
Buffalo bbu-miR-493-5p Promotes Myoblast Proliferation and Differentiation

 ,
,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Sample Sources
2.2. Library Construction and sRNA Sequencing
2.3. Treatment of Raw Sequencing Data
2.4. Identification and Analysis of miRNAs
2.5. GO Enrichment Analyses
2.6. Synthesis and Primer Design of miRNA Mimic
2.7. Synthesized cDNA and qRT-PCR Assays
2.8. Buffalo Myoblast Isolation and Culture
2.9. CCK-8 Analysis
2.10. EdU Analysis
2.11. Western Blotting
2.12. Immunofluorescence Staining (IF)
2.13. Fluorescence In Situ Hybridization (FISH)
2.14. Statistical Analysis
3. Result
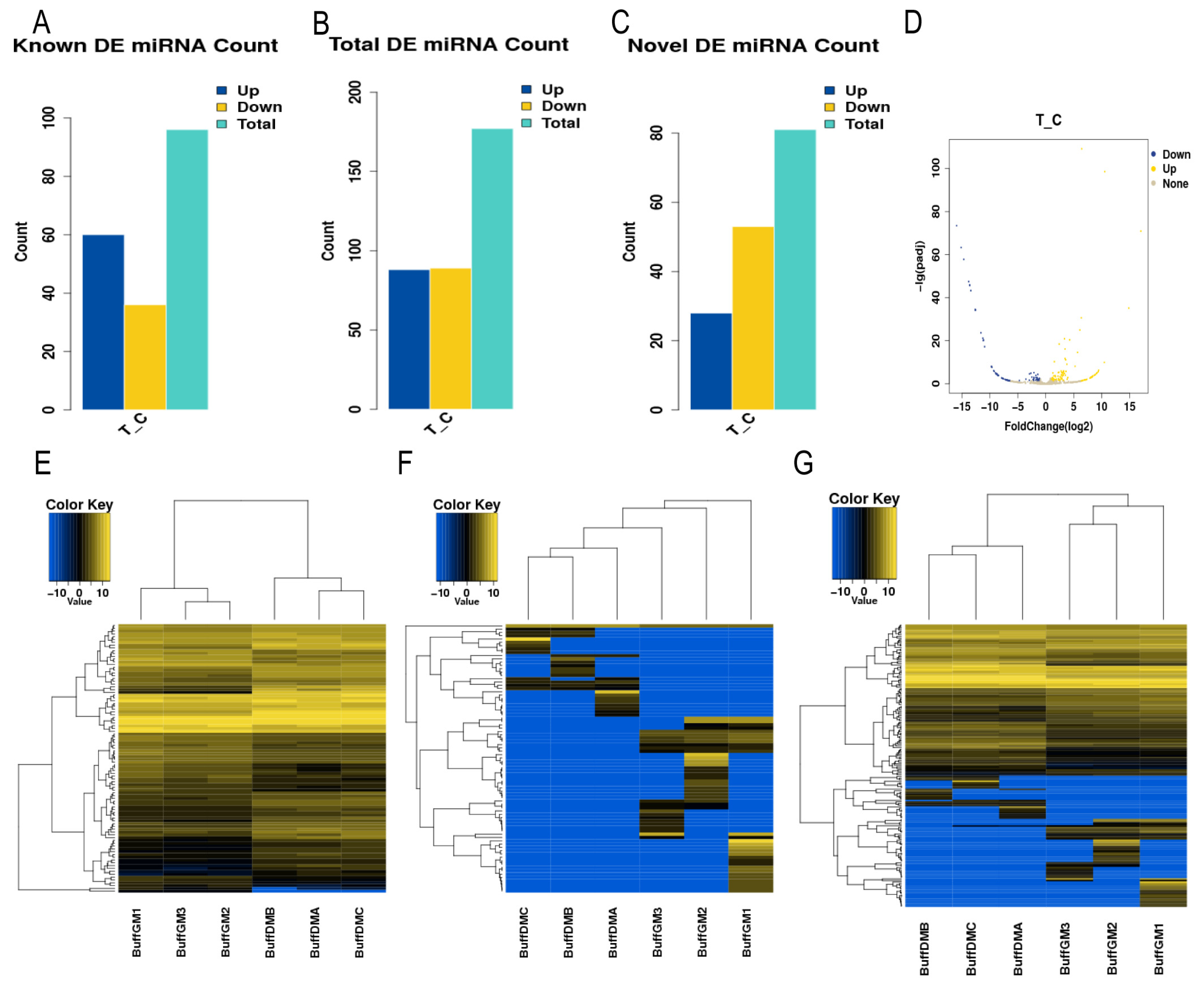
3.1. Identification of miRNAs Associated with Buffalo Myoblast Development
3.2. Screening of Differentially Expressed miRNAs
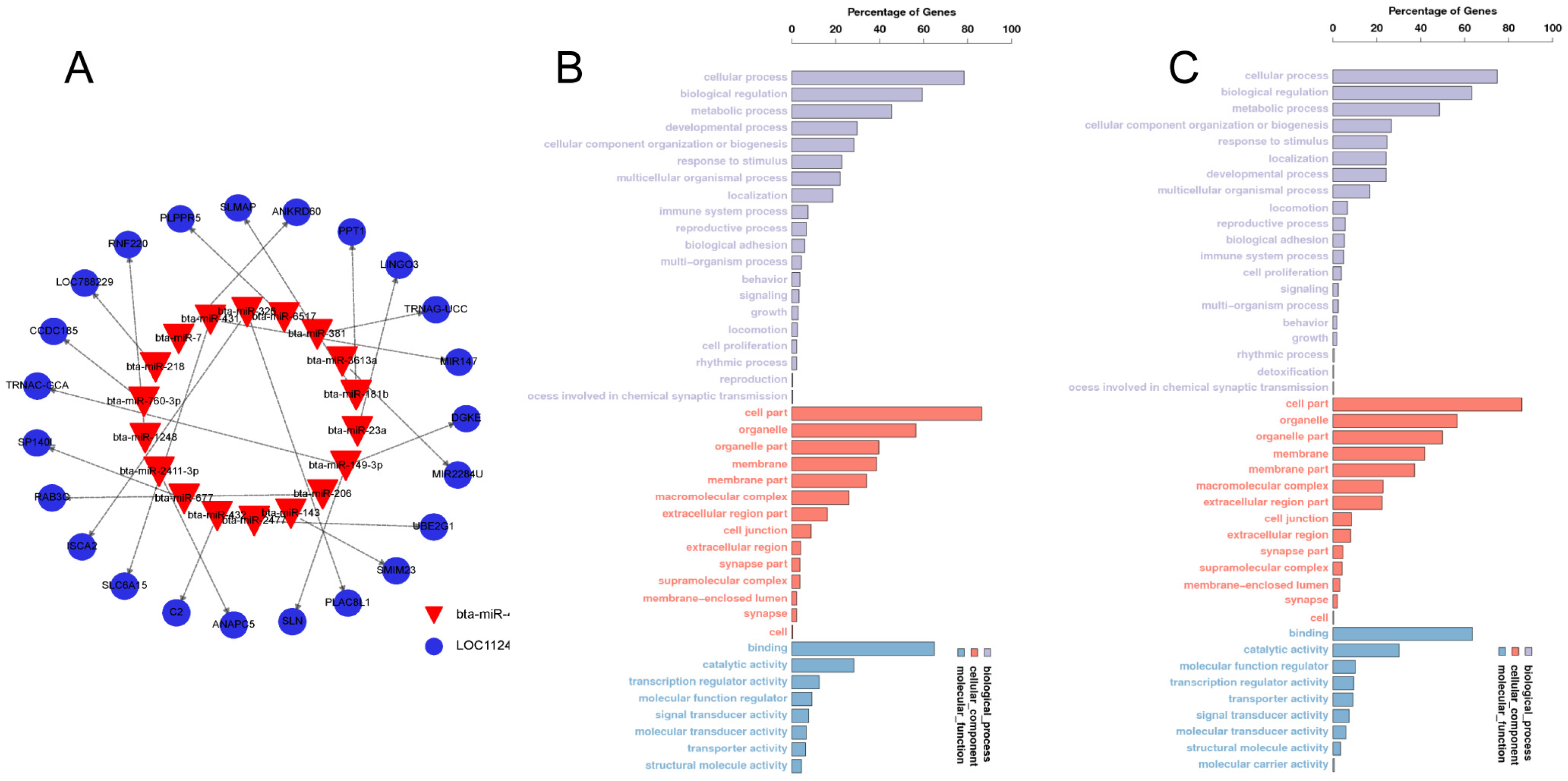
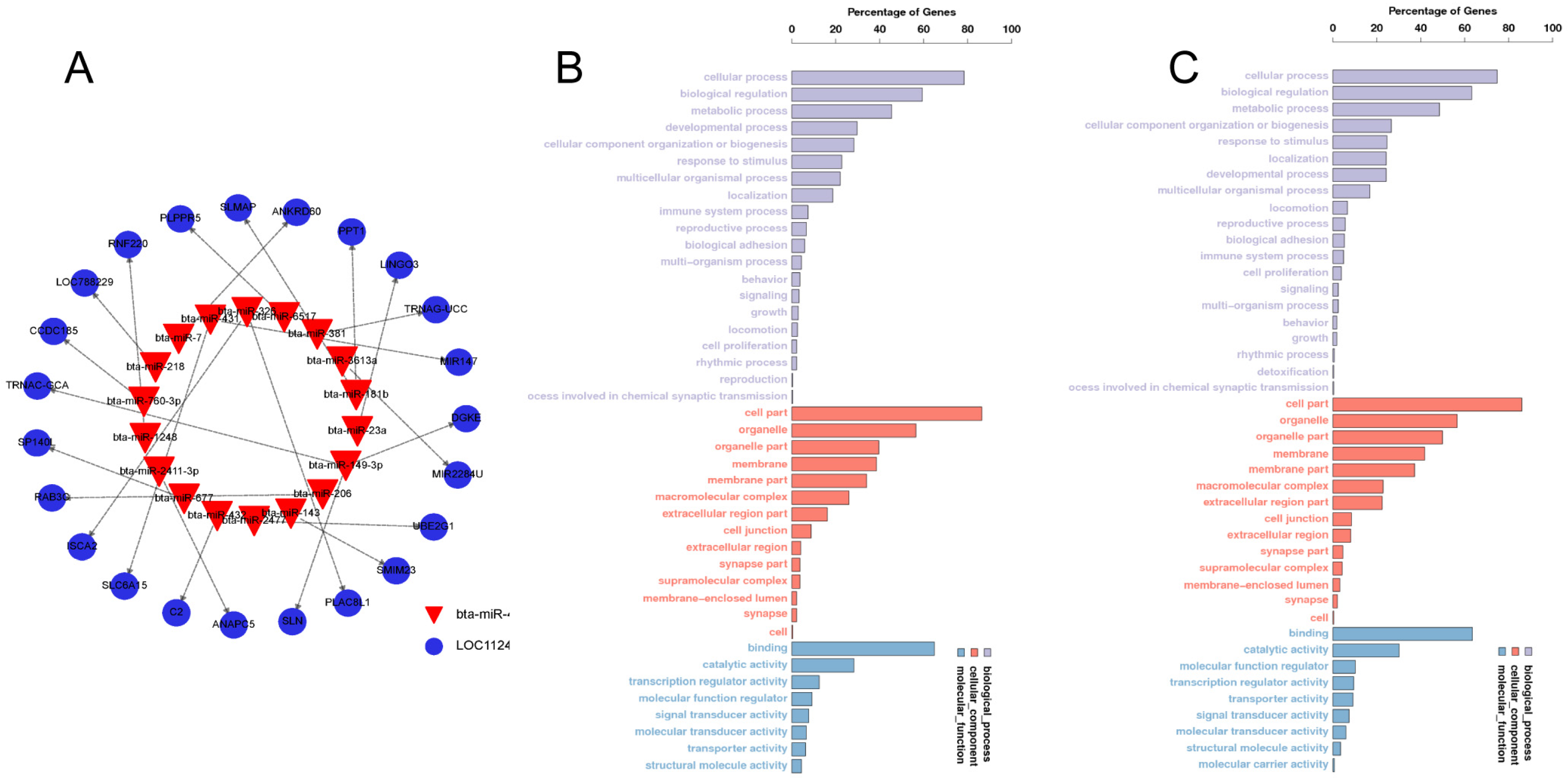
3.3. Candidate Target Gene Prediction and GO Analysis
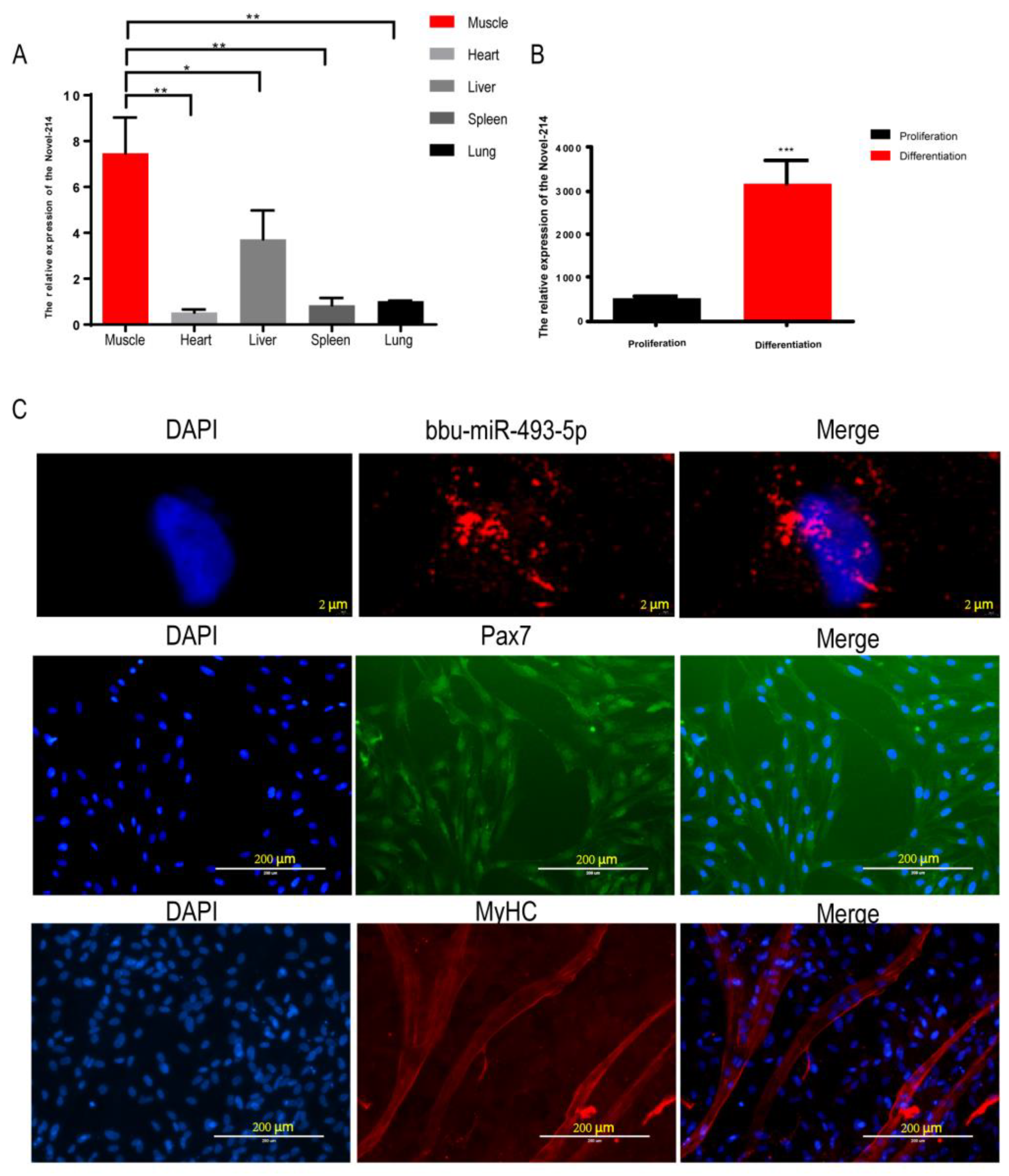
3.4. Identification of bbu-miR-493-5p as a Candidate miRNA
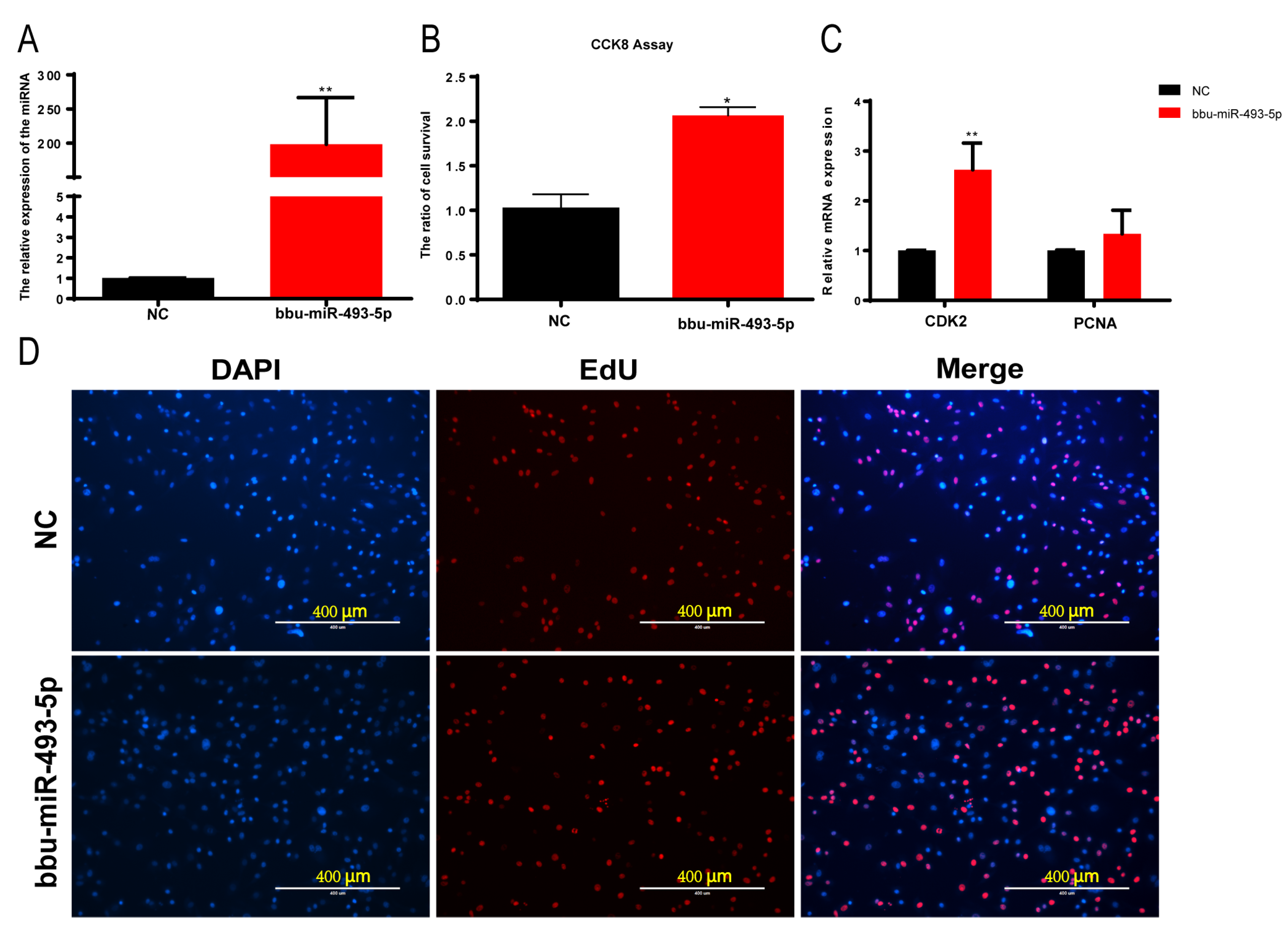
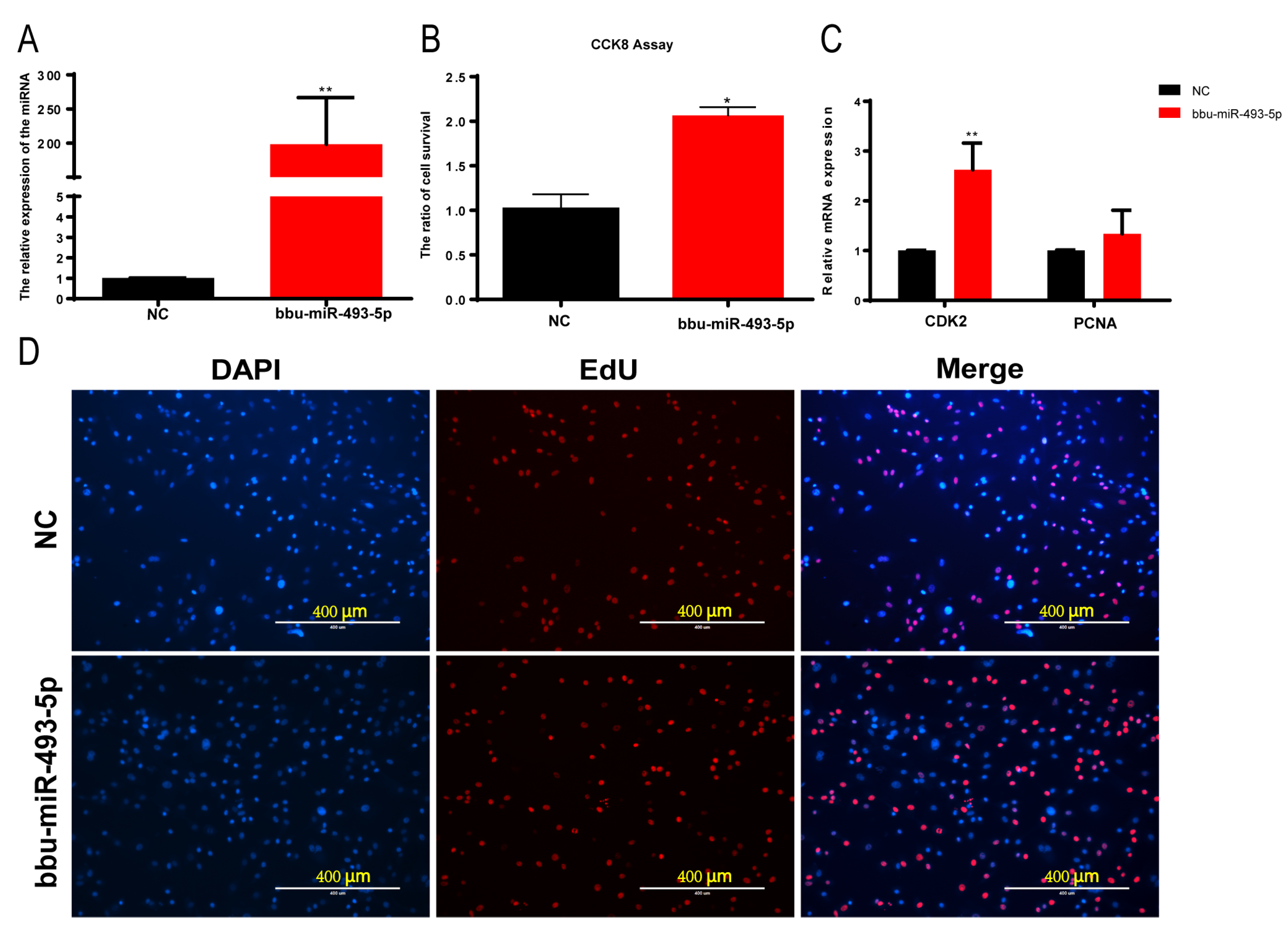
3.5. Effect of bbu-miR-493-5p on the Proliferation of Buffalo Myoblasts
3.6. Effect of Buffalo Myoblast Differentiation by bbu-miR-493-5p
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hernández-Castellano, L.E.; Nally, J.E.; Lindahl, J.; Wanapat, M.; Alhidary, I.A.; Fangueiro, D.; Grace, D.; Ratto, M.; Bambou, J.C.; De Almeida, A.M. Dairy science and health in the tropics: Challenges and opportunities for the next decades. Trop. Anim. Health Prod. 2019, 51, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Huang, K.; Wang, P.; Feng, T.; Shi, D.; Cui, K.; Luo, C.; Shafique, L.; Qian, Q.; Ruan, J.; et al. Comparison of long Non-Coding RNA expression profiles of cattle and buffalo differing in muscle characteristics. Front. Genet. 2020, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, M. Myogenic progenitor cells and skeletal myogenesis in vertebrates. Curr. Opin. Genet. Dev. 2006, 16, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Nie, M.; Deng, Z.; Liu, J.; Wang, D. Noncoding RNAs, emerging regulators of skeletal muscle development and diseases. BioMed Res. Int. 2015, 2015, 676575. [Google Scholar] [CrossRef]
- Mohammadabadi, M.; Bordbar, F.; Jensen, J.; Du, M.; Guo, W. Key genes regulating skeletal muscle development and growth in farm animals. Animals 2021, 11, 835. [Google Scholar] [CrossRef]
- Nederveen, J.P.; Fortino, S.A.; Baker, J.M.; Snijders, T.; Joanisse, S.; McGlory, C.; McKay, B.R.; Kumbhare, D.; Parise, G. Consistent expression pattern of myogenic regulatory factors in whole muscle and isolated human muscle satellite cells after eccentric contractions in humans. J. Appl. Physiol. 2019, 127, 1419–1426. [Google Scholar] [CrossRef]
- Liu, N.; Nelson, B.R.; Bezprozvannaya, S.; Shelton, J.M.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. Requirement of MEF2A, C, and D for skeletal muscle regeneration. Proc. Natl. Acad. Sci. USA 2014, 111, 4109–4114. [Google Scholar] [CrossRef]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef]
- Zhang, N.; Mendieta-Esteban, J.; Magli, A.; Lilja, K.C.; Perlingeiro, R.C.R.; Marti-Renom, M.A.; Tsirigos, A.; Dynlacht, B.D. Muscle progenitor specification and myogenic differentiation are associated with changes in chromatin topology. Nat. Commun. 2020, 11, 6222. [Google Scholar] [CrossRef]
- Callis, T.E.; Chen, J.; Wang, D. MicroRNAs in skeletal and cardiac muscle development. DNA Cell Biol. 2007, 26, 219–225. [Google Scholar] [CrossRef]
- Williams, A.H.; Liu, N.; van Rooij, E.; Olson, E.N. MicroRNA control of muscle development and disease. Curr. Opin. Cell Biol. 2009, 21, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Elsaeid Elnour, I.; Dong, D.; Wang, X.; Zhansaya, T.; Khan, R.; Jian, W.; Jie, C.; Chen, H. Bta-miR-885 promotes proliferation and inhibits differentiation of myoblasts by targeting MyoD1. J. Cell. Physiol. 2020, 235, 6625–6636. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Chen, J. MicroRNAs in skeletal myogenesis. Cell Cycle 2011, 10, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Güller, I.; Russell, A.P. MicroRNAs in skeletal muscle: Their role and regulation in development, disease and function. J. Physiol. 2010, 588 Pt 21, 4075–4087. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.J. The MyomiR network in skeletal muscle plasticity. Exerc. Sport Sci. Rev. 2011, 39, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, S.; Feng, X.; Liu, X.; Zhao, J.; Zheng, Q.; Wei, X.; Ma, Y. MiRNA transcriptome comparison between muscle and adipose tissues indi cates potential miRNAs associated with intramuscular fat in Chinese sw amp buffalo. Genome 2019, 62, 729–738. [Google Scholar] [CrossRef]
- Hao, D.; Wang, X.; Wang, X.; Thomsen, B.; Yang, Y.; Lan, X.; Huang, Y.; Chen, H. MicroRNA bta-miR-365-3p inhibits proliferation but promotes differentiation of primary bovine myoblasts by targeting the activin a receptor type I. J. Anim. Sci. Biotechnol. 2021, 12, 16. [Google Scholar] [CrossRef]
- Cacchiarelli, D.; Martone, J.; Girardi, E.; Cesana, M.; Incitti, T.; Morlando, M.; Nicoletti, C.; Santini, T.; Sthandier, O.; Barberi, L.; et al. MicroRNAs involved in molecular circuitries relevant for the Duchenne muscular dystrophy pathogenesis are controlled by the dystrophin/nNOS pathway. Cell Metab. 2010, 12, 341–351. [Google Scholar] [CrossRef]
- Wang, G.; Fang, X.; Han, M.; Wang, X.; Huang, Q. MicroRNA-493-5p promotes apoptosis and suppresses proliferation and in vasion in liver cancer cells by targeting VAMP2. Int. J. Mol. Med. 2018, 41, 1740–1748. [Google Scholar] [CrossRef]
- Yasukawa, K.; Liew, L.C.; Hagiwara, K.; Hironaka-Mitsuhashi, A.; Qin, X.; Furutani, Y.; Tanaka, Y.; Nakagama, H.; Kojima, S.; Kato, T.; et al. MicroRNA-493-5p-mediated repression of the MYCN oncogene inhibits hepa tic cancer cell growth and invasion. Cancer Sci. 2020, 111, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.; Cai, H.; Jiang, H.; Li, W.; Shi, Y. Long coding RNA CCAT2 enhances the proliferation and epithelial-mesenc hymal transition of cervical carcinoma cells via the microRNA-493-5p/C REB1 axis. Bioengineered 2021, 12, 6264–6274. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yang, X.; Luo, Q.; Fu, D.; Li, H.; Zhang, P.; Tie, C. Circular RNA CSPP1 motivates renal cell carcinoma carcinogenesis and the Warburg effect by targeting RAC1 through microRNA-493-5p. Acta Biochim. Pol. 2023, 70, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, Y.; Ding, Y. Circular RNA circPRKDC promotes tumorigenesis of gastric cancer via mo dulating insulin receptor substrate 2 (IRS2) and mediating microRNA-49 3-5p. Bioengineered 2021, 12, 7631–7643. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Xie, F.; Lin, Z.; Luo, J.; Chen, T.; Xi, Q.; Zhang, Y.; Sun, J. Effect of miR-493-5p on proliferation and differentiation of myoblast by targeting ANKRD17. Cell Tissue Res. 2023, 393, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Oh, I.; Yoon, C.; Hur, J.; Kim, J.; Kim, T.; Lee, C.; Park, K.-W.; Chae, I.-H.; Oh, B.-H.; Park, Y.-B.; et al. Involvement of E-selectin in recruitment of endothelial progenitor cells and angiogenesis in ischemic muscle. Blood 2007, 110, 3891–3899. [Google Scholar] [CrossRef] [PubMed]
- Tidball, J.G. Mechanisms of muscle injury, repair, and regeneration. Compr. Physiol. 2011, 1, 2029–2062. [Google Scholar] [CrossRef]
- Lorant, D.E.; Patel, K.D.; McIntyre, T.M.; McEver, R.P.; Prescott, S.M.; Zimmerman, G.A. Coexpression of GMP-140 and PAF by endothelium stimulated by histamine or thrombin: A juxtacrine system for adhesion and activation of neutr ophils. J. Cell Biol. 1991, 115, 223–234. [Google Scholar] [CrossRef]
- Liang, Y.; Lu, Q.; Li, W.; Zhang, D.; Zhang, F.; Zou, Q.; Chen, L.; Tong, Y.; Liu, M.; Wang, S.; et al. Reactivation of tumour suppressor in breast cancer by enhancer switching through NamiRNA network. Nucleic Acids Res. 2021, 49, 8556–8572. [Google Scholar] [CrossRef]
- Ju, D.; Liang, Y.; Hou, G.; Zheng, W.; Zhang, G.; Dun, X.; Wei, D.; Yan, F.; Zhang, L.; Lai, D.; et al. FBP1/miR-24-1/enhancer axis activation blocks renal cell carcinoma progression via Warburg effect. Front. Oncol. 2022, 12, 928373. [Google Scholar] [CrossRef]
- Younger, S.T.; Corey, D.R. Transcriptional gene silencing in mammalian cells by miRNA mimics that target gene promoters. Nucleic Acids Res. 2011, 39, 5682–5691. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene or miRNA | Primer Sequences (5′-3′) |
|---|---|
| PCNA | F: TCCAGAACAAGAGTATAGC |
| R: TACAACAGCATCTCCAAT | |
| CDK2 | F: TTTGCTGAGATGGTGACCCG |
| R: TAACTCCTGGCCAAACCACC | |
| MyoD1 | F: CCCAAAGATTGCGCTTAAGTG |
| R: GTTCCTTCGCCTCTCCTACCT | |
| MyoG | F: CAAATCCACTCCCTGAAA |
| R: GCATAGGAAGAGATGAACA | |
| MyHC | F: TGCTCATCTCACCAAGTTCC |
| R: CACTCTTCACTCTCATGGACC | |
| β-actin | F: CATCCTGACCCTCAAGTA |
| R: CTCGTTGTAGAAGGTGTG | |
| U6 | F: CGCTTCGGCAGCACATATAC |
| R: AAATATGGAACGCTTCACGA | |
| bbu-miR-493-5p | F: GCGCTTGTACATGGTAGGCT |
| R: GTGCAGGGTCCGAGGT | |
| bbu-miR-493-5p | RT:GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAATGAA |
| Sample | BuffDMA | BuffDMB | BuffDMC | BuffGM1 | BuffGM2 | BuffGM3 |
|---|---|---|---|---|---|---|
| Total Reads | 39,433,571 | 31,105,328 | 29,381,808 | 22,246,677 | 27,537,804 | 27,144,870 |
| Perfect Match Reads | 24,809,247 | 20,963,513 | 20,162,103 | 14,376,236 | 17,860,587 | 18,660,464 |
| Perfect Match Rate (%) | 62.91 | 67.4 | 68.62 | 64.62 | 64.86 | 68.74 |
| Not Match Rate (%) | 37.09 | 32.6 | 31.38 | 35.38 | 35.14 | 31.26 |
| Total Clean Reads Mature | 15,533,679 | 14,335,037 | 12,950,856 | 10,562,218 | 12,981,215 | 14,242,873 |
| Known miRNA Number | 597 | 593 | 587 | 620 | 609 | 592 |
| Novel miRNA Number | 40 | 52 | 51 | 80 | 66 | 59 |
| Total Clean Reads Novel | 16,382 | 6433 | 43,547 | 43,552 | 36,879 | 16,242 |
| miRNA | Sequence | Comparison Rate |
|---|---|---|
| bbu-miR-493-5p | UUGUACAUGGUAGGCUUUCAUU | 100% |
| hsa-miR-493-5p | UUGUACAUGGUAGGCUUUCAUU | 100% |
| chi-miR-493-5p | UUGUACAUGGUAGGCUUUCAUU | 100% |
| rno-miR-493-5p | UUGUACAUGGUAGGCUUUCAUU | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Zhong, D.; Yao, C.; Liu, Q.; Shi, D.; Jiang, M.; Wang, J.; Xiong, Z.; Li, H. Buffalo bbu-miR-493-5p Promotes Myoblast Proliferation and Differentiation. Animals 2024, 14, 533. https://doi.org/10.3390/ani14040533
Zhang L, Zhong D, Yao C, Liu Q, Shi D, Jiang M, Wang J, Xiong Z, Li H. Buffalo bbu-miR-493-5p Promotes Myoblast Proliferation and Differentiation. Animals. 2024; 14(4):533. https://doi.org/10.3390/ani14040533
Chicago/Turabian StyleZhang, Liyin, Dandan Zhong, Chengxuan Yao, Qingyou Liu, Deshun Shi, Mingsheng Jiang, Jian Wang, Zhaocheng Xiong, and Hui Li. 2024. "Buffalo bbu-miR-493-5p Promotes Myoblast Proliferation and Differentiation" Animals 14, no. 4: 533. https://doi.org/10.3390/ani14040533
APA StyleZhang, L., Zhong, D., Yao, C., Liu, Q., Shi, D., Jiang, M., Wang, J., Xiong, Z., & Li, H. (2024). Buffalo bbu-miR-493-5p Promotes Myoblast Proliferation and Differentiation. Animals, 14(4), 533. https://doi.org/10.3390/ani14040533