods. J. Anim. Physiol. Anim. Nutr. 2021, 105, 630–638. [Google Scholar] [CrossRef]
- Vázquez-Carrillo, M.F.; Montelongo-Pérez, H.D.; González-Ronquillo, M.; Castillo-Gallegos, E.; Castelán-Ortega, O.A. Effects of Three Herbs on Methane Emissions from Beef Cattle. Animals 2020, 10, 1671. [Google Scholar] [CrossRef]
- Thirumalaisamy, G.; Malik, P.K.; Kolte, A.P.; Trivedi, S.; Dhali, A.; Bhatta, R. Effect of Silkworm (Bombyx mori) Pupae Oil Supplementation on Enteric Methane Emission and Methanogens Diversity in Sheep. Anim. Biotechnol. 2022, 33, 128–140. [Google Scholar] [CrossRef]
- Barbosa, A.L.; Voltolini, T.V.; Menezes, D.R.; de Moraes, S.A.; Nascimento, J.C.S.; de Souza Rodrigues, R.T. Intake, Digestibility, Growth Performance, and Enteric Methane Emission of Brazilian Semiarid Non-Descript Breed Goats Fed Diets with Different Forage to Concentrate Ratios. Trop. Anim. Health Prod. 2018, 50, 283–289. [Google Scholar] [CrossRef]
- Heidarian Miri, V.; Tyagi, A.K.; Ebrahimi, S.H.; Mohini, M. Effect of Cumin (Cuminum cyminum) Seed Extract on Milk Fatty Acid Profile and Methane Emission in Lactating Goat. Small Rumin. Res. 2013, 113, 66–72. [Google Scholar] [CrossRef]
- Hulshof, R.B.A.; Berndt, A.; Gerrits, W.J.J.; Dijkstra, J.; Van Zijderveld, S.M.; Newbold, J.R.; Perdok, H.B. Dietary Nitrate Supplementation Reduces Methane Emission in Beef Cattle Fed Sugarcane-Based Diets 1. J. Anim. Sci 2012, 90, 2317–2323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Medrano, R.F.; Wang, M.; Beauchemin, K.A.; Ma, Z.; Wang, R.; Wen, J.; Bernard, L.A.; Tan, Z. Effects of Urea plus Nitrate Pretreated Rice Straw and Corn Oil Supplementation on Fiber Digestibility, Nitrogen Balance, Rumen Fermentation, Microbiota and Methane Emissions in Goats. J. Anim. Sci. Biotechnol 2019, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Van Zijderveld, S.M.; Gerrits, W.J.J.; Dijkstra, J.; Newbold, J.R.; Hulshof, R.B.A.; Perdok, H.B. Persistency of Methane Mitigation by Dietary Nitrate Supplementation in Dairy Cows. J. Dairy Sci. 2011, 94, 4028–4038. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.J.; Mao, S.Y.; Long, L.M.; Zhu, W.Y. Effect of Disodium Fumarate on Microbial Abundance, Ruminal Fermentation and Methane Emission in Goats under Different Forage: Concentrate Ratios. Animal 2012, 6, 1788–1794. [Google Scholar] [CrossRef] [PubMed]
- Granja-Salcedo, Y.T.; Fernandes, R.M.I.; De Araujo, R.C.; Kishi, L.T.; Berchielli, T.T.; De Resende, F.D.; Berndt, A.; Siqueira, G.R. Long-Term Encapsulated Nitrate Supplementation Modulates Rumen Microbial Diversity and Rumen Fermentation to Reduce Methane Emission in Grazing Steers. Front. Microbiol. 2019, 10, 614. [Google Scholar] [CrossRef]
- Hollmann, M.; Powers, W.J.; Fogiel, A.C.; Liesman, J.S.; Bello, N.M.; Beede, D.K. Enteric Methane Emissions and Lactational Performance of Holstein Cows Fed Different Concentrations of Coconut Oil. J. Dairy Sci. 2012, 95, 2602–2615. [Google Scholar] [CrossRef]
- Hundal, J.S.; Wadhwa, M.; Bakshi, M.P.S.; Chatli, M.K. Effect of Herbal Feed Additive Containing Saponins on the Performance of Goat Kids. Indian J. Anim. Sci. 2020, 90, 229–236. [Google Scholar] [CrossRef]
- Paengkoum, S.; Khotsakdee, J.; Paengkoum, P.; Schonewille, T.; Yuangklang, C. Nitrate Supplementation of Rations Based on Rice Straw but Not Pangola Hay, Improves Growth Performance in Meat Goats. Anim. Biosci. 2021, 34, 1022–1028. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.H.; Barnett, M.C.; Hegarty, R.S. Use of Dietary Nitrate to Increase Productivity and Reduce Methane Production of Defaunated and Faunated Lambs Consuming Protein-Deficient Chaff. Anim. Prod. Sci. 2016, 56, 290–297. [Google Scholar] [CrossRef]
- Na, Y.; Li, D.H.; Lee, S.R. Effects of Dietary Forage-to-Concentrate Ratio on Nutrient Digestibility and Enteric Methane Production in Growing Goats (Capra hircus hircus) and Sika Deer (Cervus nippon hortulorum). Asian-Australas. J. Anim. Sci. 2017, 30, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Pilajun, R.; Wanapat, M. Effect of Coconut Oil and Mangosteen Peel Supplementation on Ruminal Fermentation, Microbial Population, and Microbial Protein Synthesis in Swamp Buffaloes. Livest. Sci. 2011, 141, 148–154. [Google Scholar] [CrossRef]
- Anassori, E.; Dalir-Naghadeh, B.; Pirmohammadi, R.; Taghizadeh, A.; Asri-Rezaei, S.; Maham, M.; Farahmand-Azar, S.; Farhoomand, P. Garlic: A Potential Alternative for Monensin as a Rumen Modifier. Livest. Sci. 2011, 142, 276–287. [Google Scholar] [CrossRef]
- Guyader, J.; Eugène, M.; Doreau, M.; Morgavi, D.P.; Gérard, C.; Loncke, C.; Martin, C. Nitrate but Not Tea Saponin Feed Additives Reduce Methane in Dairy Cows. J. Anim. Sci. 2015, 93, 5367–5377. [Google Scholar] [CrossRef]
- Klevenhusen, F.; Zeitz, J.O.; Duval, S.; Kreuzer, M.; Soliva, C.R. Garlic Oil and Its Principal Component Diallyl Disulfide Fail to Mitigate Methane, but Improve Digestibility in Sheep. Anim. Feed. Sci. Technol. 2011, 166–167, 356–363. [Google Scholar] [CrossRef]
- Malik, P.K.; Kolte, A.P.; Baruah, L.; Saravanan, M.; Bakshi, B.; Bhatta, R. Enteric Methane Mitigation in Sheep through Leaves of Selected Tanniniferous Tropical Tree Species. Livest. Sci. 2017, 200, 29–34. [Google Scholar] [CrossRef]
- Verma, V.; Chaudhary, L.C.; Agarwal, N.; Bhar, R.; Kamra, D.N. Effect of Feeding Mixture of Garlic Bulb and Peppermint Oil on Methane Emission, Rumen Fermentation and Microbial Profile in Buffaloes. Anim. Nutr. Feed. Technol. 2012, 12, 157–164. [Google Scholar]
- Tomkins, N.W.; Denman, S.E.; Pilajun, R.; Wanapat, M.; McSweeney, C.S.; Elliott, R. Manipulating Rumen Fermentation and Methanogenesis Using an Essential Oil and Monensin in Beef Cattle Fed a Tropical Grass Hay. Anim. Feed. Sci. Technol. 2015, 200, 25–34. [Google Scholar] [CrossRef]
- Polyorach, S.; Wanapat, M.; Phesatcha, K.; Kang, S. Effect of Different Levels of Mangosteen Peel Powder Supplement on the Performance of Dairy Cows Fed Concentrate Containing Yeast Fermented Cassava Chip Protein. Trop. Anim. Health Prod. 2015, 47, 1473–1480. [Google Scholar] [CrossRef]
- Castro-Montoya, J.; Peiren, N.; Cone, J.W.; Zweifel, B.; Fievez, V.; De Campeneere, S. In Vivo and In Vitro Effects of a Blend of Essential Oils on Rumen Methane Mitigation. Livest. Sci. 2015, 180, 134–142. [Google Scholar] [CrossRef]
- Hundal, J.S.; Singh, I.; Wadhwa, M.; Singh, C.; Uppa, C.; Kaur, G. Effect of Punica granatum and Tecomella undulata supplementation on nutrient utilization, enteric methane emission and growth performance of Murrah male buffaloes. J. Anim. Feed Sci. 2019, 28, 110–119. [Google Scholar] [CrossRef]
- Inamdar, A.I.; Chaudhary, L.C.; Agarwal, N.; Kamra, D.N. Effect of Madhuca longifolia and Terminalia chebula on Methane Production and Nutrient Utilization in Buffaloes. Anim. Feed. Sci. Technol. 2015, 201, 38–45. [Google Scholar] [CrossRef]
- Yatoo, M.A.; Chaudhary, L.C.; Agarwal, N.; Chaturvedi, V.B.; Kamra, D.N. Effect of Feeding of Blend of Essential Oils on Methane Production, Growth, and Nutrient Utilization in Growing Buffaloes. Asian-Australas. J. Anim. Sci. 2018, 31, 672–676. [Google Scholar] [CrossRef]
- Yang, W.Z.; Benchaar, C.; Ametaj, B.N.; Chaves, A.V.; He, M.L.; McAllister, T.A. Effects of Garlic and Juniper Berry Essential Oils on Ruminal Fermentation and on the Site and Extent of Digestion in Lactating Cows. J. Dairy Sci. 2007, 90, 5671–5681. [Google Scholar] [CrossRef] [PubMed]
- Meale, S.J.; Chaves, A.V.; Mcallister, T.A.; Iwaasa, A.D.; Yang, W.Z.; Benchaar, C. Including Essential Oils in Lactating Dairy Cow Diets: Effects on Methane Emissions1. Anim. Prod. Sci. 2014, 54, 1215–1218. [Google Scholar] [CrossRef]
- Manasri, N.; Wanapat, M.; Navanukraw, C. Improving Rumen Fermentation and Feed Digestibility in Cattle by Mangosteen Peel and Garlic Pellet Supplementation. Livest. Sci. 2012, 148, 291–295. [Google Scholar] [CrossRef]
- ManhN, N.S.; Wanapat, M.; Uriyapongson, S.; Khejornsart, P.; Chanthakhoun, V. Effect of Eucalyptus (Camaldulensis) Leaf Meal Powder on Rumen Fermentation Characteristics in Cattle Fed on Rice Straw. Afr. J. Agric. Res. 2012, 7, 614. [Google Scholar] [CrossRef]
- Chaves, A.V.; Stanford, K.; Dugan, M.E.R.; Gibson, L.L.; McAllister, T.A.; Van Herk, F.; Benchaar, C. Effects of Cinnamaldehyde, Garlic and Juniper Berry Essential Oils on Rumen Fermentation, Blood Metabolites, Growth Performance, and Carcass Characteristics of Growing Lambs. Livest. Sci. 2008, 117, 215–224. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Mcginn, S.M. Methane Emissions from Beef Cattle: Effects of Fumaric Acid, Essential Oil, and Canola Oil 1. J. Anim. Sci. 2006, 84, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Matloup, O.H.; Abd El Tawab, A.M.; Hassan, A.A.; Hadhoud, F.I.; Khattab, M.S.A.; Khalel, M.S.; Sallam, S.M.A.; Kholif, A.E. Performance of Lactating Friesian Cows Fed a Diet Supplemented with Coriander Oil: Feed Intake, Nutrient Digestibility, Ruminal Fermentation, Blood Chemistry, and Milk Production. Anim. Feed. Sci. Technol. 2017, 226, 88–97. [Google Scholar] [CrossRef]
- Wall, E.H.; Doane, P.H.; Donkin, S.S.; Bravo, D. The Effects of Supplementation with a Blend of Cinnamaldehyde and Eugenol on Feed Intake and Milk Production of Dairy Cows. J. Dairy Sci. 2014, 97, 5709–5717. [Google Scholar] [CrossRef] [PubMed]
- Giannenas, I.; Skoufos, J.; Giannakopoulos, C.; Wiemann, M.; Gortzi, O.; Lalas, S.; Kyriazakis, I. Effects of Essential Oils on Milk Production, Milk Composition, and Rumen Microbiota in Chios Dairy Ewes. J. Dairy Sci. 2011, 94, 5569–5577. [Google Scholar] [CrossRef] [PubMed]
- Malik, P.; Singhal, K. Effect of Alfalfa Fodder Supplementation on Enteric Methane Emission Measured by Sulfur Hexafluoride Technique in Murrah Buffaloes. Buffalo Bull. 2016, 35, 125–134. [Google Scholar]
- Chiofalo, V.; Liotta, L.; Fiumanò, R.; Riolo, E.B.; Chiofalo, B. Influence of Dietary Supplementation of Rosmarinus officinalis L. on Performances of Dairy Ewes Organically Managed. Small Rumin. Res. 2012, 104, 122–128. [Google Scholar] [CrossRef]
- Muñoz, C.; Herrera, D.; Hube, S.; Morales, J.; Ungerfeld, E.M. Effects of Dietary Concentrate Supplementation on Enteric Methane Emissions and Performance of Late Lactation Dairy Cows. Chil. J. Agric. Res. 2018, 78, 429–437. [Google Scholar] [CrossRef]
- Olijhoek, D.W.; Løvendahl, P.; Lassen, J.; Hellwing, A.L.F.; Höglund, J.K.; Weisbjerg, M.R.; Noel, S.J.; McLean, F.; Højberg, O.; Lund, P. Methane Production, Rumen Fermentation, and Diet Digestibility of Holstein and Jersey Dairy Cows Being Divergent in Residual Feed Intake and Fed at 2 Forage-to-Concentrate Ratios. J. Dairy Sci. 2018, 101, 9926–9940. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Terranova, M.; Kreuzer, M.; Marquardt, S.; Eggerschwiler, L.; Schwarm, A. Supplementation of Pelleted Hazel (Corylus avellana) Leaves Decreases Methane and Urinary Nitrogen Emissions by Sheep at Unchanged Forage Intake. Sci. Rep. 2018, 8, 5427. [Google Scholar] [CrossRef] [PubMed]
- Alves, T.P.; Dall-Orsoletta, A.C.; Ribeiro-Filho, H.M.N. The Effects of Supplementing Acacia mearnsii Tannin Extract on Dairy Cow Dry Matter Intake, Milk Production, and Methane Emission in a Tropical Pasture. Trop. Anim. Health Prod. 2017, 49, 1663–1668. [Google Scholar] [CrossRef] [PubMed]
- Tekippe, J.A.; Tacoma, R.; Hristov, A.N.; Lee, C.; Oh, J.; Heyler, K.S.; Cassidy, T.W.; Varga, G.A.; Bravo, D. Effect of Essential Oils on Ruminal Fermentation and Lactation Performance of Dairy Cows. J. Dairy Sci. 2013, 96, 7892–7903. [Google Scholar] [CrossRef]
- Sun, Y.K.; Yan, X.G.; Ban, Z.B.; Yang, H.M.; Hegarty, R.S.; Zhao, Y.M. The Effect of Cysteamine Hydrochloride and Nitrate Supplementation on In-Vitro and In-Vivo Methane Production and Productivity of Cattle. Anim. Feed. Sci. Technol. 2017, 232, 49–56. [Google Scholar] [CrossRef]
- Abdalla, A.L.; Louvandini, H.; Sallam, S.M.A.H.; da Bueno, I.C.S.; Tsai, S.M.; de Figueira, A.V.O. In Vitro Evaluation, In Vivo Quantification, and Microbial Diversity Studies of Nutritional Strategies for Reducing Enteric Methane Production. Trop. Anim. Health Prod. 2012, 44, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Chilliard, Y.; Martin, C.; Rouel, J.; Doreau, M. Milk Fatty Acids in Dairy Cows Fed Whole Crude Linseed, Extruded Linseed, or Linseed Oil, and Their Relationship with Methane Output. J. Dairy Sci. 2009, 92, 5199–5211. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Kamra, D.N.; Bhar, R.; Kumar, R.; Agarwal, N. Effect of Terminalia chebula and Allium sativum on In Vivo Methane Emission by Sheep. J. Anim. Physiol. Anim. Nutr. 2011, 95, 187–191. [Google Scholar] [CrossRef]
- Ramírez-Restrepo, C.A.; Tan, C.; O’Neill, C.J.; López-Villalobos, N.; Padmanabha, J.; Wang, J.; McSweeney, C.S. Methane Production, Fermentation Characteristics, and Microbial Profiles in the Rumen of Tropical Cattle Fed Tea Seed Saponin Supplementation. Anim. Feed. Sci. Technol. 2016, 216, 58–67. [Google Scholar] [CrossRef]
- Mao, H.L.; Wang, J.K.; Zhou, Y.Y.; Liu, J.X. Effects of Addition of Tea Saponins and Soybean Oil on Methane Production, Fermentation and Microbial Population in the Rumen of Growing Lambs. Livest. Sci. 2010, 129, 56–62. [Google Scholar] [CrossRef]
- Bayat, A.R.; Tapio, I.; Vilkki, J.; Shingfield, K.J.; Leskinen, H. Plant Oil Supplements Reduce Methane Emissions and Improve Milk Fatty Acid Composition in Dairy Cows Fed Grass Silage-Based Diets without Affecting Milk Yield. J. Dairy Sci. 2018, 101, 1136–1151. [Google Scholar] [CrossRef]
- Aguerre, M.J.; Wattiaux, M.A.; Powell, J.M.; Broderick, G.A.; Arndt, C. Effect of Forage-to-Concentrate Ratio in Dairy Cow Diets on Emission of Methane, Carbon Dioxide, and Ammonia, Lactation Performance, and Manure Excretion. J. Dairy Sci. 2011, 94, 3081–3093. [Google Scholar] [CrossRef]
- Olijhoek, D.W.; Hellwing, A.L.F.; Brask, M.; Weisbjerg, M.R.; Højberg, O.; Larsen, M.K.; Dijkstra, J.; Erlandsen, E.J.; Lund, P. Effect of Dietary Nitrate Level on Enteric Methane Production, Hydrogen Emission, Rumen Fermentation, and Nutrient Digestibility in Dairy Cows. J. Dairy Sci. 2016, 99, 6191–6205. [Google Scholar] [CrossRef] [PubMed]
- Serumula, D.M. In Vitro Fermentation and Growth Performance of Merino Lambs Fed on Umbrella Thorn (Vachellia tortilis) Leaf Meal and Sunflower Oil. Ph.D. Thesis, University of KwaZulu-Natal, Pietermaritzburg, South Africa, 2020. [Google Scholar]
- Haisan, J.; Sun, Y.; Guan, L.; Beauchemin, K.A.; Iwaasa, A.; Duval, S.; Kindermann, M.; Barreda, D.R.; Oba, M. The Effects of Feeding 3-Nitrooxypropanol at Two Doses on Milk Production, Rumen Fermentation, Plasma Metabolites, Nutrient Digestibility, and Methane Emissions in Lactating Holstein Cows. Anim. Prod. Sci. 2017, 57, 282–289. [Google Scholar] [CrossRef]
- Vyas, D.; McGinn, S.M.; Duval, S.M.; Kindermann, M.K.; Beauchemin, K.A. Optimal Dose of 3-Nitrooxypropanol for Decreasing Enteric Methane Emissions from Beef Cattle Fed High-Forage and High-Grain Diets. Anim. Prod. Sci. 2018, 58, 1049–1055. [Google Scholar] [CrossRef]
- Romero-Perez, A.; Okine, E.K.; McGinn, S.M.; Guan, L.L.; Oba, M.; Duval, S.M.; Kindermann, M.; Beauchemin, K.A. The Potential of 3-Nitrooxypropanol to Lower Enteric Methane Emissions from Beef Cattle. J. Anim. Sci. 2014, 92, 4682–4693. [Google Scholar] [CrossRef]
- Alemu, A.W.; Pekrul, L.K.D.; Shreck, A.L.; Booker, C.W.; McGinn, S.M.; Kindermann, M.; Beauchemin, K.A. 3-Nitrooxypropanol Decreased Enteric Methane Production from Growing Beef Cattle in a Commercial Feedlot: Implications for Sustainable Beef Cattle Production. Front. Anim. Sci. 2021, 2, 4. [Google Scholar] [CrossRef]
- Zhang, X.M.; Smith, M.L.; Gruninger, R.J.; Kung, L.; Vyas, D.; Mcginn, S.M.; Kindermann, M.; Wang, M.; Tan, Z.L.; Beauchemin, K.A. Combined Effects of 3-Nitrooxypropanol and Canola Oil Supplementation on Methane Emissions, Rumen Fermentation and Biohydrogenation, and Total Tract Digestibility in Beef Cattle. J. Anim. Sci. 2021, 99, skab081. [Google Scholar] [CrossRef]
- Singh, B.K.; Chopra, R.C.; Rai, S.N.; Verma, M.P.; Mohanta, R.K. Nutritional Evaluation of Seaweed on Nutrient Digestibility, Nitrogen Balance, Milk Production and Composition in Sahiwal Cows. Proc. Natl. Acad. Sci. India Sect. B—Biol. Sci. 2017, 87, 437–443. [Google Scholar] [CrossRef]
- Phongphanith, S.; Preston, T.R. Effect of Rice-Wine Distillers’ Byproduct and Biochar on Growth Performance and Methane Emissions in Local “Yellow” Cattle Fed Ensiled Cassava Root, Urea, Cassava Foliage and Rice Straw. Livest. Res. Rural. Dev. 2016, 28, 178. [Google Scholar]
- Terler, G.; Winter, M.; Mandl, M.; Sweeney, J.; Steinwidder, A. Effect of Biochar or Biochar and Urea Supplementation on Feed Intake, Milk Yield, Feed Conversion and Methane Production of Dairy Cows. Czech J. Anim. Sci. 2023, 68, 245–254. [Google Scholar] [CrossRef]
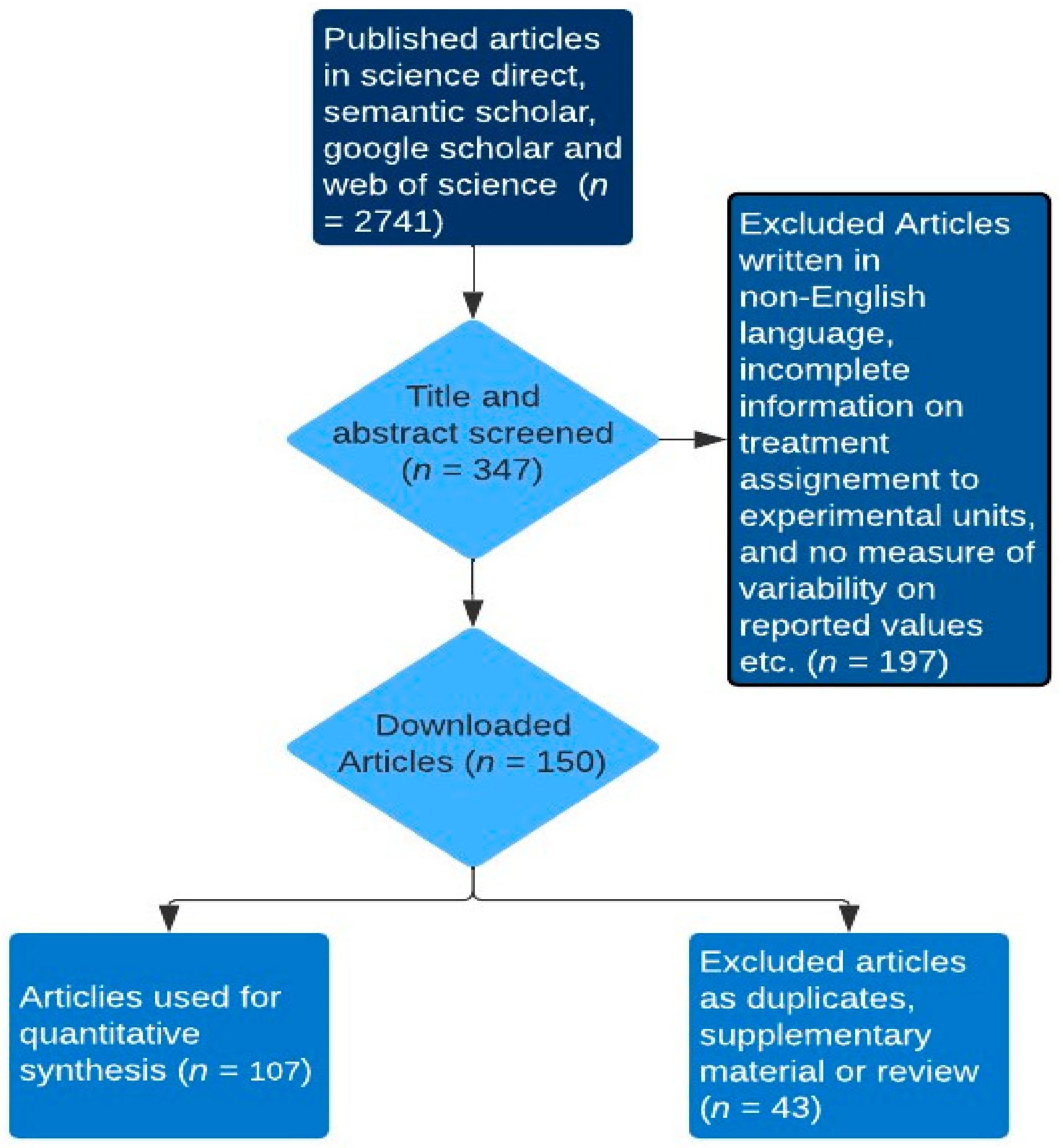
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).