
The Hippocampus in Pigeons Contributes to the Model-Based Valuation and the Relationship between Temporal Context States

 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
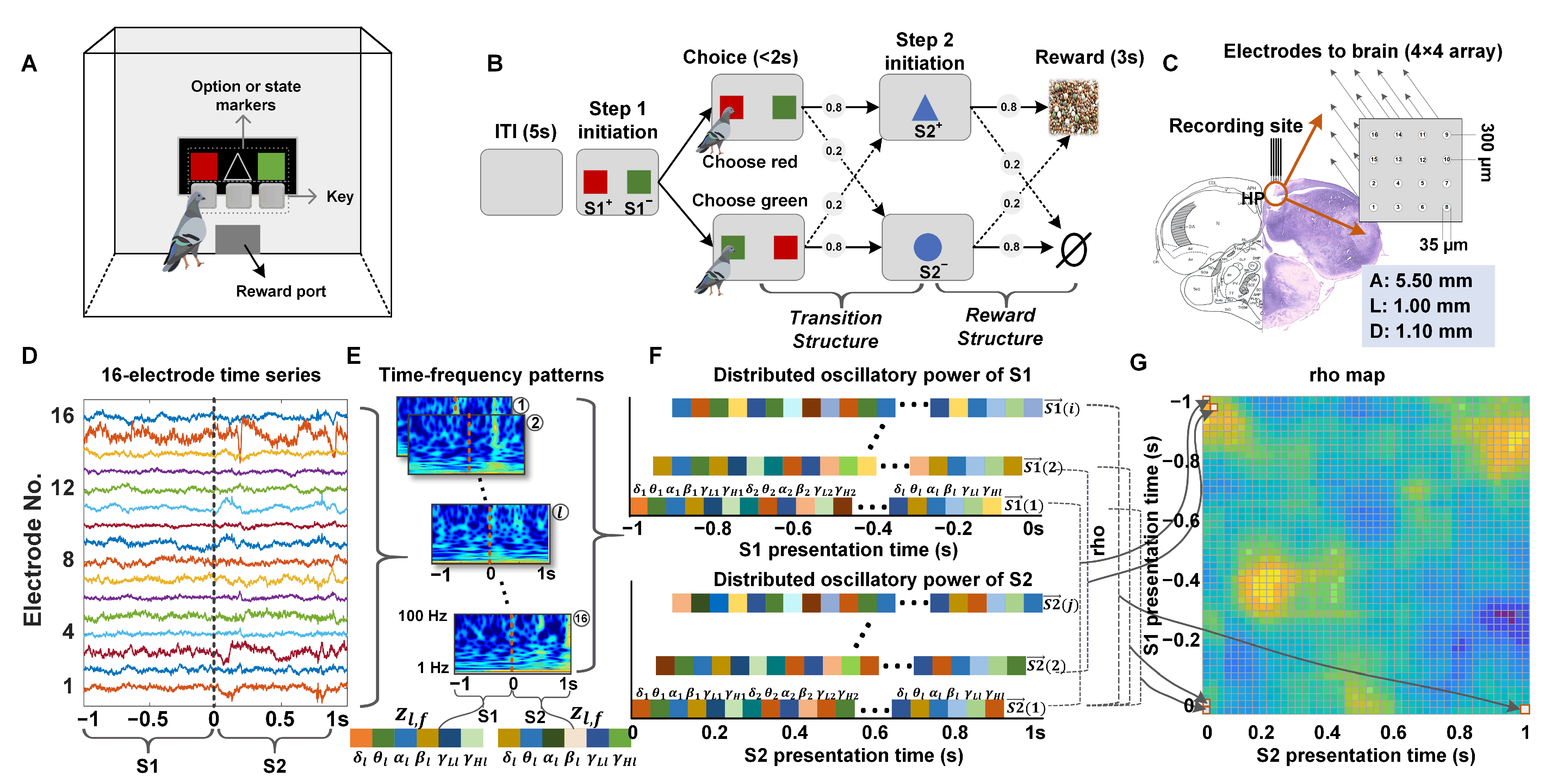
2.1. Experimental Procedure and Data Acquisition
2.1.1. Behavioral Task and Apparatus
2.1.2. Subjects and Surgery
2.1.3. Acquisition of Neurophysiological Data
2.1.4. Histological Reconstructions
2.2. Dynamic Reinforcement Learning Model for Pigeon Behaviors
2.3. LFP Analysis
2.3.1. Preprocessing of LFP Data
2.3.2. Time-Frequency Power Estimates of LFPs
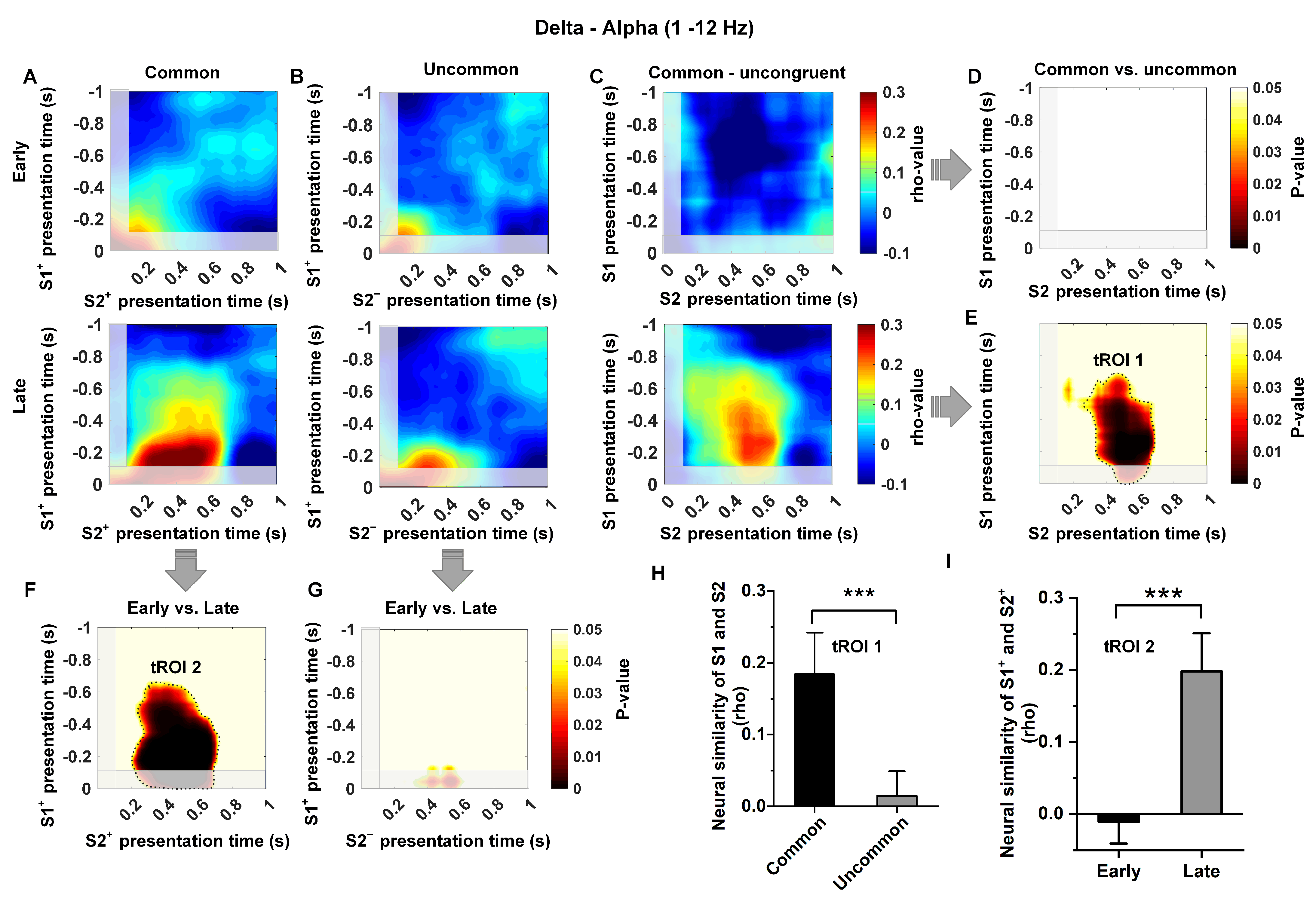
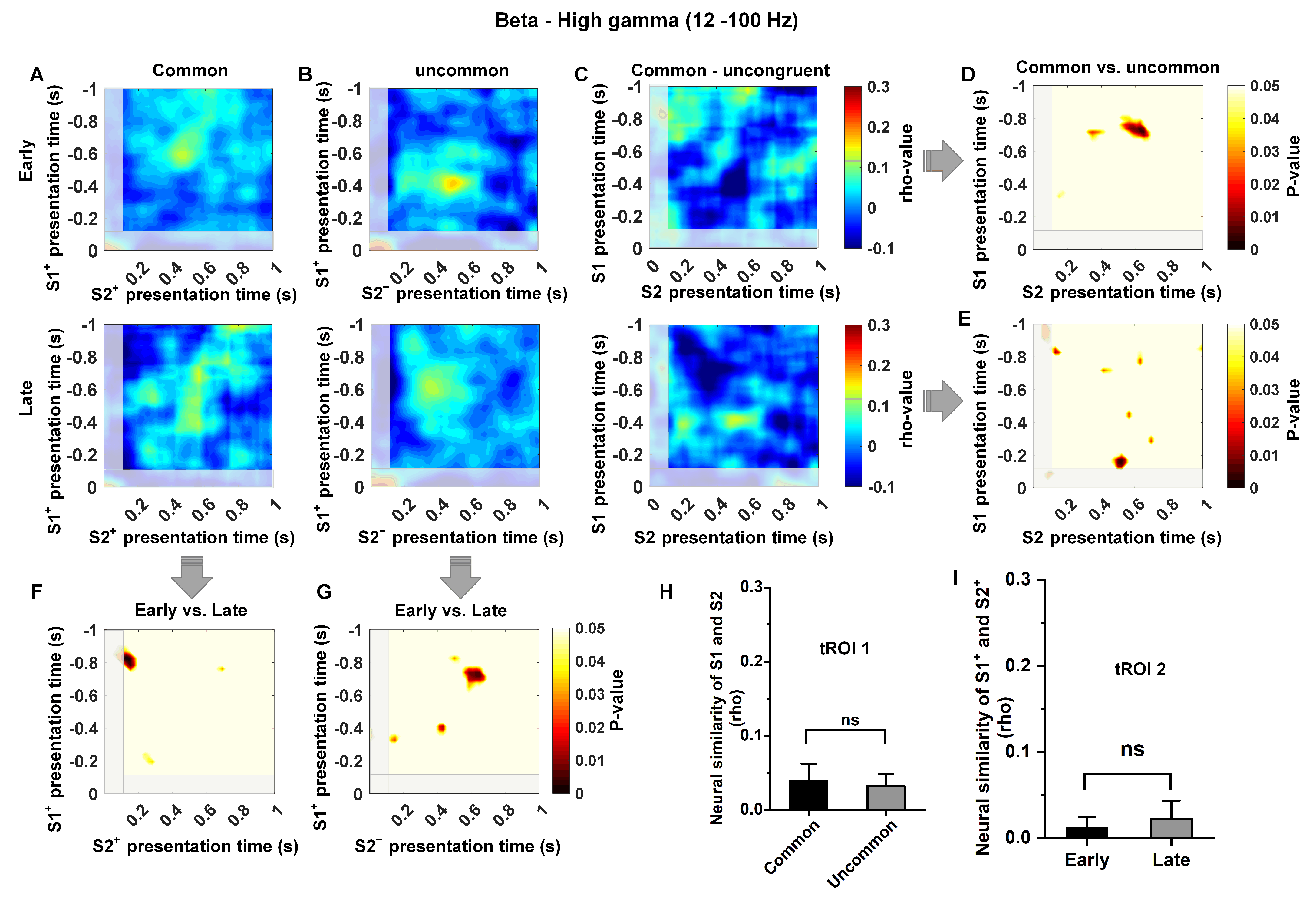
2.3.3. Analysis of Neural Similarity Representations in Temporal Context States
2.4. Correlation between Hippocampal LFPs Power and Model-Based Valuation
2.4.1. Regression Analysis
2.4.2. Behavior Choice Decoding
2.5. Statistical Analysis
3. Results
3.1. Behavioral Results
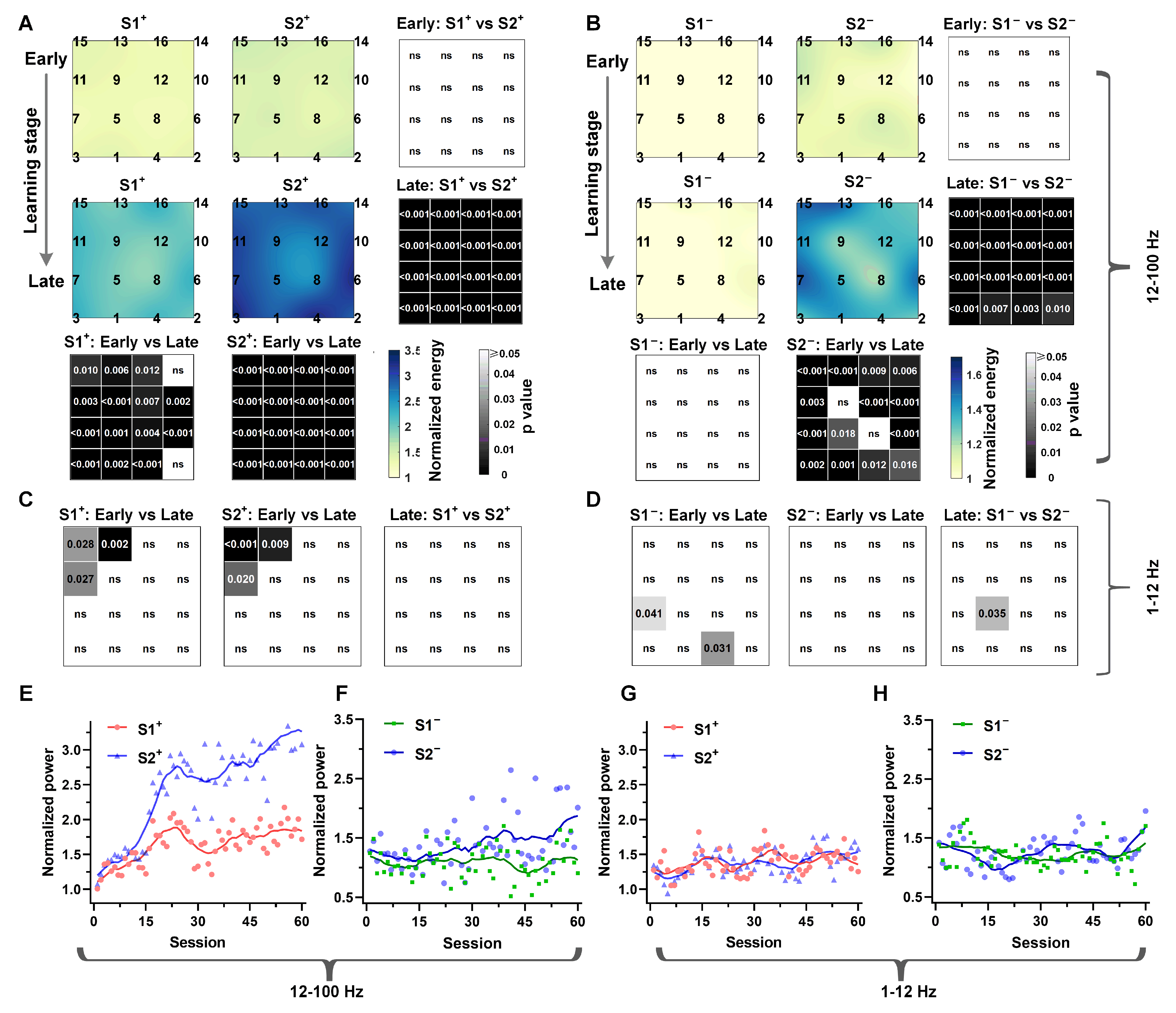
3.2. The Temporal Context States Modulate the Hippocampal LFPs High-Frequency Power
3.3. High-Frequency Power in the Hp Correlates of Model-Based Valuations
3.4. Low-Frequency Dependence of the Relationship between Temporal Context States
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, K.J.; Venditto, S.J.C. Multi-step planning in the brain. Curr. Opin. Behav. Sci. 2021, 38, 29–39. [Google Scholar] [CrossRef]
- Brunec, I.K.; Momennejad, I. Predictive representations in hippocampal and prefrontal hierarchies. J. Neurosci. 2022, 42, 299–312. [Google Scholar] [CrossRef]
- Stachenfeld, K.L.; Botvinick, M.M.; Gershman, S.J. The hippocampus as a predictive map. Nat. Neurosci. 2017, 20, 1643–1653. [Google Scholar] [CrossRef]
- Mehrotra, D.; Dubé, L. Accounting for multiscale processing in adaptive real-world decision-making via the hippocampus. Front. Neurosci. 2023, 17, 1200842. [Google Scholar] [CrossRef] [PubMed]
- Balleine, B.W.; O’Doherty, J.P. Human and rodent homologies in action control: Corticostriatal determinants of goal-directed and habitual action. Neuropsychopharmacology 2010, 35, 48–69. [Google Scholar] [CrossRef]
- Daw, N.D.; Niv, Y.; Dayan, P. Uncertainty-based competition between prefrontal and dorsolateral striatal systems for behavioral control. Nat. Neurosci. 2005, 8, 1704–1711. [Google Scholar] [CrossRef] [PubMed]
- Tolman, E.C. Cognitive maps in rats and men. Psychol. Rev. 1948, 55, 189–208. [Google Scholar] [CrossRef]
- Iigaya, K.; O’Doherty, J.P. Hippocampus is what happens while you’re busy making other plans. Neuron 2019, 102, 517–519. [Google Scholar] [CrossRef]
- Vikbladh, O.M.; Meager, M.R.; King, J.; Blackmon, K.; Devinsky, O.; Shohamy, D.; Burgess, N.; Daw, N.D. Hippocampal contributions to model-based planning and spatial memory. Neuron 2019, 102, 683–693.e4. [Google Scholar] [CrossRef]
- Miller, K.J.; Botvinick, M.M.; Brody, C.D. Dorsal hippocampus contributes to model-based planning. Nat. Neurosci. 2017, 20, 1269–1276. [Google Scholar] [CrossRef]
- George, T.M.; De Cothi, W.; Stachenfeld, K.L.; Barry, C. Rapid learning of predictive maps with SDTP and theta phase precession. eLife 2023, 12, e80663. [Google Scholar] [CrossRef]
- Morris, R.G.M.; Garrud, P.; Rawlins, J.N.P.; O’Keefe, J. Place navigation impaired in rats with hippocampal lesions. Nature 1982, 297, 681–683. [Google Scholar] [CrossRef]
- Packard, M.G.; McGaugh, J.L. Inactivation of hippocampus or caudate nucleus with lidocaine differentially affects expression of place and response learning. Neurobiol. Learn. Mem. 1996, 65, 65–72. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.; Dostrovsky, J. The Hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 1971, 34, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, B.E.; Foster, D.J. Hippocampal place-cell sequences depict future paths to remembered goals. Nature 2013, 497, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Wikenheiser, A.M.; Redish, A.D. Hippocampal theta sequences reflect current goals. Nat. Neurosci. 2015, 18, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Foster, D.J.; Knierim, J.J. Sequence learning and the role of the hippocampus in rodent navigation. Curr. Opin. Neurobiol. 2012, 22, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Koene, R.A.; Gorchetchnikov, A.; Cannon, R.C.; Hasselmo, M.E. Modeling goal-directed spatial navigation in the rat based on physiological data from the hippocampal formation. Neural Netw. 2003, 16, 577–584. [Google Scholar] [CrossRef]
- Pezzulo, G.; Van Der Meer, M.A.A.; Lansink, C.S.; Pennartz, C.M.A. Internally generated sequences in learning and executing goal-directed behavior. Trends Cogn. Sci. 2014, 18, 647–657. [Google Scholar] [CrossRef]
- Bingman, V.P.; Muzio, R.N. Reflections on the structural-functional evolution of the hippocampus: What is the big deal about a dentate gyrus. Brain Behav. Evol. 2017, 90, 53–61. [Google Scholar] [CrossRef]
- Herold, C.; Coppola, V.J.; Bingman, V.P. The maturation of research into the avian hippocampal formation: Recent discoveries from one of the nature’s foremost navigators. Hippocampus 2015, 25, 1193–1211. [Google Scholar] [CrossRef]
- Herold, C.; Schlömer, P.; Mafoppa-Fomat, I.; Mehlhorn, J.; Amunts, K.; Axer, M. The hippocampus of birds in a view of evolutionary connectomics. Cortex 2019, 118, 165–187. [Google Scholar] [CrossRef]
- Herold, C.; Ockermann, P.N.; Amunts, K. Behavioral training related neurotransmitter receptor expression dynamics in the nidopallium caudolaterale and the hippocampal formation of pigeons. Front. Physiol. 2022, 13, 883029. [Google Scholar] [CrossRef]
- Daw, N.D.; Gershman, S.J.; Seymour, B.; Dayan, P.; Dolan, R.J. Model-based influences on humans’ choices and striatal prediction errors. Neuron 2011, 69, 1204–1215. [Google Scholar] [CrossRef]
- Wang, F.; Schoenbaum, G.; Kahnt, T. Interactions between human orbitofrontal cortex and hippocampus support model-based inference. PLoS Biol. 2020, 18, e3000578. [Google Scholar] [CrossRef]
- Wurm, F.; Ernst, B.; Steinhauser, M. The influence of internal models on feedback-related brain activity. Cogn. Affect. Behav. Neurosci. 2020, 20, 1070–1089. [Google Scholar] [CrossRef] [PubMed]
- Halpern, B.P.; Karten, H.J.; Hodos, W. A Stereotaxic atlas of the brain of the pigeon (Columbia Livia). Am. J. Psychol. 1968, 81, 289. [Google Scholar] [CrossRef]
- Güntürkün, O.; Verhoye, M.; De Groof, G.; Van Der Linden, A. A 3-dimensional digital atlas of the ascending sensory and the descending motor systems in the pigeon brain. Brain Struct. Funct. 2013, 218, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Dayan, P. Improving generalization for temporal difference learning: The successor representation. Neural Comput. 1993, 5, 613–624. [Google Scholar] [CrossRef]
- Ekman, M.; Kusch, S.; De Lange, F.P. Successor-like representation guides the prediction of future events in human visual cortex and hippocampus. eLife 2023, 12, e78904. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Aronov, D.; Abbott, L.; Mackevicius, E.L. Neural learning rules for generating flexible predictions and computing the successor representation. eLife 2023, 12, e80680. [Google Scholar] [CrossRef] [PubMed]
- Momennejad, I.; Russek, E.M.; Cheong, J.H.; Botvinick, M.M.; Daw, N.D.; Gershman, S.J. The successor representation in human reinforcement learning. Nat. Hum. Behav. 2017, 1, 680–692. [Google Scholar] [CrossRef] [PubMed]
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Dunson, D.B.; Vehtari, A.; Rubin, D.B. Bayesian Data Analysis; Chapman and Hall/CRC: New York, NY, USA, 2013; ISBN 978-0-429-11307-9. [Google Scholar]
- Xinyu, L.; Hong, W.; Shan, L.; Yan, C.; Li, S. Adaptive common average reference for in vivo multichannel local field potentials. Biomed. Eng. Lett. 2017, 7, 7–15. [Google Scholar] [CrossRef]
- Wan, H.; Yang, H.; Xinyu, L.; Shang, Z. Research of partial least squares decoding method for motion intent. J. Biomed. Eng. 2016, 33, 626–630. [Google Scholar] [CrossRef]
- Daubechies, I. The Wavelet Transform, Time-frequency localization and signal analysis. IEEE Trans. Inf. Theory 1990, 36, 961–1005. [Google Scholar] [CrossRef]
- Kriegeskorte, N. Representational Similarity Analysis—Connecting the Branches of Systems Neuroscience. Front. Syst. Neurosci. 2008, 2, 4. [Google Scholar] [CrossRef]
- Pacheco Estefan, D.; Sánchez-Fibla, M.; Duff, A.; Principe, A.; Rocamora, R.; Zhang, H.; Axmacher, N.; Verschure, P.F.M.J. Coordinated representational reinstatement in the human hippocampus and lateral temporal cortex during episodic memory retrieval. Nat. Commun. 2019, 10, 2255. [Google Scholar] [CrossRef]
- Yaffe, R.B.; Kerr, M.S.D.; Damera, S.; Sarma, S.V.; Inati, S.K.; Zaghloul, K.A. Reinstatement of distributed cortical oscillations occurs with precise spatiotemporal dynamics during successful memory retrieval. Proc. Natl. Acad. Sci. USA 2014, 111, 18727–18732. [Google Scholar] [CrossRef] [PubMed]
- Cortes, C.; Vapnik, V. Support-vector networks. Mach. Learn. 1995, 20, 273–297. [Google Scholar] [CrossRef]
- Feher Da Silva, C.; Hare, T.A. Humans primarily use model-based inference in the two-stage task. Nat. Hum. Behav. 2020, 4, 1053–1066. [Google Scholar] [CrossRef]
- Kim, M.; Kim, J.-S.; Choi, M.-S.; Park, J.-H. Adaptive discount factor for deep reinforcement learning in continuing tasks with uncertainty. Sensors 2022, 22, 7266. [Google Scholar] [CrossRef]
- Bakkour, A.; Palombo, D.J.; Zylberberg, A.; Kang, Y.H.; Reid, A.; Verfaellie, M.; Shadlen, M.N.; Shohamy, D. The hippocampus supports deliberation during value-based decisions. eLife 2019, 8, e46080. [Google Scholar] [CrossRef]
- Ballard, I.C.; Wagner, A.D.; McClure, S.M. Hippocampal pattern separation supports reinforcement learning. Nat. Commun. 2019, 10, 1073. [Google Scholar] [CrossRef] [PubMed]
- Momennejad, I. Learning structures: Predictive representations, replay, and generalization. Curr. Opin. Behav. Sci. 2020, 32, 155–166. [Google Scholar] [CrossRef]
- Cabral, H.O.; Vinck, M.; Fouquet, C.; Pennartz, C.M.A.; Rondi-Reig, L.; Battaglia, F.P. Oscillatory dynamics and place field maps reflect hippocampal ensemble processing of sequence and place memory under nmda receptor control. Neuron 2014, 81, 402–415. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Nishida, H.; David Redish, A.; Lauwereyns, J. Theta phase shift in spike timing and modulation of gamma oscillation: A dynamic code for spatial alternation during fixation in rat hippocampal area CA1. J. Neurophysiol. 2014, 111, 1601–1614. [Google Scholar] [CrossRef]
- Jezek, K.; Henriksen, E.J.; Treves, A.; Moser, E.I.; Moser, M.-B. Theta-paced flickering between place-cell maps in the hippocampus. Nature 2011, 478, 246–249. [Google Scholar] [CrossRef]
- Levy, W.B. A Sequence Predicting CA3 is a flexible associator that learns and uses context to solve hippocampal-like tasks. Hippocampus 1996, 6, 579–590. [Google Scholar] [CrossRef]
- Shahbaba, B.; Li, L.; Agostinelli, F.; Saraf, M.; Cooper, K.W.; Haghverdian, D.; Elias, G.A.; Baldi, P.; Fortin, N.J. Hippocampal ensembles represent sequential relationships among an extended sequence of nonspatial events. Nat. Commun. 2022, 13, 787. [Google Scholar] [CrossRef]
- Mattar, M.G.; Thompson-Schill, S.L.; Bassett, D.S. The network architecture of value learning. Netw. Neurosci. 2018, 2, 128–149. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Linear Regression of the Normalized Power and | ||||||
|---|---|---|---|---|---|---|
| R2 | p-Values | F-Statistics | R2 | p-Values | F-Statistics | |
| 1–12 Hz | 0.0927 | 0.0503 | 6.0308 | 0.0537 | 0.0747 | 3.30 |
| 12–100 Hz | 0.3554 | <0.0001 | 32.23 | <0.0001 | 0.9064 | 0.01 |
| R2 | p-Values | F-Statistics | R2 | p-Values | F-Statistics | |
|---|---|---|---|---|---|---|
| 1–12 Hz | 0.2186 | 0.0002 | 16.23 | 0.0018 | 0.7469 | 0.11 |
| 12–100 Hz | 0.7551 | <0.0001 | 158.30 | 0.3042 | <0.0001 | 26.88 |
| Pigeon ID | Choice Decoding Accuracy | |
|---|---|---|
| 1–12 Hz | 12–100 Hz | |
| P090 | 0.752 ± 0.014 | 0.922 ± 0.012 |
| P093 | 0.652 ± 0.019 | 0.815 ± 0.013 |
| P014 | 0.521 ± 0.014 | 0.782 ± 0.016 |
| P025 | 0.464 ± 0.016 | 0.778 ± 0.025 |
| P021 | 0.651 ± 0.027 | 0.786 ± 0.020 |
| P029 | 0.712 ± 0.020 | 0.801 ± 0.022 |
| Average | 0.626 | 0.814 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Jin, F.; Yang, L.; Li, J.; Li, Z.; Li, M.; Shang, Z. The Hippocampus in Pigeons Contributes to the Model-Based Valuation and the Relationship between Temporal Context States. Animals 2024, 14, 431. https://doi.org/10.3390/ani14030431
Yang L, Jin F, Yang L, Li J, Li Z, Li M, Shang Z. The Hippocampus in Pigeons Contributes to the Model-Based Valuation and the Relationship between Temporal Context States. Animals. 2024; 14(3):431. https://doi.org/10.3390/ani14030431
Chicago/Turabian StyleYang, Lifang, Fuli Jin, Long Yang, Jiajia Li, Zhihui Li, Mengmeng Li, and Zhigang Shang. 2024. "The Hippocampus in Pigeons Contributes to the Model-Based Valuation and the Relationship between Temporal Context States" Animals 14, no. 3: 431. https://doi.org/10.3390/ani14030431
APA StyleYang, L., Jin, F., Yang, L., Li, J., Li, Z., Li, M., & Shang, Z. (2024). The Hippocampus in Pigeons Contributes to the Model-Based Valuation and the Relationship between Temporal Context States. Animals, 14(3), 431. https://doi.org/10.3390/ani14030431