
Effects of Probiotic-Fermented Feed on the Growth Profile, Immune Functions, and Intestinal Microbiota of Bamei Piglets

 ,
,  , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Characteristics of Bacterial Strains
2.2. Fermented Feed Preparation
2.3. Analyses of the Fermented Feeds
2.4. Animal Experimental Design
2.5. Collection of Fecal Samples
2.6. Blood Sample Collection
2.7. 16S rRNA Sequencing
2.8. Statistical Analysis
3. Results
3.1. Fermented Feed Characteristics
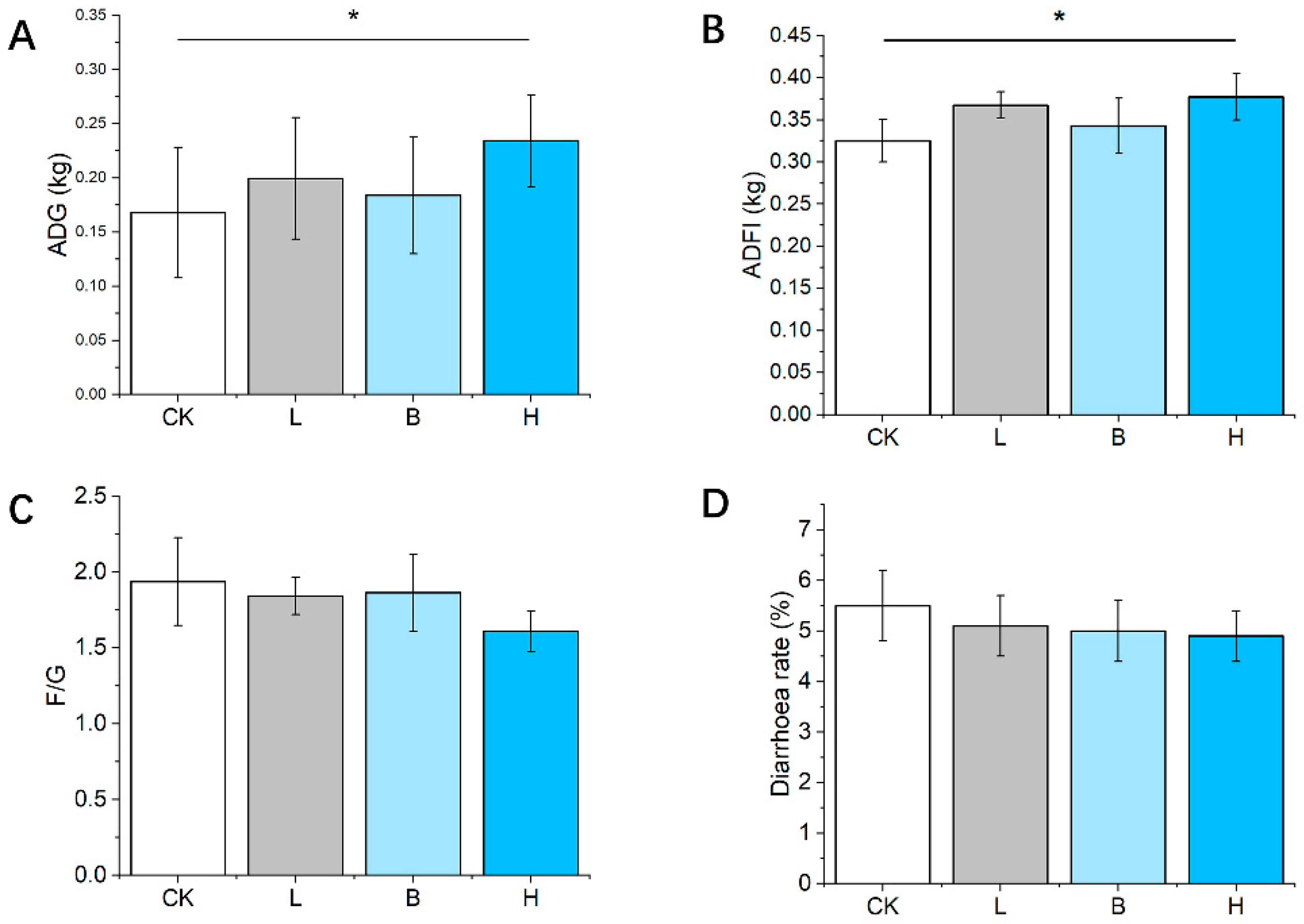
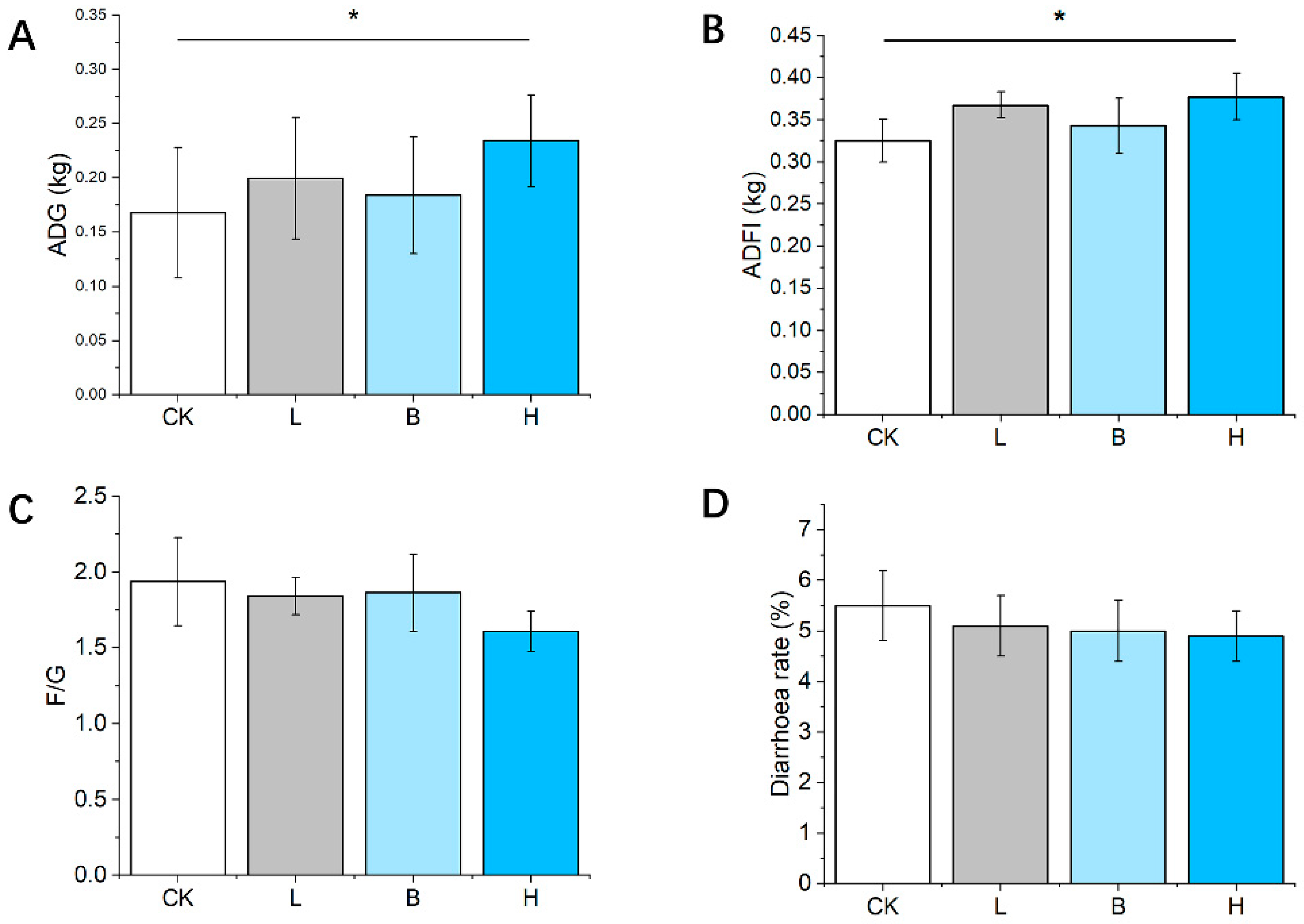
3.2. Growth Performance and Diarrhea Rate of the Piglets
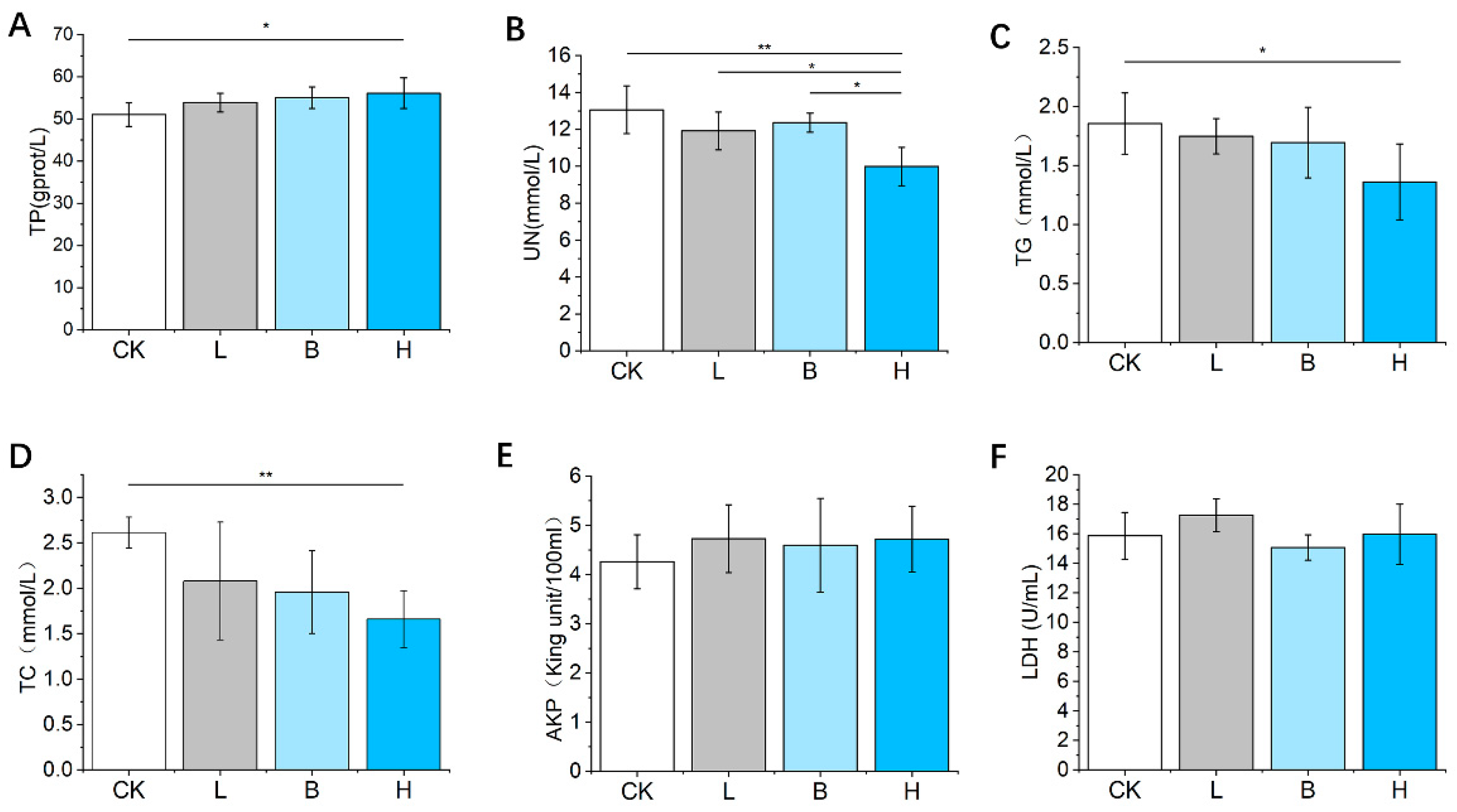
3.3. Changes in Serum Immune Factor Concentrations of the Piglets
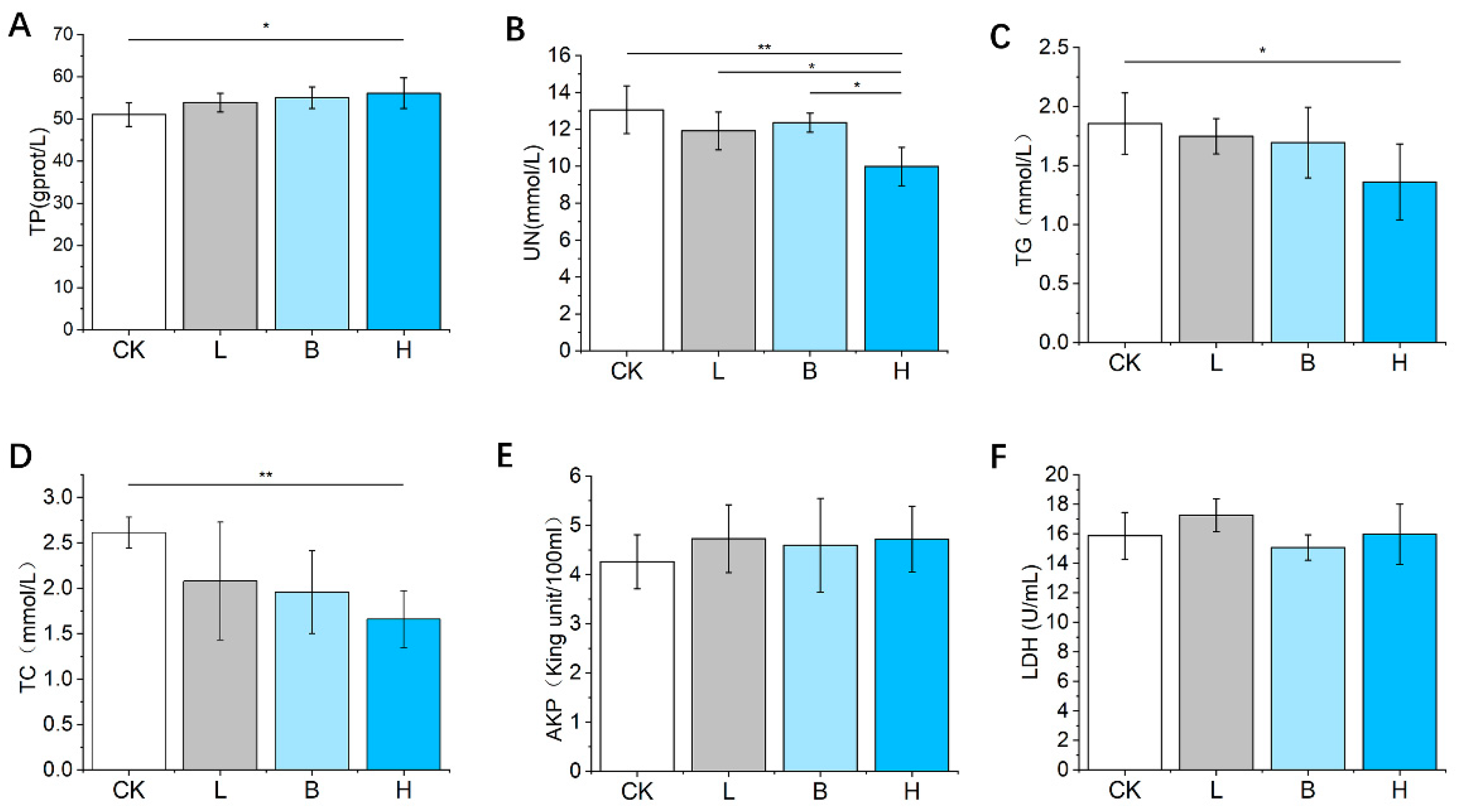
3.4. Changes in Serum Biochemical Factor Concentrations of the Piglets
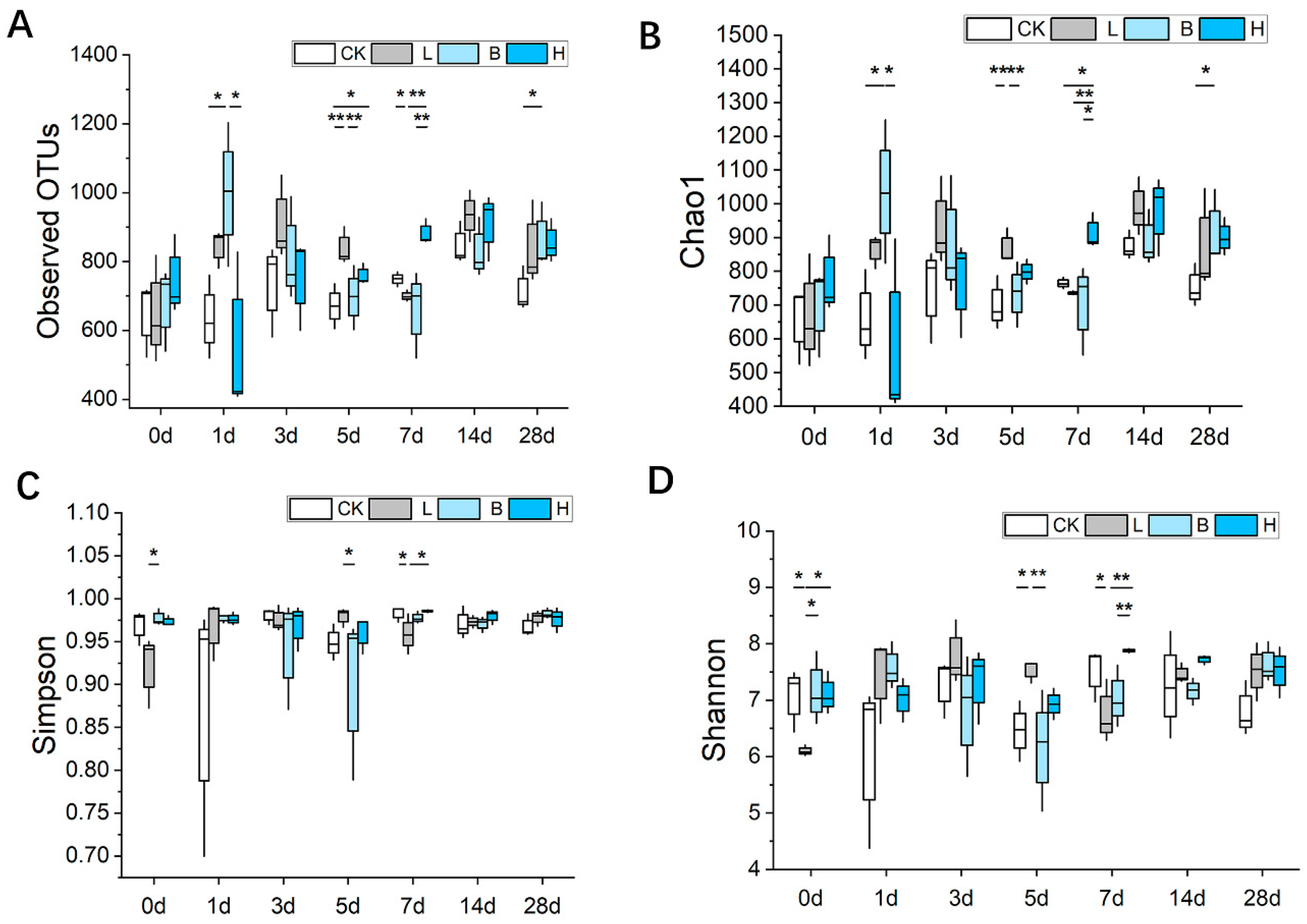
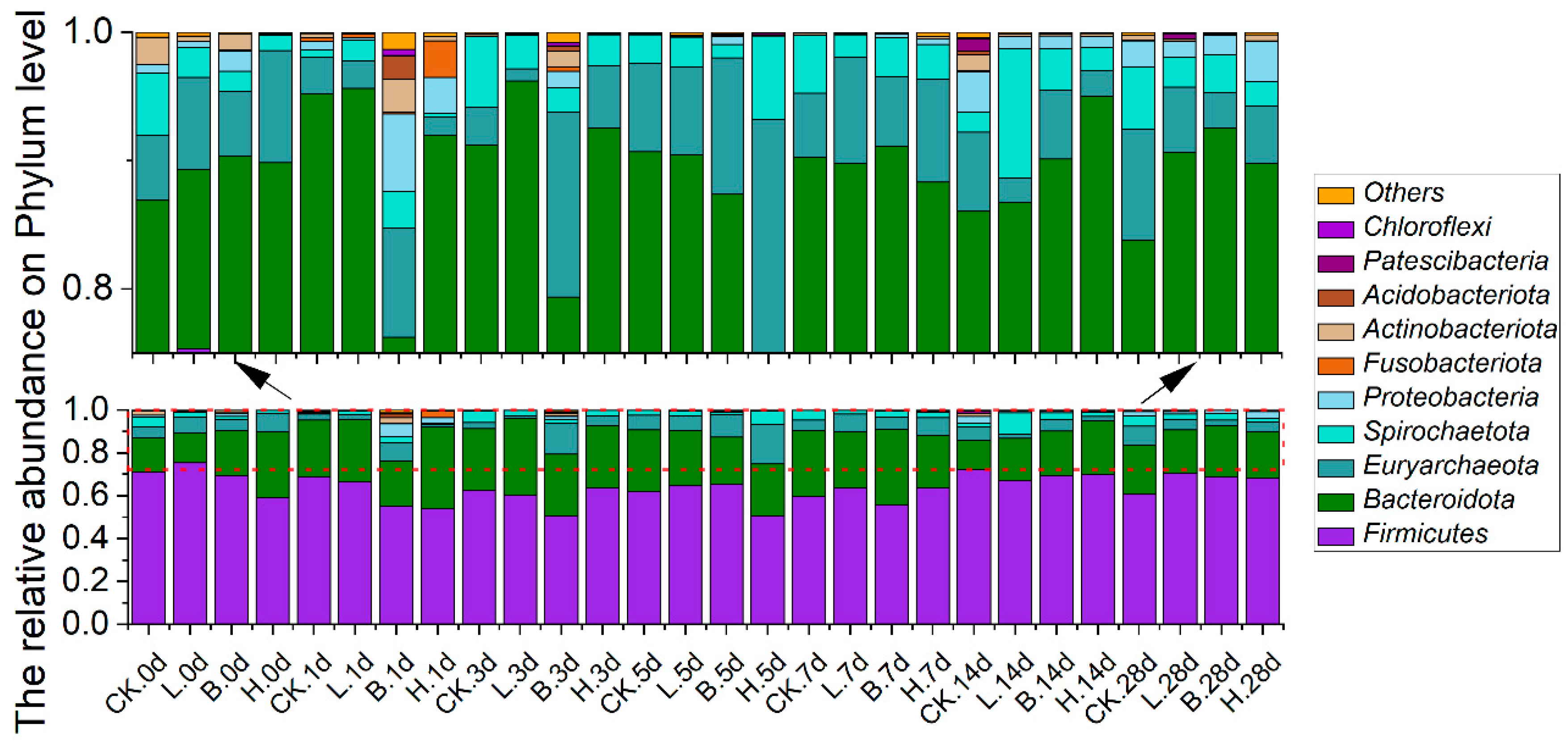
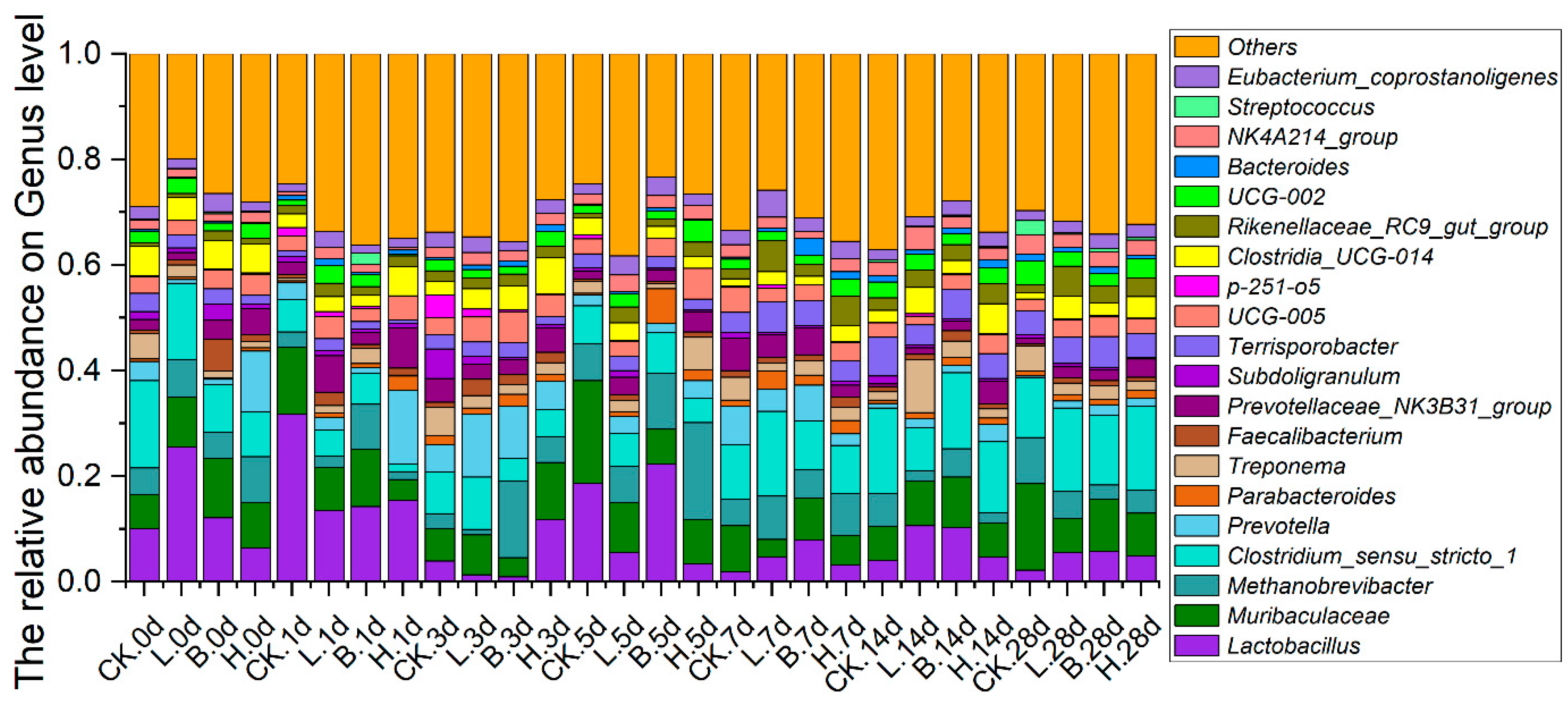
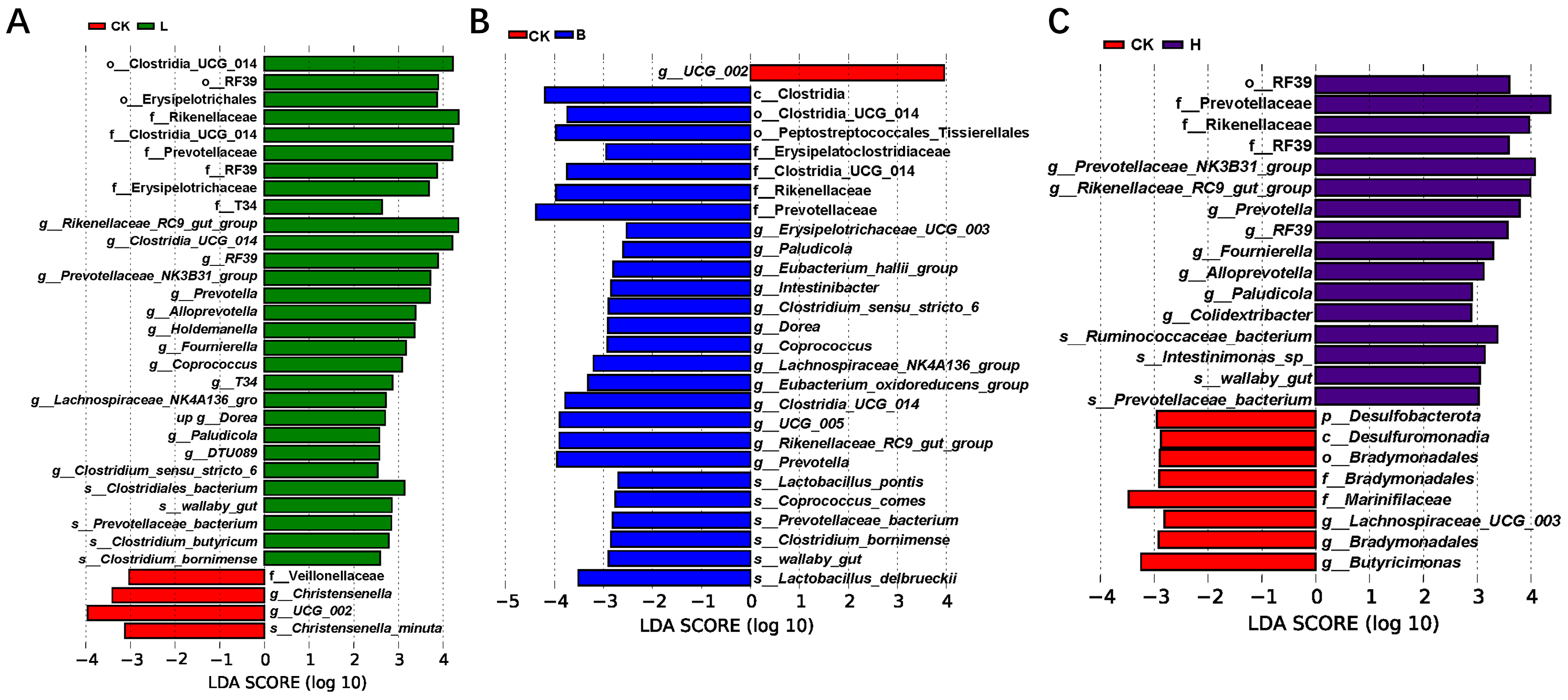
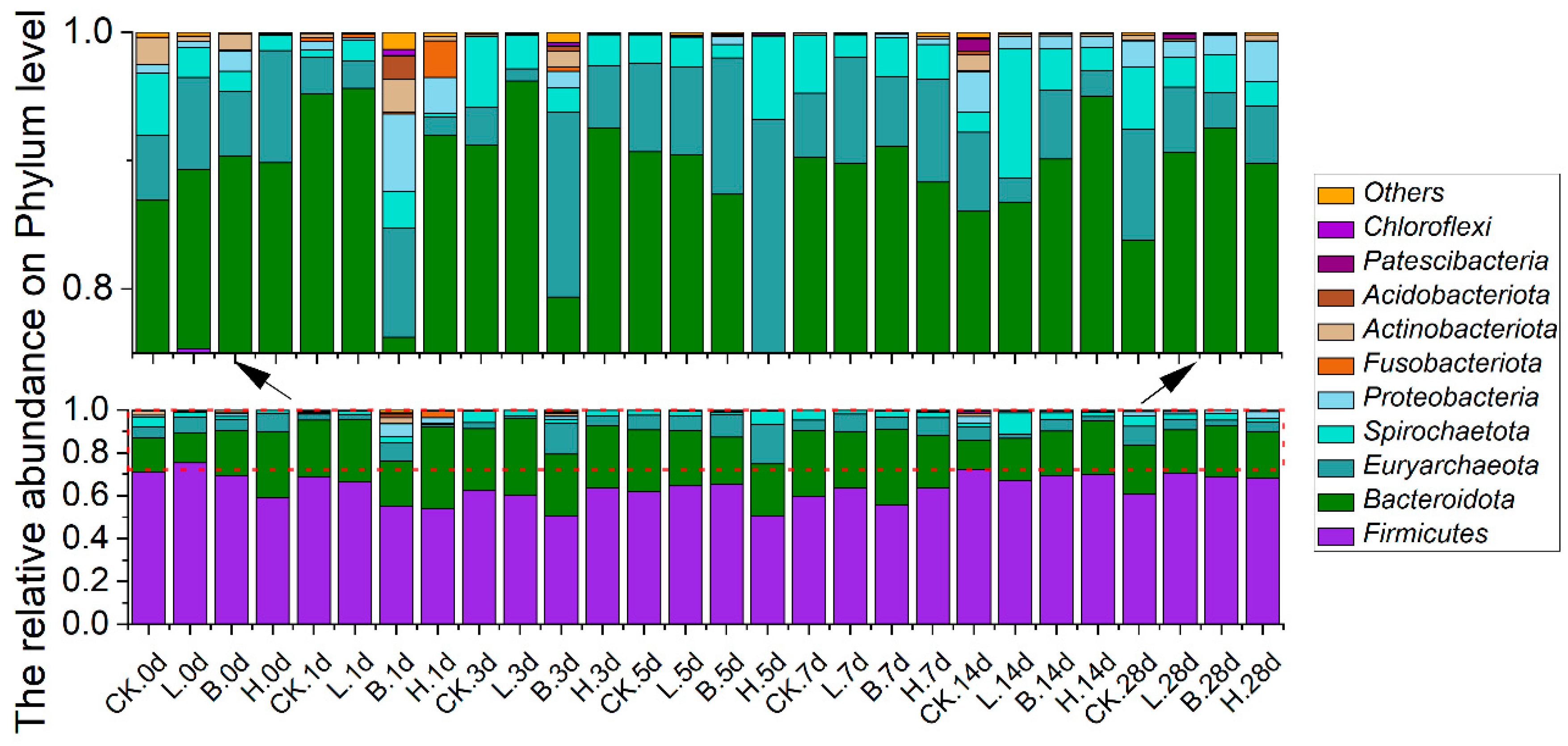
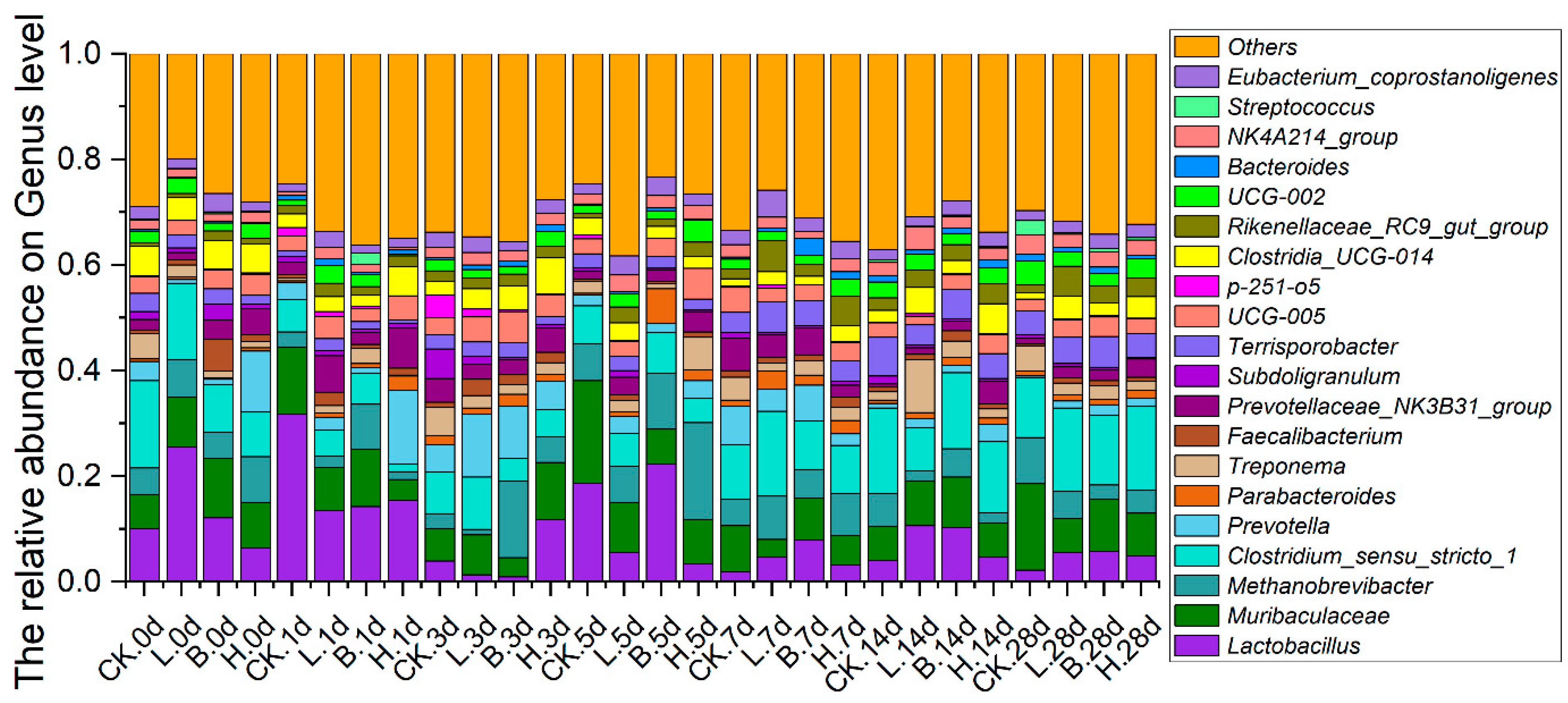
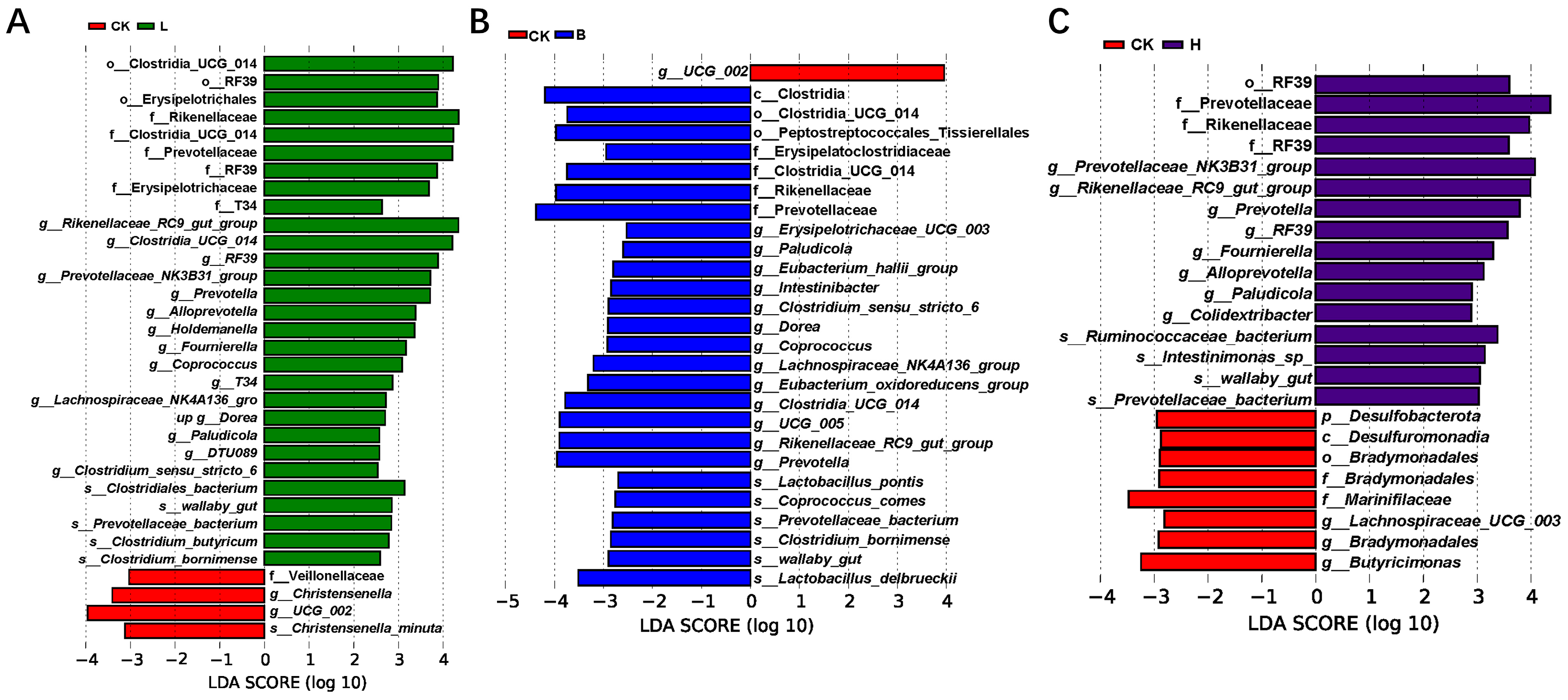
3.5. Changes in the Intestinal Microbiota of the Piglets
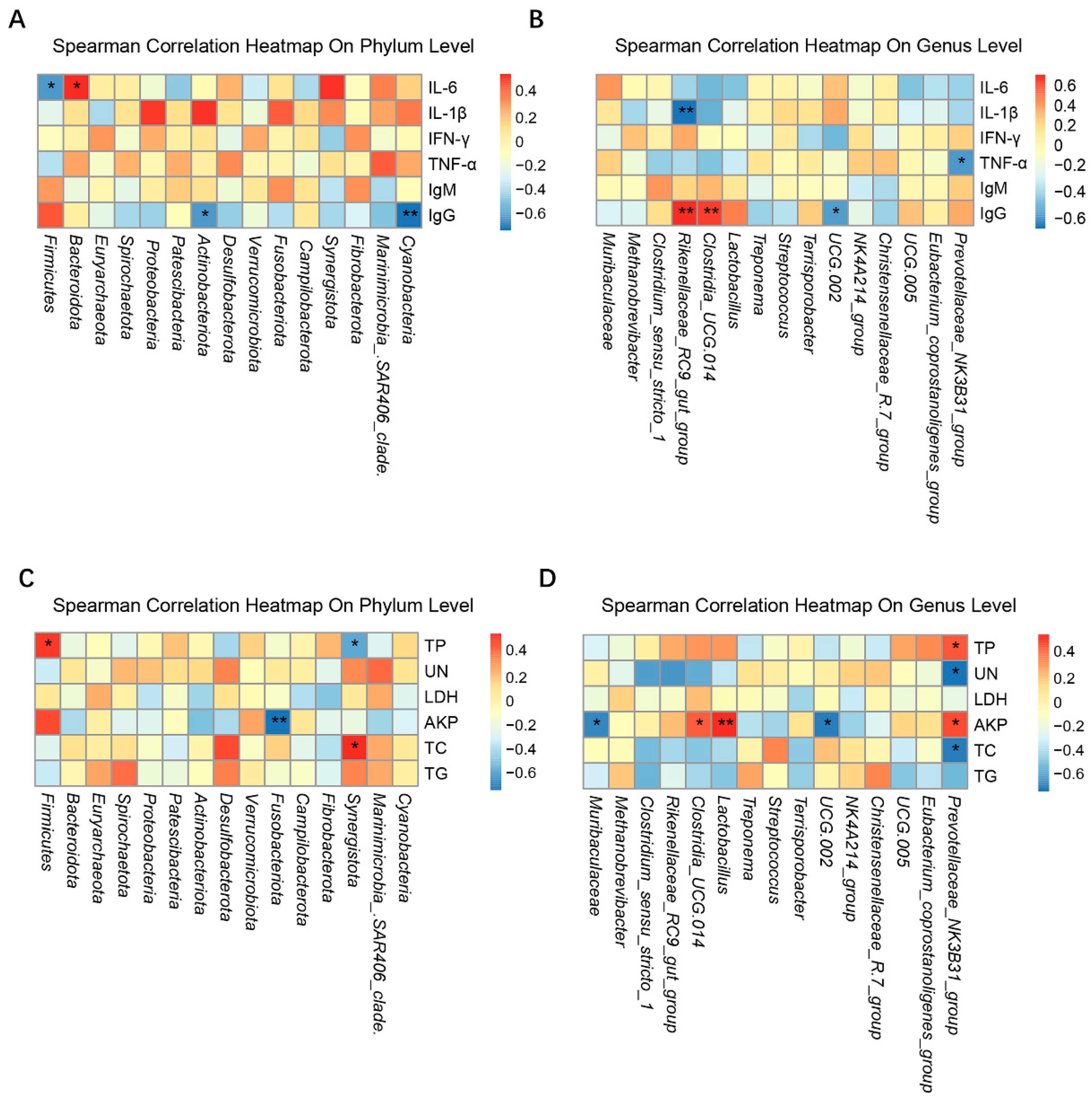
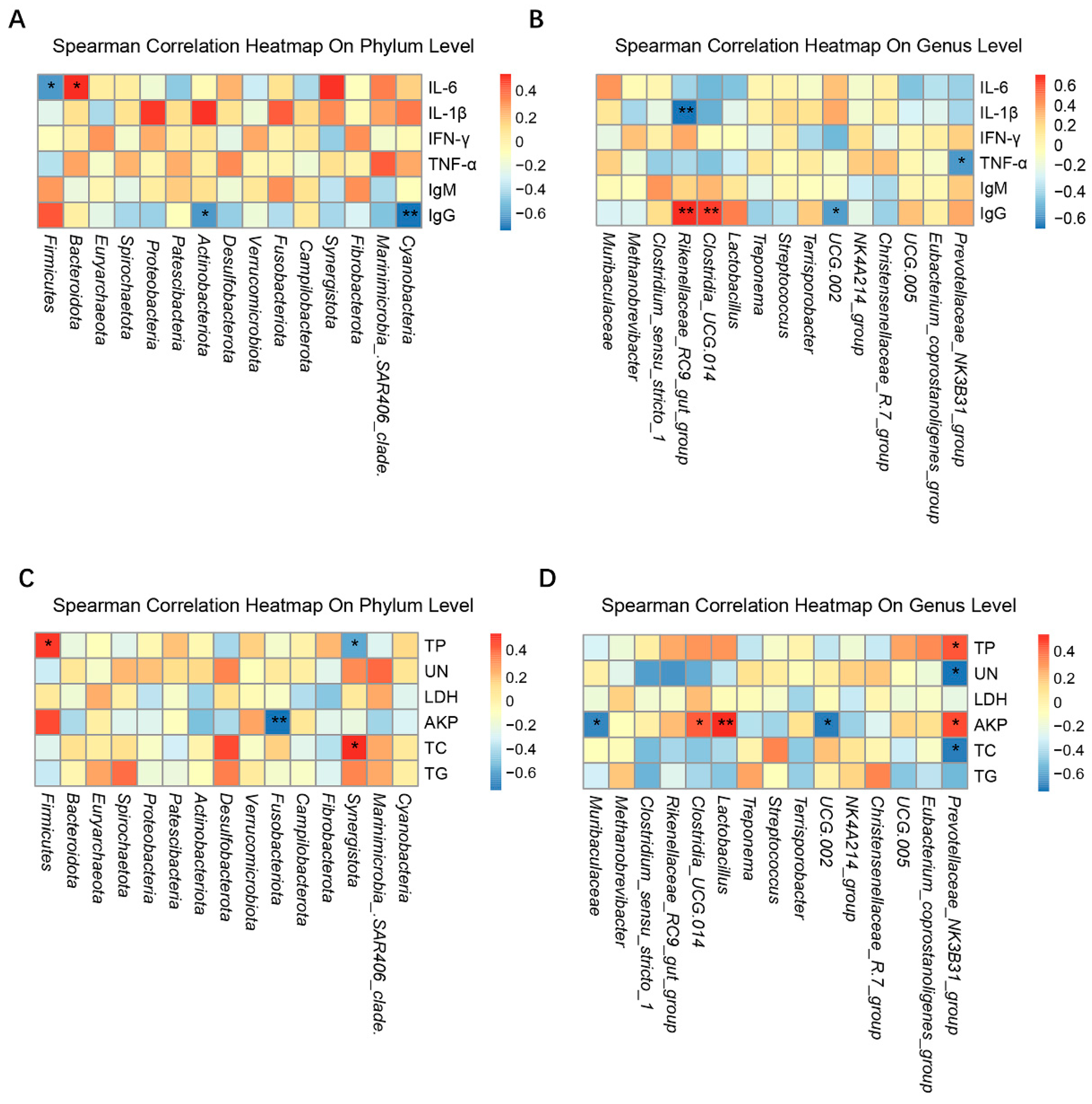
3.6. Correlations between the Serum Immune and Biochemical Factors and the Intestinal Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, B.; Nyachoti, C.M. Husbandry practices and gut health outcomes in weaned piglets: A review. Anim. Nutr. 2017, 3, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, C.; Peng, Y.; Zhang, Y.; Liu, Y.; Liu, Y.; Yin, Y. Research Progress on Anti-Stress Nutrition Strategies in Swine. Anim. Nutr. 2023, 13, 342–360. [Google Scholar] [CrossRef] [PubMed]
- Fouhse, J.M.; Yang, K.; More-Bayona, J.; Gao, Y.; Goruk, S.; Plastow, G.; Field, C.J.; Barreda, D.R.; Willing, B.P. Neonatal Exposure to Amoxicillin Alters Long-Term Immune Response Despite Transient Effects on Gut-Microbiota in Piglets. Front. Immunol. 2019, 10, 2059. [Google Scholar] [CrossRef]
- Shekhar, S.; Brar, N.K.; Petersen, F.C. Suppressive Effect of Therapeutic Antibiotic Regimen on Antipneumococcal Th1/Th17 Responses in Neonatal Mice. Pediatr. Res. 2023, 93, 818–826. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, X.; Zhang, Y.; Zheng, K.; Xiang, Q.; Chen, N.; Zhang, N.; Zhu, J.; He, Q. Antibiotic-Induced Disruption of Gut Microbiota Alters Local Metabolomes and Immune Responses. Front. Cell. Infect. Microbiol. 2019, 9, 99. [Google Scholar] [CrossRef]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23, 795. [Google Scholar] [CrossRef]
- Kumar, S.B.; Arnipalli, S.R.; Ziouzenkova, O. Antibiotics in Food Chain: The Consequences for Antibiotic Resistance. Antibiotics 2020, 9, 688. [Google Scholar] [CrossRef]
- Landers, T.F.; Cohen, B.; Wittum, T.E.; Larson, E.L. A Review of Antibiotic Use in Food Animals: Perspective, Policy, and Potential. Public Health Rep. 2012, 127, 4–22. [Google Scholar] [CrossRef] [PubMed]
- Wan, M.L.Y.; Forsythe, S.J.; El-Nezami, H. Probiotics interaction with foodborne pathogens: A potential alternative to antibiotics and future challenges. Crit. Rev. Food Sci. Nutr. 2019, 59, 3320–3333. [Google Scholar] [CrossRef]
- Tsukahara, T.; Kimura, Y.; Inoue, R.; Iwata, T. Preliminary investigation of the use of dietary supplementation with probiotic Bacillus subtilis strain QST713 shows that it attenuates antimicrobial-induced dysbiosis in weaned piglets. Anim. Sci. J. 2020, 91, e13475. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Zhang, M.; Tu, W.; Zhang, Q.; Jin, M.; Fang, R.; Jiang, S. Bacillus subtilis inhibits intestinal inflammation and oxidative stress by regulating gut flora and related metabolites in laying hens. Animal 2022, 16, 100474. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Qian, Y.; Yu, B.; Zhang, T.; Gao, J.; He, J.; Huang, Z.; Zheng, P.; Mao, X.; Chen, D.; et al. Effects of Bacillus subtilis Dsm32315 Supplementation and Dietary Crude Protein Level on Performance, Gut Barrier Function and Microbiota Profile in Weaned Piglets1. J. Anim. Sci. 2019, 97, 2125–2138. [Google Scholar] [CrossRef]
- Namted, S.; Poungpong, K.; Loongyai, W.; Rakangthong, C.; Bunchasak, C. Dietary autolysed yeast modulates blood profiles, small intestinal morphology and caecal microbiota of weaning pigs. Animal 2022, 16, 100660. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Su, W.; Li, W.; Wen, C.; Du, S.; He, H.; Zhang, Y.; Gong, T.; Wang, X.; Lu, Z.; et al. Bacillus amyloliquefaciens 40 Regulates Piglet Performance, Antioxidant Capacity, Immune Status and Gut Microbiota. Anim. Nutr. 2023, 12, 116–127. [Google Scholar] [CrossRef]
- Wang, K.; Cao, G.; Zhang, H.; Li, Q.; Yang, C. Effects of Clostridium Butyricum and Enterococcus Faecalis on Growth Performance, Immune Function, Intestinal Morphology, Volatile Fatty Acids, and Intestinal Flora in a Piglet Model. Food Funct. 2019, 10, 7844–7854. [Google Scholar] [CrossRef]
- Shi, C.; Zhang, Y.; Lu, Z.; Wang, Y. Solid-state fermentation of corn-soybean meal mixed feed with Bacillus subtilis and Enterococcus faecium for degrading antinutritional factors and enhancing nutritional value. J. Anim. Sci. Biotechnol. 2017, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Zhu, L.; Fu, J.; Li, Z.; Wang, Y.; Jin, M. Overall Assessment of Fermented Feed for Pigs: A Series of Meta-Analyses. J. Anim. Sci. 2019, 97, 4810–4821. [Google Scholar] [CrossRef]
- Zhu, X.; Tao, L.; Liu, H.; Yang, G. Effects of fermented feed on growth performance, immune organ indices, serum biochemical parameters, cecal odorous compound production, and the microbiota community in broilers. Poult. Sci. 2023, 102, 102629. [Google Scholar] [CrossRef]
- Yang, A.; Zuo, Y.; Cheng, Z.; Wu, X.; Li, P. Tong, and H. Chen. Degradation of Major Allergens and Allergenicity Reduction of Soybean Meal through Solid-State Fermentation with Microorganisms. Food Funct. 2018, 9, 1899–1909. [Google Scholar] [CrossRef]
- Huang, J.; Dai, Y.; Zhang, Y.; Liu, G.; Peng, F.; Xie, M.; Xiong, T. Dynamics of Bacterial Community, Metabolites Profile and Physicochemical Characteristics During Solid-State Fermentation of Soybean Meal and Corn Mixed Substrates Inoculated with Bacillus pumilus and Limosilactobacillus fermentum. J. Sci. Food Agric. 2023, 103, 5588–5599. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, B.; Liu, G.; Shi, H.; Wang, J. Effect of Bacillus Subtilis and Lactobacillus plantarum on Solid-State Fermentation of Soybean Meal. J. Sci. Food Agric. 2023, 103, 6070–6079. [Google Scholar] [CrossRef]
- Garrido-Galand, S.; Asensio-Grau, A.; Calvo-Lerma, J.; Heredia, A.; Andrés, A. The potential of fermentation on nutritional and technological improvement of cereal and legume flours: A review. Food Res. Int. 2021, 145, 110398. [Google Scholar] [CrossRef]
- Dai, C.; Hou, Y.; Xu, H.; Huang, L.; Dabbour, M.; Mintah, B.K.; He, R.; Ma, H. Effect of solid-state fermentation by three different Bacillus species on composition and protein structure of soybean meal. J. Sci. Food Agric. 2022, 102, 557–566. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, R.; Lei, H.; Hang, Y.; Xue, H.; Cai, X.; Lu, Y. Supplementation with Fermented Feedstuff Enhances Orexin Expression and Secretion Associated with Increased Feed Intake and Weight Gain in Weaned Pigs. Animals 2022, 12, 1329. [Google Scholar] [CrossRef]
- Liu, S.; Xiao, H.; Xiong, Y.; Chen, J.; Wu, Q.; Wen, X.; Jiang, Z.; Wang, L. Effects of Fermented Feed on the Growth Per-formance, Intestinal Function, and Microbiota of Piglets Weaned at Different Age. Front. Vet. Sci. 2022, 9, 841762. [Google Scholar] [CrossRef]
- Xie, K.; Dai, Y.; Zhang, A.; Yu, B.; Luo, Y.; Li, H.; He, J. Effects of Fermented Soybean Meal on Growth Performance, Meat Quality, and Antioxidant Capacity in Finishing Pigs. J. Funct. Foods 2022, 94, 105128. [Google Scholar] [CrossRef]
- Zhu, J.M.; Gao, R.; Zhang, Z.; Sun, C.; Wang, F.; Yang, T.; Huang Tingting, S.; Qu, S.; Zhao, L.; Li, Y.; et al. Effects of Soybean Meal Fermented by L. plantarum, B. subtilis and S. cerevisieae on Growth, Immune Function and Intestinal Morphology in Weaned Piglets. Microb. Cell Fact. 2017, 16, 191. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhu, M.; Cao, H.; Zhang, X.; Wang, Z.; Zhang, X.; Li, X.; Hu, J.; Yang, G.; Shi, X.E. Impact of Fermented Corn-Soybean Meal on Gene Expression of Immunity in the Blood, Level of Secretory Immunoglobulin a, and Mucosa-Associated Bacterial Community in the Intestine of Grower–Finisher Pigs. Front. Vet. Sci. 2020, 7, 246. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.; Bustamante-García, D.; Kim, J.W.; Nyachoti, C.M. Health-Promoting Effects of Lactobacillus-Fermented Barley in Weaned Pigs Challenged with Escherichia coli K88. Animal 2020, 14, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Farzan, A.; Wang, Q.; Yu, H.; Yin, Y.; Hou, Y.; Friendship, R.; Gong, J. Reduction of Salmonella enterica Serovar Typhimurium Dt104 Infection in Experimentally Challenged Weaned Pigs Fed a Lactobacillus-Fermented Feed. Foodborne Pathog. Dis. 2014, 11, 628–634. [Google Scholar] [CrossRef]
- Wu, G.; Tang, X.; Fan, C.; Wang, L.; Shen, W.; Ren, S.E.; Zhang, L.Z.; Zhang, Y.M. Gastrointestinal Tract and Dietary Fiber Driven Alterations of Gut Microbiota and Metabolites in Durco x Bamei Crossbred Pigs. Front. Nutr. 2022, 8, 806646. [Google Scholar] [CrossRef]
- Chen, J.; Pang, H.; Wang, L.; Ma, C.; Wu, G.; Liu, Y.; Guan, Y.; Zhang, M.; Qin, G.; Tan, Z. Bacteriocin-Producing Lactic Acid Bacteria Strains with Antimicrobial Activity Screened from Bamei Pig Feces. Foods 2022, 11, 709. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Lv, J.; Liu, Y.; Ma, H.; Chen, B.; Hao, K.; Feng, J.; Min, Y. Effects of Different Fermented Feeds on Production Performance, Cecal Microorganisms, and Intestinal Immunity of Laying Hens. Animals 2021, 11, 2799. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Feng, J.; Wang, Y.; Lv, J.; Li, J.; Guo, L.; Min, Y. Fermented Corn–Soybean Meal Mixed Feed Modulates Intestinal Morphology, Barrier Functions and Cecal Microbiota in Laying Hens. Animals 2021, 11, 3059. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Miao, H.; Zhang, C.; Wang, Y.; Liu, S.; Jiao, P.; Li, W.; Li, Y.; Huang, Z. Effect of dietary Bacillus coagulans on the performance and intestinal microbiota of weaned piglets. Animal 2022, 16, 100561. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zeng, X.; Qiao, S. Advances in research on solid-state fermented feed and its utilization: The pioneer of private customization for intestinal microorganisms. Anim. Nutr. 2021, 7, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.H.; Cho, S.J. Improvement of Bioactivity of Soybean Meal by Solid-State Fermentation with Bacillus amyloliquefaciens Versus Lactobacillus spp. And Saccharomyces cerevisiae. LWT-Food Sci. Technol. 2016, 68, 619–625. [Google Scholar] [CrossRef]
- Akhtar, N.; Cai, H.Y.; Kiarie, E.G.; Li, J. A novel Bacillus sp. with rapid growth property and high enzyme activity that allows efficient fermentation of soybean meal for improving digestibility in growing pigs. J. Appl. Microbiol. 2022, 133, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Wang, B.; Zhu, J.; Yang, G.; Zhou, Y.; Pan, Y.; Tao, Y.; Zhong, J. Effects of Lactic Acid Bacteria and Molasses Additives on the Microbial Community and Fermentation Quality of Soybean Silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef]
- Varsha, K.K.; Priya, S.; Devendra, L.; Nampoothiri, K.M. Control of Spoilage Fungi by Protective Lactic Acid Bacteria Displaying Probiotic Properties. Appl. Biochem. Biotechnol. 2014, 172, 3402–3413. [Google Scholar] [CrossRef]
- Xu, B.; Li, Z.; Wang, C.; Fu, J.; Zhang, Y.; Wang, Y.; Lu, Z. Effects of Fermented Feed Supplementation on Pig Growth Performance: A Meta-Analysis. J. Anim. Sci. 2019, 97, 125–126. [Google Scholar] [CrossRef]
- Vadopalas, L.; Ruzauskas, M.; Lele, V.; Starkute, V.; Zavistanaviciute, P.; Zokaityte, E.; Bartkevics, V.; Pugajeva, I.; Reinolds, I.; Badaras, S.; et al. Combination of Antimicrobial Starters for Feed Fermentation: Influence on Piglet Feces Microbiota and Health and Growth Performance, Including Mycotoxin Biotransformation in vivo. Front. Vet. Sci. 2020, 7, 528990. [Google Scholar] [CrossRef]
- Meinlschmidt, P.; Ueberham, J.; Lehmann, J.; Schweiggert-Weisz, U.; Eisner, P. Immunoreactivity, Sensory and Physico-chemical Properties of Fermented Soy Protein Isolate. Food Chem. 2016, 205, 229–238. [Google Scholar] [CrossRef]
- Wang, C.; Lin, C.; Su, W.; Zhang, Y.; Wang, F.; Wang, Y.; Wang, C.; Shi, C.; Lu, Z. Effects of Supplementing Sow Diets with Fermented Corn and Soybean Meal Mixed Feed During Lactation on the Performance of Sows and Progeny. J. Anim. Sci. 2018, 96, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, X.; Hao, Y.; Duan, Z.; Meng, Z.; An, X.; Qi, J. Dietary Fermented Soybean Meal Replacement Alleviates Diarrhea in Weaned Piglets Challenged with Enterotoxigenic Escherichia coli K88 by Modulating Inflammatory Cytokine Levels and Cecal Microbiota Composition. BMC Vet. Res. 2020, 16, 245. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.H.; Yu, Y.H. Evaluation of Bacillus licheniformis-Fermented Feed Additive as an Antibiotic Substitute: Effect on the Growth Performance, Diarrhea Incidence, and Cecal Microbiota in Weaning Piglets. Animals 2020, 10, 1649. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.K.; Sarker, M.; Wan, D. Immunomodulatory Effects of Probiotics on Cytokine Profiles. BioMed Res. Int. 2018, 2018, 8063647. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chu, D.; Kalantar-Zadeh, K.; George, J.; Young, H.A.; Liu, G. Cytokines: From Clinical Significance to Quantification. Adv. Sci. 2021, 8, e2004433. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Hu, H.; Chen, D.; Tang, J.; Yu, B.; Luo, J.; Yu, J.; Mao, X. Dietary Pectic Oligosaccharide Administration Improves Growth Performance and Immunity in Weaned Pigs Infected by Rotavirus. J. Agric. Food Chem. 2017, 65, 2923–2929. [Google Scholar] [CrossRef] [PubMed]
- Mallery, D.L.; McEwan, W.A.; Bidgood, S.R.; Towers, G.J.; Johnson, C.M.; James, L.C. Antibodies Mediate Intracellular Immunity through Tripartite Motif-Containing 21 (Trim21). Proc. Natl. Acad. Sci. USA 2010, 107, 19985–19990. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, X.; Liu, Y.; Cao, H.; Han, Q.; Xie, B.; Fan, L.; Li, X.; Hu, J.; Yang, G.; et al. Effect of Fermented Corn-Soybean Meal on Serum Immunity, the Expression of Genes Related to Gut Immunity, Gut Microbiota, and Bacterial Metabolites in Grower-Finisher Pigs. Front. Microbiol. 2019, 10, 2620. [Google Scholar] [CrossRef]
- Mizumachi, K.; Aoki, R.; Ohmori, H.; Saeki, M.; Kawashima, T. Effect of fermented liquid diet prepared with Lactobacillus plantarum LQ80 on the immune response in weaning pigs. Animal 2009, 3, 670–676. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Li-Wen, S.; Horng, Y.B.; Yu-Hsiang, Y. Effects of Soybean Meal Fermented by Lactobacillus Species and Clostridium Butyricum on Growth Performance, Diarrhea Incidence, and Fecal Bacteria in Weaning Piglets. Ann. Anim. Sci. 2019, 19, 1051–1062. [Google Scholar] [CrossRef]
- Wang, S.; Yao, B.; Gao, H.; Zang, J.; Tao, S.; Zhang, S.; Huang, S.; He, B.; Wang, J. Combined Supplementation of Lactobacillus fermentum and Pediococcus acidilactici Promoted Growth Performance, Alleviated Inflammation, and Modulated Intestinal Microbiota in Weaned Pigs. BMC Vet. Res. 2019, 15, 239. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Wang, Y.; Li, Z.; Qamar, H.; Mehmood, K.; Zhang, L.; Liu, J.; Zhang, H.; Li, J. Probiotics isolated from yaks improves the growth performance, antioxidant activity, and cytokines related to immunity and inflammation in mice. Microb. Cell Factories 2019, 18, 112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Liu, F.; Mao, Y.; Zhang, Y.; Zeng, H.; Ren, S.; Guo, L.; Chen, Z.; Hrabchenko, N.; et al. Mechanisms and Applications of Probiotics in Prevention and Treatment of Swine Diseases. Porc. Health Manag. 2023, 9, 5. [Google Scholar] [CrossRef]
- Sun, P.; Wang, J.; Zhang, H. Effects of Bacillus subtilis natto on performance and immune function of preweaning calves. J. Dairy Sci. 2010, 93, 5851–5855. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Xu, L.N.; Guo, X.J.; Wang, W.; Hao, Q.H.; Wang, S.; Zhu, B. The Impacts of Fermented Feed on Laying Performance, Egg Quality, Immune Function, Intestinal Morphology and Microbiota of Laying Hens in the Late Laying Cycle. Animal 2022, 16, 100676. [Google Scholar] [CrossRef] [PubMed]
- Vimalraj, S. Alkaline phosphatase: Structure, expression and its function in bone mineralization. Gene 2020, 754, 144855. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Wang, C.; Ye, J.; Chen, H.; Tao, R. Effects of Dietary Supplementation of Fermented Ginkgo biloba L. Residues on Growth Performance, Nutrient Digestibility, Serum Biochemical Parameters and Immune Function in Weaned Piglets. Anim. Sci. J. 2015, 86, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Qu, T.; Yang, L.; Wang, Y.; Jiang, B.; Shen, M.; Ren, D. Reduction of serum cholesterol and its mechanism by Lactobacillus plantarum H6 screened from local fermented food products. Food Funct. 2020, 11, 1397–1409. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, T.M.; Sun, W.; Bumbie, G.Z.; Dosoky, W.M.; Rao, Z.; Hu, P.; Wu, L.; Tang, Z. Effect of Dietary Supplementation of Bacillus subtilis on Growth Performance, Organ Weight, Digestive Enzyme Activities, and Serum Biochemical Indices in Broiler. Animals 2022, 12, 1558. [Google Scholar] [CrossRef] [PubMed]
- Ashayerizadeh, A.; Dastar, B.; Shargh, M.S.; Mahoonak, A.S.; Zerehdaran, S. Effects of feeding fermented rapeseed meal on growth performance, gastrointestinal microflora population, blood metabolites, meat quality, and lipid metabolism in broiler chickens. Livest. Sci. 2018, 216, 183–190. [Google Scholar] [CrossRef]
- Kim, H.B.; Borewicz, K.; White, B.A.; Singer, R.S.; Sreevatsan, S.; Tu, Z.J.; Isaacson, R.E. Longitudinal investigation of the age-related bacterial diversity in the feces of commercial pigs. Vet. Microbiol. 2011, 153, 124–133. [Google Scholar] [CrossRef]
- Isaacson, R.; Kim, H.B. The intestinal microbiome of the pig. Anim. Health Res. Rev. 2012, 13, 100–109. [Google Scholar] [CrossRef]
- Nowland, T.L.; Plush, K.J.; Barton, M.; Kirkwood, R.N. Development and Function of the Intestinal Microbiome and Potential Implications for Pig Production. Animals 2019, 9, 76. [Google Scholar] [CrossRef]
- Li, L.; Zhao, X. Comparative analyses of fecal microbiota in Tibetan and Chinese Han living at low or high altitude by barcoded 454 pyrosequencing. Sci. Rep. 2015, 5, 14682. [Google Scholar] [CrossRef]
- Zhang, D.; Ji, H.; Liu, H.; Wang, S.; Wang, J.; Wang, Y. Changes in the Diversity and Composition of Gut Mi-crobiota of Weaned Piglets after Oral Administration of Lactobacillus or an Antibiotic. Appl. Microbiol. Biotechnol. 2016, 100, 10081–10093. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Litvak, Y.; Byndloss, M.X.; Tsolis, R.M.; Bäumler, A.J. Dysbiotic Proteobacteria Expansion: A Microbial Signature of Epithelial Dysfunction. Curr. Opin. Microbiol. 2017, 39, 1–6. [Google Scholar] [CrossRef]
- Reuben, R.; Roy, P.; Sarkar, S.; Alam, A.R.U.; Jahid, I. Characterization and evaluation of lactic acid bacteria from indigenous raw milk for potential probiotic properties. J. Dairy Sci. 2020, 103, 1223–1237. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Shi, J.; Zhu, J.; Shao, D.; Huang, Q.; Yang, H.; Jin, M. Capacity of Lactic Acid Bacteria in Immunity Enhancement and Cancer Prevention. Appl. Microbiol. Biotechnol. 2017, 101, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Kuroki, Y.; Oka, K.; Takahashi, M.; Rao, S.; Sukegawa, S.; Fujimura, T. Effects of Dietary Supplementation with Enterococcus faecium and Clostridium butyricum, Either Alone or in Combination, on Growth and Fecal Microbiota Composition of Post-weaning Pigs at a Commercial Farm. Front. Vet. Sci. 2019, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liang, J.; Liu, Y.; Zhou, X.; Peng, C.; Long, C.; Huang, P.; Feng, J.; Zhang, Z. Combined Supplementation with Lactobacillus Sp. And Bifidobacterium thermacidophilum Isolated from Tibetan Pigs Improves Growth Performance, Immunity, and Microbiota Composition in Weaned Piglets. J. Anim. Sci. 2023, 101, skad220. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, M.; Ma, Y.; Yang, Y.; Cheng, Y.; Ma, H.; Ren, D.; Chen, P. Regulation of Viable/Inactivated/Lysed Probiotic Lactobacillus plantarum H6 on Intestinal Microbiota and Metabolites in Hypercholesterolemic Mice. NPJ Sci. Food 2022, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.; Gálvez, E.J.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B.; et al. Sequence and cultivation study of Muribaculaceae reveals novel species, host preference, and functional potential of this yet undescribed family. Microbiome 2019, 7, 28. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Tian, S.; Liu, J.; Huang, S.; Yang, M.; Yang, X.; Xu, R.; Lin, J.; Han, L.; Zhang, D. Combination Therapy with Indigo and Indirubin for Ulcerative Colitis Via Reinforcing Intestinal Barrier Function. Oxid. Med. Cell. Longev. 2023, 2023, 2894695. [Google Scholar] [CrossRef]
- Yang, W.-Y.; Lee, Y.; Lu, H.; Chou, C.-H.; Wang, C. Analysis of gut microbiota and the effect of lauric acid against necrotic enteritis in Clostridium perfringens and Eimeria side-by-side challenge model. PLoS ONE 2019, 14, e0205784. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Wang, H.-G.; Zhang, M.-N.; Zhang, M.-H.; Wang, H.; Yang, X.-Z. Fecal microbiota transplantation ameliorates experimental colitis via gut microbiota and T-cell modulation. World J. Gastroenterol. 2021, 27, 2834–2849. [Google Scholar] [CrossRef] [PubMed]
- Burakova, I.; Smirnova, Y.; Gryaznova, M.; Syromyatnikov, M.; Chizhkov, P.; Popov, E.; Popov, V. The Effect of Short-Term Consumption of Lactic Acid Bacteria on the Gut Microbiota in Obese People. Nutrients 2022, 14, 3384. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, Y.; Liu, J.; Wang, H.; Guo, Y.; Du, M.; Cai, C.; Zhao, Y.; Lu, C.; Guo, X.; et al. Composition of the Fecal Microbiota of Piglets at Various Growth Stages. Front. Vet. Sci. 2021, 8, 661671. [Google Scholar] [CrossRef]
- Holman, D.B.; Gzyl, K.E. A meta-analysis of the bovine gastrointestinal tract microbiota. FEMS Microbiol. Ecol. 2019, 95, fiz072. [Google Scholar] [CrossRef]
- Ramsay, T.G.; Arfken, A.M.; Summers, K.L. Enteroendocrine peptides, growth, and the microbiome during the porcine weaning transition. Anim. Microbiome 2022, 4, 56. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | QP28-1 | QB8 |
|---|---|---|
| Species | Lactobacillus plantarum | Bacillus subtilis |
| Collecting location | Huzhu County, Qinghai, China | |
| Sample sources | Feces of Bamei pig | |
| Shape | rod | rod |
| Gram stain | + | + |
| Growth in NaCl | ||
| 3.5 | ++ | − |
| 6.5 | ++ | ++ |
| Growth performance at Temperature (°C) | ||
| 5 | w | − |
| 10 | + | + |
| 30 | ++ | ++ |
| 45 | + | ++ |
| 50 | w | + |
| Growth performance at pH | ||
| 3 | − | − |
| 4 | + | − |
| 5 | ++ | + |
| 8 | ++ | +++ |
| 9 | ++ | ++ |
| 10 | + | + |
| Antibacterial activity | ||
| Escherichia coli ATCC 30105 | ++ | + |
| Micrococcus luteus ATCC 4698 | +++ | ++ |
| Staphvlococcus aureus ATCC 29213 | ++ | ++ |
| Pseudomonas aeruginosa ATCC 27853 | ++ | − |
| Listeria monocytogenes BAA | ++++ | ++++ |
| Salmonella enterica ATCC 43971 | ++ | ++ |
| Ingredients | Contents (%) | Nutritional Indicators 2 | |
|---|---|---|---|
| Corn | 60.00 | Digestible energy (MJ/kg) | 14.78 |
| Soybean meal | 26.00 | DM (%) | 89.16 |
| Fish meal | 2.00 | CP (%) | 19.45 |
| Wheat bran | 3.00 | Crude fiber (%) | 3.12 |
| Whey powder | 5.00 | Crude fat (%) | 2.15 |
| Premix 1 | 4.00 | Crude ash (%) | 5.18 |
| Total | 100 | ADF (%) | 4.15 |
| NDF (%) | 13.45 | ||
| NaCl (%) | 0.50 | ||
| Calcium (%) | 1.02 | ||
| Total Phosphorus (%) | 0.58 |
| Items | Prior to Fermentation | After Fermentation | SEM | p-Value | |
|---|---|---|---|---|---|
| L. plantarum QP28-1 Fermented Feed | B. subtilis QB-8 Fermented Feed | ||||
| CP (%) | 19.45 B | 22.63 A | 22.80 A | 0.13 | 0.017 |
| Crude fat (%) | 2.15 | 1.86 | 1.81 | 0.05 | 0.055 |
| Crude ash (%) | 5.18 | 4.91 | 4.85 | 0.05 | 0.052 |
| ADF (%) | 4.15 | 4.05 | 4.00 | 0.02 | 0.071 |
| NDF (%) | 13.35 A | 12.17 B | 11.5 B | 0.03 | 0.047 |
| DM (%) | 89.16 | 87.11 | 87.35 | 1.24 | 0.061 |
| pH | 6.24 A | 4.44 B | 4.71 B | 0.15 | 0.009 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Yang, Z.; Wu, G.; Xu, F.; Zhang, J.; Luo, X.; Ma, Y.; Pang, H.; Duan, Y.; Chen, J.; et al. Effects of Probiotic-Fermented Feed on the Growth Profile, Immune Functions, and Intestinal Microbiota of Bamei Piglets. Animals 2024, 14, 647. https://doi.org/10.3390/ani14040647
Zhang M, Yang Z, Wu G, Xu F, Zhang J, Luo X, Ma Y, Pang H, Duan Y, Chen J, et al. Effects of Probiotic-Fermented Feed on the Growth Profile, Immune Functions, and Intestinal Microbiota of Bamei Piglets. Animals. 2024; 14(4):647. https://doi.org/10.3390/ani14040647
Chicago/Turabian StyleZhang, Miao, Zhenyu Yang, Guofang Wu, Fafang Xu, Jianbo Zhang, Xuan Luo, Yuhong Ma, Huili Pang, Yaoke Duan, Jun Chen, and et al. 2024. "Effects of Probiotic-Fermented Feed on the Growth Profile, Immune Functions, and Intestinal Microbiota of Bamei Piglets" Animals 14, no. 4: 647. https://doi.org/10.3390/ani14040647
APA StyleZhang, M., Yang, Z., Wu, G., Xu, F., Zhang, J., Luo, X., Ma, Y., Pang, H., Duan, Y., Chen, J., Cai, Y., Wang, L., & Tan, Z. (2024). Effects of Probiotic-Fermented Feed on the Growth Profile, Immune Functions, and Intestinal Microbiota of Bamei Piglets. Animals, 14(4), 647. https://doi.org/10.3390/ani14040647