


Lactobacillus johnsonii Improves Intestinal Barrier Function and Reduces Post-Weaning Diarrhea in Piglets: Involvement of the Endocannabinoid System
 ,
,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Reagents
2.2. Animals and Diets
2.3. Diarrhea Index and Growth Performance
2.4. Reverse Transcription-PCR
2.5. L. johnsonii Absolute Quantification
2.6. Statistical Analysis
3. Results
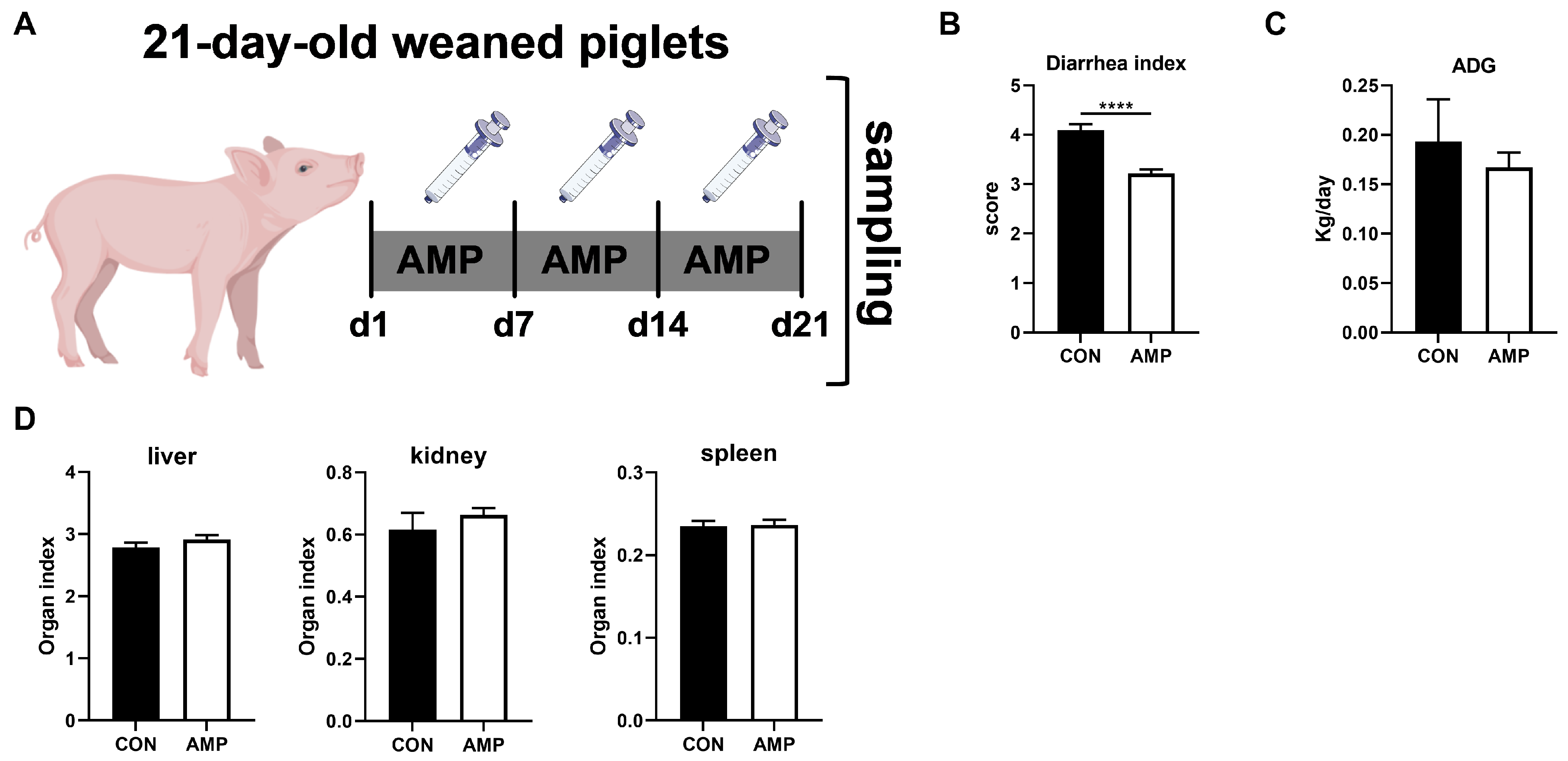
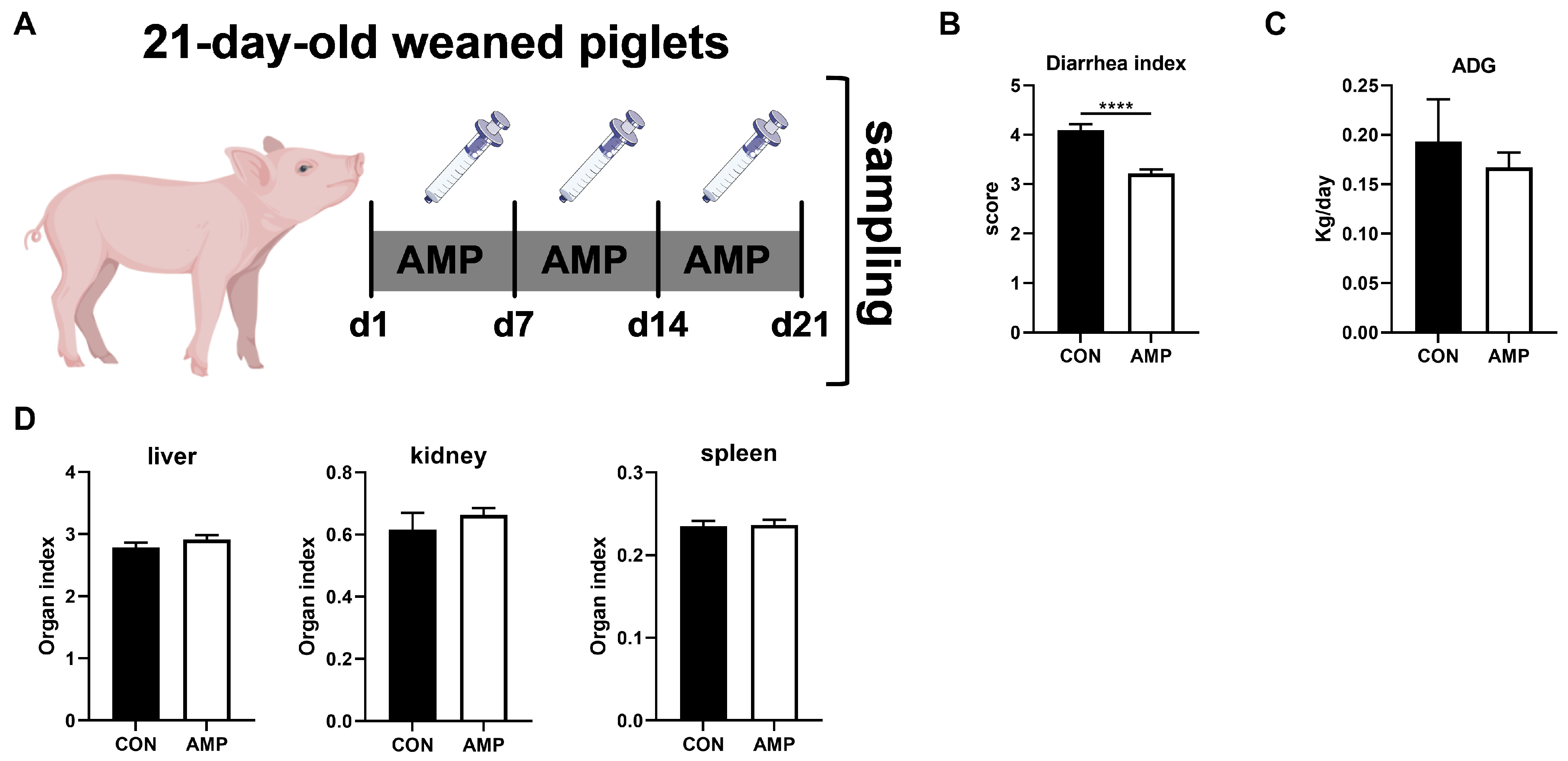
3.1. AMP Treatment Reduced Diarrhea in Weaned Piglets
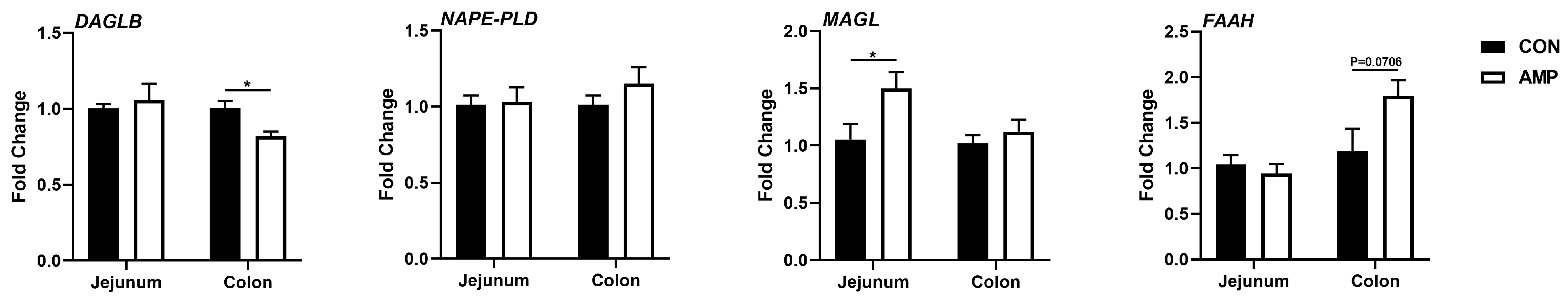
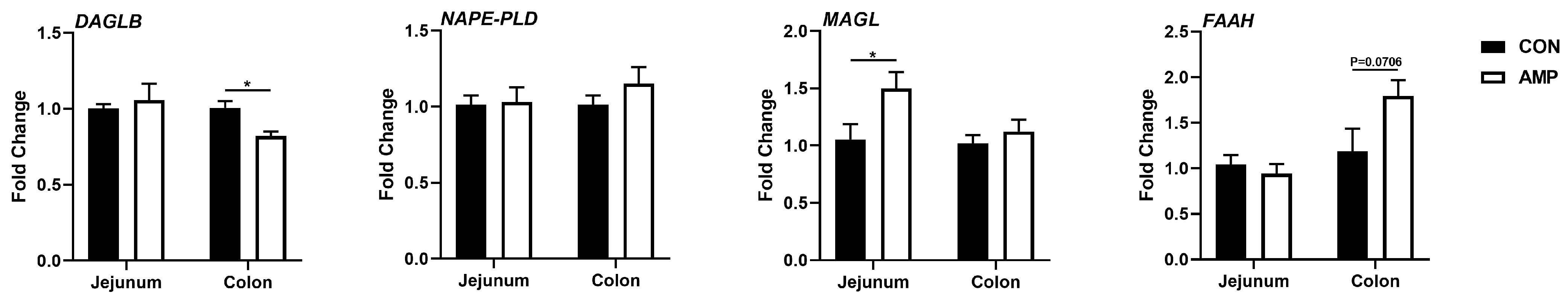
3.2. AMP Treatment Downregulated the Expression of DAGLB
3.3. L. johnsonii Was Correlated with the Expression of DAGLB
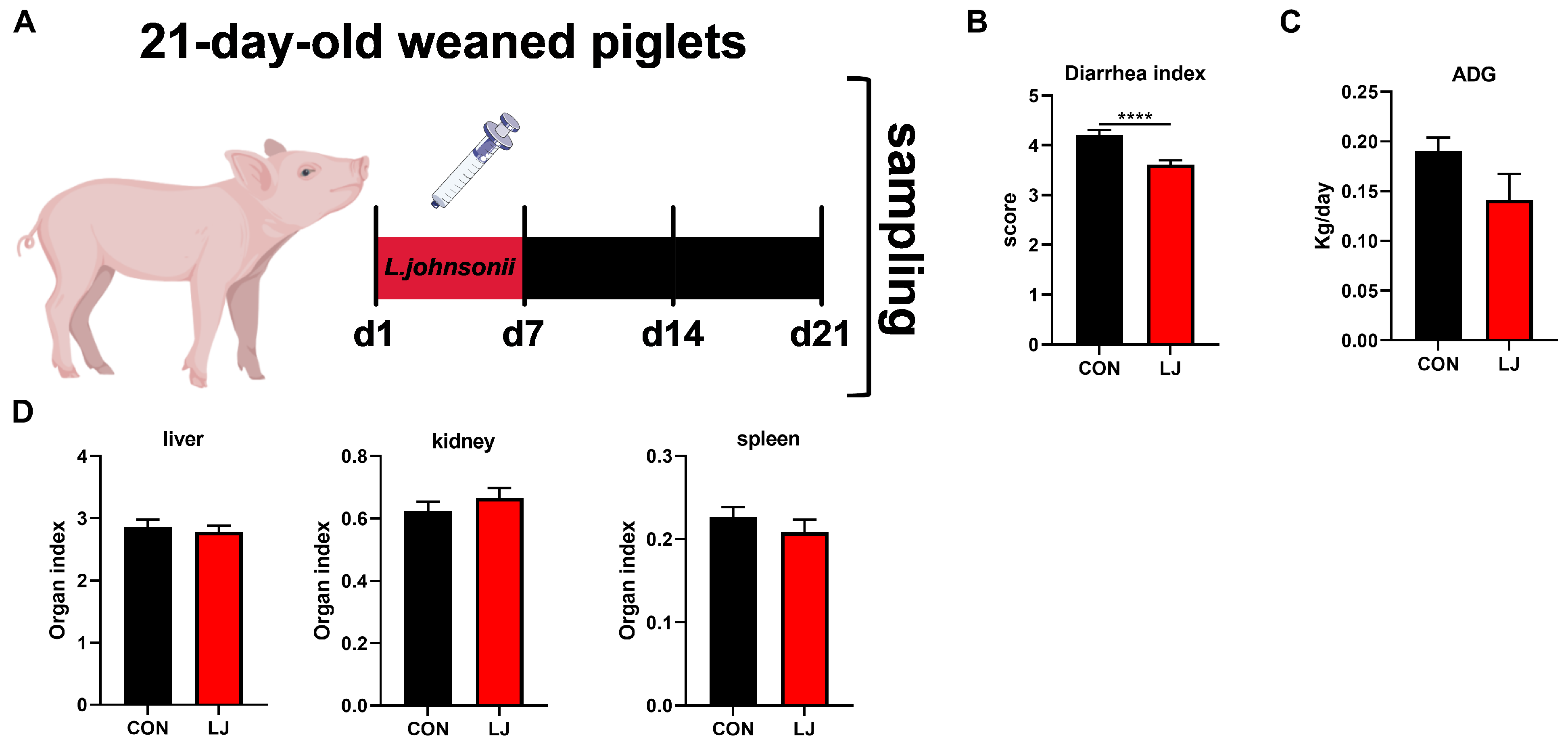
3.4. Dietary L. johnsonii Reduced Diarrhea in Weaned Piglets
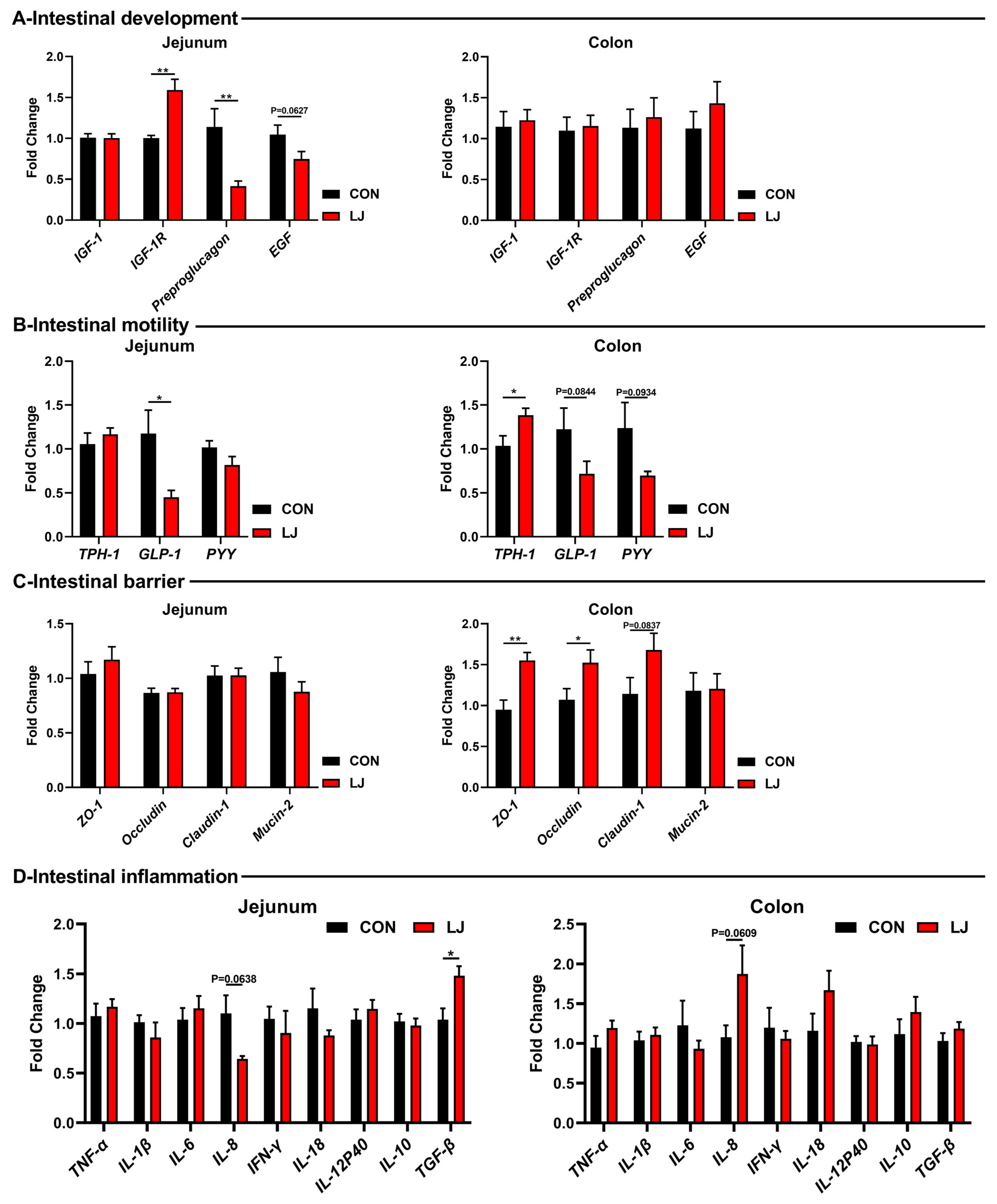
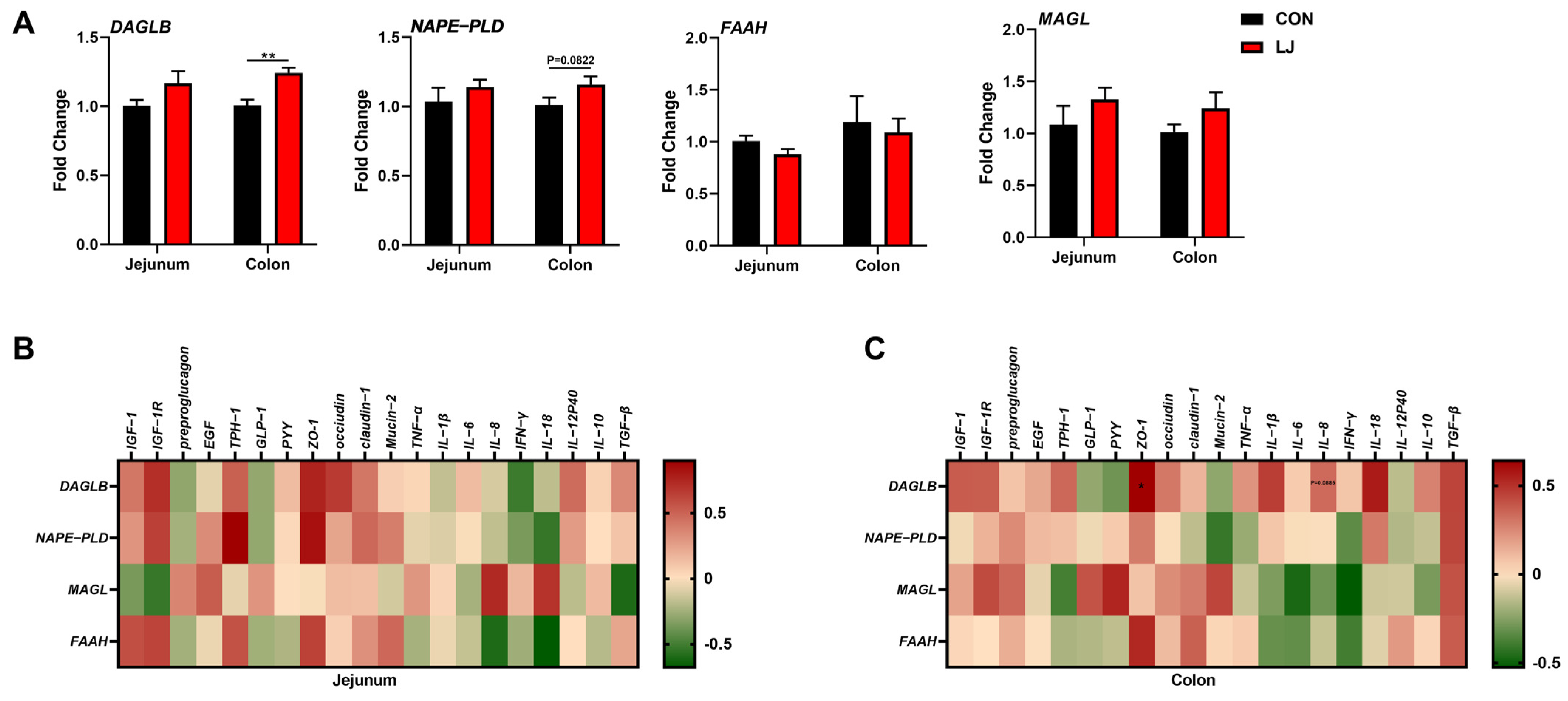
3.5. Dietary L. johnsonii Altered the Expressions of Intestinal Health-Related Genes
3.6. Dietary L. johnsonii-Mediated ECS Reprogramming Was Related to the Intestinal Barrier Function of Piglets
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bonetti, A.; Tugnoli, B.; Piva, A.; Grilli, E. Towards Zero Zinc Oxide: Feeding Strategies to Manage Post-Weaning Diarrhea in Piglets. Animals 2021, 11, 642. [Google Scholar] [CrossRef]
- van Beers-Schreurs, H.M.; Vellenga, L.; Wensing, T.; Breukink, H.J. The pathogenesis of the post-weaning syndrome in weaned piglets: A review. Vet. Q. 1992, 14, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; He, L.; Yang, C.; He, X.; Chen, H.; Feng, Y.; Tang, W.; Li, J.; Liu, D.; Li, T. Crosstalk between trace elements and T-cell immunity during early-life health in pigs. Sci. China Life Sci. 2023, 66, 1994–2005. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Ma, L.; Nie, Y.; Chen, J.; Zheng, W.; Wang, X.; Xie, C.; Zheng, Z.; Wang, Z.; Yang, T.; et al. A Microbiota-Derived Bacteriocin Targets the Host to Confer Diarrhea Resistance in Early-Weaned Piglets. Cell Host Microbe 2018, 24, 817–832.e818. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, B.; Nyachoti, C.M. Husbandry practices and gut health outcomes in weaned piglets: A review. Anim. Nutr. 2017, 3, 205–211. [Google Scholar] [CrossRef]
- Angelakis, E.; Merhej, V.; Raoult, D. Related actions of probiotics and antibiotics on gut microbiota and weight modification. Lancet Infect. Dis. 2013, 13, 889–899. [Google Scholar] [CrossRef]
- Robles-Jimenez, L.E.; Aranda-Aguirre, E.; Castelan-Ortega, O.A.; Shettino-Bermudez, B.S.; Ortiz-Salinas, R.; Miranda, M.; Li, X.; Angeles-Hernandez, J.C.; Vargas-Bello-Pérez, E.; Gonzalez-Ronquillo, M. Worldwide Traceability of Antibiotic Residues from Livestock in Wastewater and Soil: A Systematic Review. Animals 2021, 12, 60. [Google Scholar] [CrossRef]
- Jechalke, S.; Heuer, H.; Siemens, J.; Amelung, W.; Smalla, K. Fate and effects of veterinary antibiotics in soil. Trends Microbiol. 2014, 22, 536–545. [Google Scholar] [CrossRef]
- Casewell, M. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef]
- Fassarella, M.; Blaak, E.E.; Penders, J.; Nauta, A.; Smidt, H.; Zoetendal, E.G. Gut microbiome stability and resilience: Elucidating the response to perturbations in order to modulate gut health. Gut 2021, 70, 595–605. [Google Scholar] [CrossRef]
- Schroeder, B.O.; Bäckhed, F. Signals from the gut microbiota to distant organs in physiology and disease. Nat. Med. 2016, 22, 1079–1089. [Google Scholar] [CrossRef]
- Helmink, B.A.; Khan, M.A.W.; Hermann, A.; Gopalakrishnan, V.; Wargo, J.A. The microbiome, cancer, and cancer therapy. Nat. Med. 2019, 25, 377–388. [Google Scholar] [CrossRef]
- Huang, W.; Ma, T.; Liu, Y.; Kwok, L.-Y.; Li, Y.; Jin, H.; Zhao, F.; Shen, X.; Shi, X.; Sun, Z.; et al. Spraying compound probiotics improves growth performance and immunity and modulates gut microbiota and blood metabolites of suckling piglets. Sci. China Life Sci. 2022, 66, 1092–1107. [Google Scholar] [CrossRef]
- Cheng, C.S.; Wei, H.K.; Wang, P.; Yu, H.C.; Zhang, X.M.; Jiang, S.W.; Peng, J. Early intervention with faecal microbiota transplantation: An effective means to improve growth performance and the intestinal development of suckling piglets. Animal 2019, 13, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Zhao, F.; Zhu, W.; Wang, J. Effects of early-life lactoferrin intervention on growth performance, small intestinal function and gut microbiota in suckling piglets. Food Funct. 2019, 10, 5361–5373. [Google Scholar] [CrossRef]
- Ren, W.; Yu, B.; Yu, J.; Zheng, P.; Huang, Z.; Luo, J.; Mao, X.; He, J.; Yan, H.; Wu, J.; et al. Lower abundance of Bacteroides and metabolic dysfunction are highly associated with the post-weaning diarrhea in piglets. Sci. China Life Sci. 2022, 65, 2062–2075. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Hu, C.; Tang, W.; Azad, M.A.K.; Zhu, Q.; He, Q.; Kong, X. The Enhancement of Intestinal Immunity in Offspring Piglets by Maternal Probiotic or Synbiotic Supplementation Is Associated With the Alteration of Gut Microbiota. Front. Nutr. 2021, 8, 686053. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.; Wu, X.; Pan, Y.; Wang, L.; Cui, C.; Guo, Y.; Zhu, L.; Peng, J.; Wei, H. Early-Life Intervention Using Fecal Microbiota Combined with Probiotics Promotes Gut Microbiota Maturation, Regulates Immune System Development, and Alleviates Weaning Stress in Piglets. Int. J. Mol. Sci. 2020, 21, 503. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, Y.; Tian, Y.; Zhou, F.; Ma, J.; Xia, S.; Yang, T.; Ma, L.; Zeng, Q.; Liu, G.; et al. Obese Ningxiang pig-derived microbiota rewires carnitine metabolism to promote muscle fatty acid deposition in lean DLY pigs. Innovation 2023, 4, 100486. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zhou, M.; Song, Z.; Deng, Y.; Xia, S.; Li, Y.; Huang, X.; Xiao, D.; Yin, Y.; Yin, J. Clec7a drives gut fungus-mediated host lipid deposition. Microbiome 2023, 11, 264. [Google Scholar] [CrossRef]
- Zhen, Y.; Wang, Y.; He, F.; Chen, Y.; Hu, L.; Ge, L.; Wang, Y.; Wei, W.; Rahmat, A.; Loor, J.J.; et al. Homeostatic crosstalk among gut microbiome, hypothalamic and hepatic circadian clock oscillations, immunity and metabolism in response to different light–dark cycles: A multiomics study. J. Pineal Res. 2023, 75, e12892. [Google Scholar] [CrossRef]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, K.A.; Wiley, J.W. The Role of the Endocannabinoid System in the Brain-Gut Axis. Gastroenterology 2016, 151, 252–266. [Google Scholar] [CrossRef]
- Lu, H.C.; Mackie, K. Review of the Endocannabinoid System. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2021, 6, 607–615. [Google Scholar] [CrossRef]
- Cani, P.D.; Plovier, H.; Van Hul, M.; Geurts, L.; Delzenne, N.M.; Druart, C.; Everard, A. Endocannabinoids—At the crossroads between the gut microbiota and host metabolism. Nat. Rev. Endocrinol. 2015, 12, 133–143. [Google Scholar] [CrossRef]
- Li, Y.; Huang, X.; Yang, G.; Xu, K.; Yin, Y.; Brecchia, G.; Yin, J. CD36 favours fat sensing and transport to govern lipid metabolism. Prog. Lipid Res. 2022, 88, 101193. [Google Scholar] [CrossRef]
- Yin, J.; Li, Y.; Han, H.; Chen, S.; Gao, J.; Liu, G.; Wu, X.; Deng, J.; Yu, Q.; Huang, X.; et al. Melatonin reprogramming of gut microbiota improves lipid dysmetabolism in high-fat diet-fed mice. J. Pineal Res. 2018, 65, e12524. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, G.; Siopi, E.; Guenin-Mace, L.; Pascal, M.; Laval, T.; Rifflet, A.; Boneca, I.G.; Demangel, C.; Colsch, B.; Pruvost, A.; et al. Effect of gut microbiota on depressive-like behaviors in mice is mediated by the endocannabinoid system. Nat. Commun. 2020, 11, 6363. [Google Scholar] [CrossRef]
- Cota, D.; Marsicano, G.; Tschop, M.; Grubler, Y.; Flachskamm, C.; Schubert, M.; Auer, D.; Yassouridis, A.; Thone-Reineke, C.; Ortmann, S.; et al. The endogenous cannabinoid system affects energy balance via central orexigenic drive and peripheral lipogenesis. J. Clin. Investig. 2003, 112, 423–431. [Google Scholar] [CrossRef]
- Lacroix, S.; Pechereau, F.; Leblanc, N.; Boubertakh, B.; Houde, A.; Martin, C.; Flamand, N.; Silvestri, C.; Raymond, F.; Di Marzo, V.; et al. Rapid and Concomitant Gut Microbiota and Endocannabinoidome Response to Diet-Induced Obesity in Mice. mSystems 2019, 4, e00407-19. [Google Scholar] [CrossRef]
- Chen, M.; Hou, P.; Zhou, M.; Ren, Q.; Wang, X.; Huang, L.; Hui, S.; Yi, L.; Mi, M. Resveratrol attenuates high-fat diet-induced non-alcoholic steatohepatitis by maintaining gut barrier integrity and inhibiting gut inflammation through regulation of the endocannabinoid system. Clin. Nutr. 2020, 39, 1264–1275. [Google Scholar] [CrossRef]
- Song, M.; Liu, Y.; Soares, J.A.; Che, T.M.; Osuna, O.; Maddox, C.W.; Pettigrew, J.E. Dietary clays alleviate diarrhea of weaned pigs1. J. Anim. Sci. 2012, 90, 345–360. [Google Scholar] [CrossRef]
- Yu, L.; Li, H.; Peng, Z.; Ge, Y.; Liu, J.; Wang, T.; Wang, H.; Dong, L. Early Weaning Affects Liver Antioxidant Function in Piglets. Animals 2021, 11, 2679. [Google Scholar] [CrossRef]
- Jiao, J.; Wu, J.; Zhou, C.; Tang, S.; Wang, M.; Tan, Z. Composition of Ileal Bacterial Community in Grazing Goats Varies across Non-rumination, Transition and Rumination Stages of Life. Front. Microbiol. 2016, 7, 1364. [Google Scholar] [CrossRef]
- Pluske, J.R.; Turpin, D.L.; Kim, J.-C. Gastrointestinal tract (gut) health in the young pig. Anim. Nutr. 2018, 4, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, X.; Ma, Y.; Cai, S.; Yang, L.; Fan, Y.; Zeng, X.; Qiao, S. Lactobacillus reuteri improves the development and maturation of fecal microbiota in piglets through mother-to-infant microbe and metabolite vertical transmission. Microbiome 2022, 10, 211. [Google Scholar] [CrossRef]
- Durack, J.; Lynch, S.V. The gut microbiome: Relationships with disease and opportunities for therapy. J. Exp. Med. 2019, 216, 20–40. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Grasset, E.; Mannerås Holm, L.; Karsenty, G.; Macpherson, A.J.; Olofsson, L.E.; Bäckhed, F. Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proc. Natl. Acad. Sci. USA 2018, 115, 6458–6463. [Google Scholar] [CrossRef] [PubMed]
- Obata, Y.; Castaño, Á.; Boeing, S.; Bon-Frauches, A.C.; Fung, C.; Fallesen, T.; de Agüero, M.G.; Yilmaz, B.; Lopes, R.; Huseynova, A.; et al. Neuronal programming by microbiota regulates intestinal physiology. Nature 2020, 578, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Manca, C.; Boubertakh, B.; Leblanc, N.; Deschênes, T.; Lacroix, S.; Martin, C.; Houde, A.; Veilleux, A.; Flamand, N.; Muccioli, G.G.; et al. Germ-free mice exhibit profound gut microbiota-dependent alterations of intestinal endocannabinoidome signaling. J. Lipid Res. 2020, 61, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Guida, F.; Turco, F.; Iannotta, M.; De Gregorio, D.; Palumbo, I.; Sarnelli, G.; Furiano, A.; Napolitano, F.; Boccella, S.; Luongo, L.; et al. Antibiotic-induced microbiota perturbation causes gut endocannabinoidome changes, hippocampal neuroglial reorganization and depression in mice. Brain Behav. Immun. 2018, 67, 230–245. [Google Scholar] [CrossRef] [PubMed]
- Davoren, M.J.; Liu, J.; Castellanos, J.; Rodríguez-Malavé, N.I.; Schiestl, R.H. A novel probiotic, Lactobacillus johnsonii 456, resists acid and can persist in the human gut beyond the initial ingestion period. Gut Microbes 2018, 10, 458–480. [Google Scholar] [CrossRef]
- Zhao, K.; Yin, H.; Yan, H.; Tang, W.; Diao, H.; Wang, Q.; Qi, R.; Liu, J. Dietary Supplementation of Lactobacillus johnsonii RS-7 Improved Antioxidant and Immune Function of Weaned Piglets. Animals 2023, 13, 1595. [Google Scholar] [CrossRef]
- Rousseaux, C.; Thuru, X.; Gelot, A.; Barnich, N.; Neut, C.; Dubuquoy, L.; Dubuquoy, C.; Merour, E.; Geboes, K.; Chamaillard, M.; et al. Lactobacillus acidophilus modulates intestinal pain and induces opioid and cannabinoid receptors. Nat. Med. 2006, 13, 35–37. [Google Scholar] [CrossRef]
- Berg, D.; Clemente, J.C.; Colombel, J.-F. Can inflammatory bowel disease be permanently treated with short-term interventions on the microbiome? Expert. Rev. Gastroenterol. Hepatol. 2015, 9, 781–795. [Google Scholar] [CrossRef]
- Muccioli, G.G.; Naslain, D.; Bäckhed, F.; Reigstad, C.S.; Lambert, D.M.; Delzenne, N.M.; Cani, P.D. The endocannabinoid system links gut microbiota to adipogenesis. Mol. Syst. Biol. 2010, 6, 392. [Google Scholar] [CrossRef]
- Tomé, D.; Geurts, L.; Muccioli, G.G.; Delzenne, N.M.; Cani, P.D. Chronic Endocannabinoid System Stimulation Induces Muscle Macrophage and Lipid Accumulation in Type 2 Diabetic Mice Independently of Metabolic Endotoxaemia. PLoS ONE 2013, 8, e55963. [Google Scholar] [CrossRef]
- Alhamoruni, A.; Lee, A.C.; Wright, K.L.; Larvin, M.; O’Sullivan, S.E. Pharmacological Effects of Cannabinoids on the Caco-2 Cell Culture Model of Intestinal Permeability. J. Pharmacol. Exp. Ther. 2010, 335, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Xu, T.; Qian, M.; Zhang, Y.; Yang, Z.; Han, X. Faecal microbiota transplantation alleviates early-life antibiotic-induced gut microbiota dysbiosis and mucosa injuries in a neonatal piglet model. Microbiol. Res. 2022, 255, 126942. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) |
|---|---|---|
| β-actin | AGAGCGCAAGTACTCCGTGT | ACATCTGCTGGAAGGTGGAC |
| GAPDH | TCGGAGTGAACGGATTTGGC | TGACAAGCTTCCCGTTCTCC |
| FAAH | TGCCACCGTGCAAGAAAATG | CCACTGCCCTAACAACGACT |
| MAGL | CACTTCTCCGGCATGGTTCT | CGTGAAACGGCGTTGAGC |
| NAPE-PLD | ACCGGCCTCTGAGAAAATGG | AGGGTTAACTGGGGAGACCT |
| DAGLB | TTTGTAATCCCGGACCACGG | GACCTGCCGAGGAATACGGA |
| IGF-1 | TCACTGGAGGATGGAATACAGC | CCTGAACTCCCTCTACTTGTGTTC |
| IGF-1R | ATGGAGGAAGTGACAGGGACTA | GTGGTGGTGGAGGTGAAGTG |
| Preproglucagon | ACTCACAGGGCACGTTTACCA | AGGTCCCTTCAGCATGTCTCT |
| EGF | ATCTCAGGAATGGGAGTCAACC | TCACTGGAGGATGGAATACAGC |
| TPH-1 | TGGATCTGAACTGGATGCTG | CGGTTCCCCAGGTCTTAATC |
| GLP-1 | GACATGCTGAAGGGACCTTTAC | GGCCTTTCACCAGCCAC |
| PYY | AGATATGCTAATACACCGAT | CCAAACCCTTCTCAGATG |
| ZO-1 | GAGGATGGTCACACCGTGGT | GGAGGATGCTGTTGTCTCGG |
| Occludin | ATGCTTTCTCAGCCAGCGTA | AAGGTTCCATAGCCTCGGTC |
| Claudin-1 | GCATCATTTCCTCCCTGTT | TCTTGGCTTTGGGTGGTT |
| Mucin-2 | CTGTGTGGGGCCTGACAA | AGTGCTTGCAGTCGAACTCA |
| TNF-α | CCACGCTCTTCTGCCTACTGC | GCTGTCCCTCGGCTTTGAC |
| IL-1β | AGTGGAGAAGCCGATGAAGA | CATTGCACGTTTCAAGGATG |
| IL-6 | CCTCTCCGGACAAAACTGAA | TCTGCCAGTACCTCCTTGCT |
| IL-8 | TAGGACCAGAGCCAGGAAGA | AGCAGGAAAACTGCCAAGAA |
| IFN-γ | TCCAGCGCAAAGCCATCAGTG | ATGCTCTCTGGCCTTGGAACATAGT |
| IL-18 | TATGCCTGATTCTGACTGTT | ATGAAGACTCAAACTGTATCT |
| IL-12P40 | GATGCTGGCCAGTACACC | TCCAGCACGACCTCAATG |
| IL-10 | CTGCCTCCCACTTTCTCTTG | TCAAAGGGGCTCCCTAGTTT |
| TGF-β | GAAGATGCTTGGAGCTGAGG | TGGGACTTTGTCTTGGGAAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, Z.; Wang, K.; Liu, Y.; Li, Y.; He, F.; Yin, J.; Tang, W. Lactobacillus johnsonii Improves Intestinal Barrier Function and Reduces Post-Weaning Diarrhea in Piglets: Involvement of the Endocannabinoid System. Animals 2024, 14, 493. https://doi.org/10.3390/ani14030493
Yin Z, Wang K, Liu Y, Li Y, He F, Yin J, Tang W. Lactobacillus johnsonii Improves Intestinal Barrier Function and Reduces Post-Weaning Diarrhea in Piglets: Involvement of the Endocannabinoid System. Animals. 2024; 14(3):493. https://doi.org/10.3390/ani14030493
Chicago/Turabian StyleYin, Zhangzheng, Kaijun Wang, Yun Liu, Yunxia Li, Fang He, Jie Yin, and Wenjie Tang. 2024. "Lactobacillus johnsonii Improves Intestinal Barrier Function and Reduces Post-Weaning Diarrhea in Piglets: Involvement of the Endocannabinoid System" Animals 14, no. 3: 493. https://doi.org/10.3390/ani14030493
APA StyleYin, Z., Wang, K., Liu, Y., Li, Y., He, F., Yin, J., & Tang, W. (2024). Lactobacillus johnsonii Improves Intestinal Barrier Function and Reduces Post-Weaning Diarrhea in Piglets: Involvement of the Endocannabinoid System. Animals, 14(3), 493. https://doi.org/10.3390/ani14030493