

The Multi-Loci Genotypes of the Myostatin Gene Associated with Growth Indicators of Intensively Fattened Lambs of Latvian Sheep
 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fattening Characteristics of Intensively Fed Lambs
2.2. DNA Extraction
2.3. MSTN Gene Multi-Loci Genotypes
2.4. Statistical Analyses
3. Results
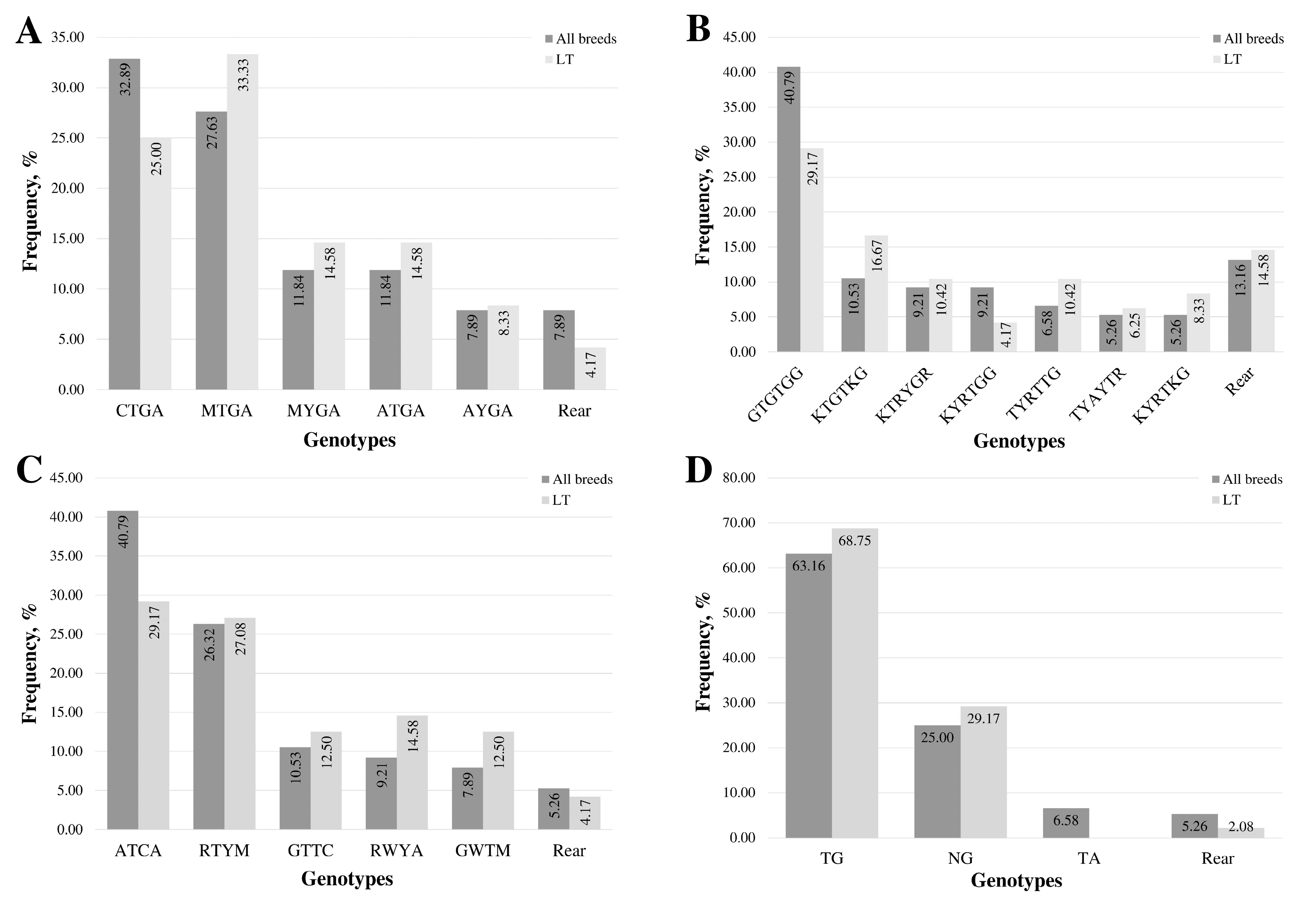
3.1. Genotypes of Promoter and Exon 1 Region of MSTN Gene
3.2. Genotypes in the Intron 1 Region of the MSTN Gene
3.3. Genotypes in the Intron 2 Region of MSTN Gene
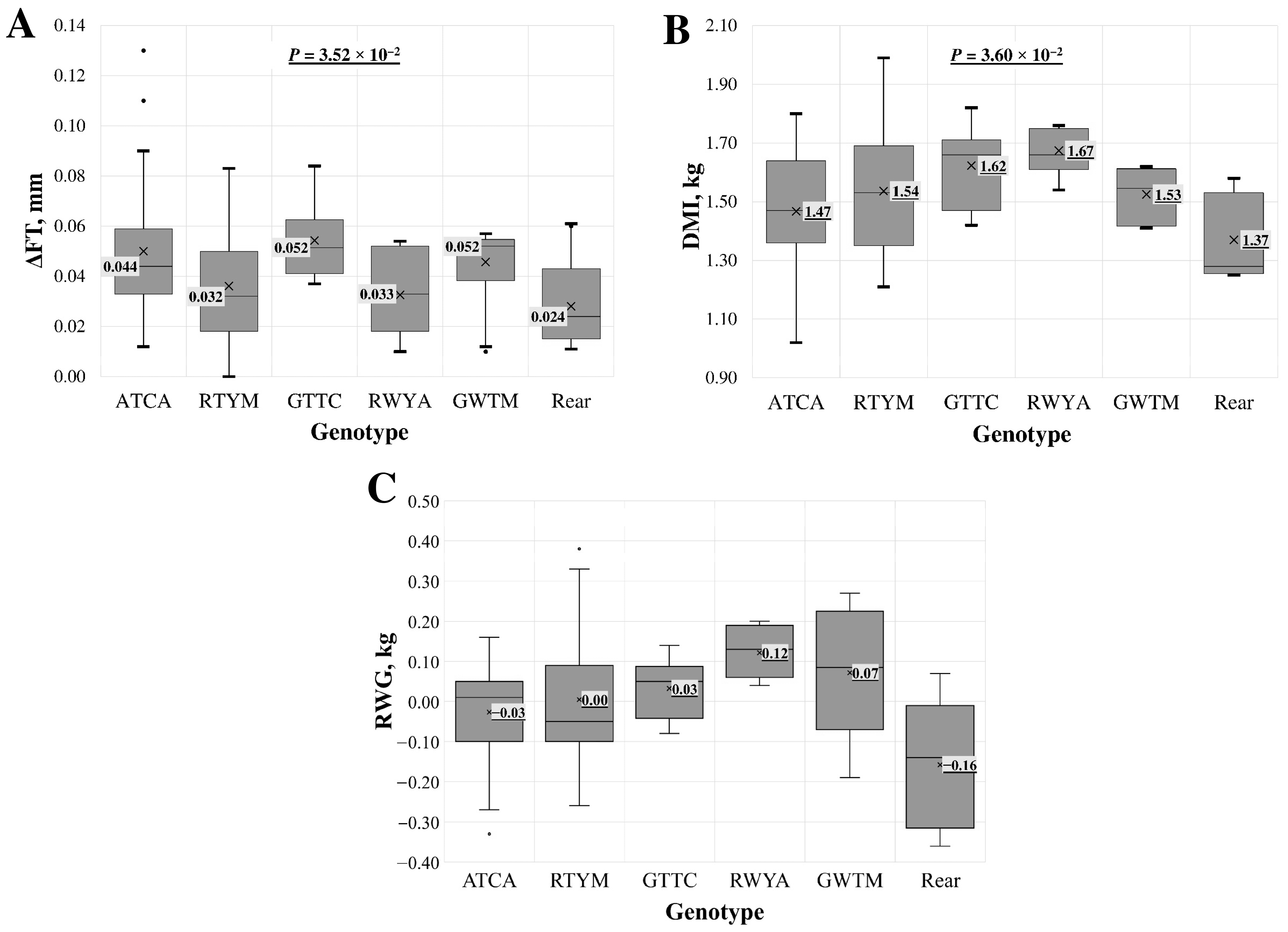
3.4. Genotypes in the 3′UTR Region of the MSTN Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sjakste, T.; Paramonova, N.; Grislis, Z.; Trapina, I.; Kairisa, D. Analysis of the Single-Nucleotide Polymorphism in the 5′UTR and Part of Intron I of the Sheep MSTN Gene. DNA Cell Biol. 2011, 30, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Osman, N.M.; Shafey, H.I.; Abdelhafez, M.A.; Sallam, A.M.; Mahrous, K.F. Genetic variations in the Myostatin gene affecting growth traits in sheep. Vet. World 2021, 14, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Wang, H.; Meng, C.; Gui, H.; Li, Y.; Chen, F.; Zhang, C.; Zhang, H.; Ding, Q.; Zhang, J.; et al. Efficient and Specific Generation of MSTN-Edited Hu Sheep Using C-CRISPR. Genes 2023, 14, 1216. [Google Scholar] [CrossRef] [PubMed]
- Aiello, D.; Patel, K.; Lasagna, E. The myostatin gene: An overview of mechanisms of action and its relevance to livestock animals. Anim. Genet. 2018, 49, 505–519. [Google Scholar] [CrossRef]
- Du, C.; Zhou, X.; Zhang, K.; Huang, S.; Wang, X.; Zhou, S.; Chen, Y. Inactivation of the MSTN gene expression changes the composition and function of the gut microbiome in sheep. BMC Microbiol. 2022, 22, 273. [Google Scholar] [CrossRef]
- Chen, M.-M.; Zhao, Y.; Xu, X.-L.; Zhang, X.-S.; Zhang, J.-L.; Wu, S.-J.; Liu, Z.-M.; Yuan, Y.-M.; Guo, X.-F.; Qi, S.-Y.; et al. A MSTNDel273C mutation with FGF5 knockout sheep by CRISPR/Cas9 promotes skeletal muscle myofiber hyperplasia via MEK-ERK-FOSL1 axis. eLife 2023, 12, RP86827. [Google Scholar] [CrossRef]
- Chacko Kaitholil, S.R.; Mooney, M.H.; Aubry, A.; Rezwan, F.; Shirali, M. Insights into the influence of diet and genetics on feed efficiency and meat production in sheep. Anim. Genet. 2024, 55, 20–46. [Google Scholar] [CrossRef]
- Clop, A.; Marcq, F.; Takeda, H.; Pirottin, D.; Tordoir, X.; Bibé, B.; Bouix, J.; Caiment, F.; Elsen, J.-M.; Eychenne, F.; et al. A mutation creating a potential illegitimate microRNA target site in the myostatin gene affects muscularity in sheep. Nat. Genet. 2006, 38, 813–818. [Google Scholar] [CrossRef]
- Hickford, J.G.H.; Forrest, R.H.; Zhou, H.; Fang, Q.; Han, J.; Frampton, C.M.; Horrell, A.L. Polymorphisms in the ovine myostatin gene (MSTN) and their association with growth and carcass traits in New Zealand Romney sheep. Anim. Genet. 2010, 41, 64–72. [Google Scholar] [CrossRef]
- Han, J.; Forrest, R.H.; Hickford, J.G.H. Genetic variations in the myostatin gene (MSTN) in New Zealand sheep breeds. Mol. Biol. Rep. 2013, 40, 6379–6384. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, H.; Hu, J.; Li, S.; Luo, Y.; Hickford, J.G.H. Two single nucleotide polymorphisms in the promoter of the ovine myostatin gene (MSTN) and their effect on growth and carcass muscle traits in New Zealand Romney sheep. J. Anim. Breed. Genet. 2016, 133, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Grochowska, E.; Borys, B.; Lisiak, D.; Mroczkowski, S. Genotypic and allelic effects of the myostatin gene (MSTN) on carcass, meat quality, and biometric traits in Colored Polish Merino sheep. Meat Sci. 2019, 151, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, M.; Grochowska, E.; Milewski, S.; Mroczkowski, S. The association between the polymorphism in the myostatin gene and growth traits in Kamieniec and Pomeranian sheep breeds. Small Rumin. Res. 2019, 177, 29–35. [Google Scholar] [CrossRef]
- Trapina, I.; Plavina, S.; Krasņevska, N.; Paramonovs, J.; Kairisa, D.; Paramonova, N. MSTN gene polymorphisms are associated with the feed efficiency of fattened lambs in Latvian sheep breeds. Agron. Res. 2024, 22, 1–15. [Google Scholar] [CrossRef]
- Zhou, H.; Hickford, J.G.H.; Fang, Q. Variation in the coding region of the myostatin (GDF8) gene in sheep. Mol. Cell. Probes 2008, 22, 67–68. [Google Scholar] [CrossRef]
- Boman, I.A.; Klemetsdal, G.; Blichfeldt, T.; Nafstad, O.; Våge, D.I. A frameshift mutation in the coding region of the myostatin gene (MSTN) affects carcass conformation and fatness in Norwegian White Sheep (Ovis aries). Anim. Genet. 2009, 40, 418–422. [Google Scholar] [CrossRef]
- Trukhachev, V.; Yatsyk, O.; Telegina, E.; Krivoruchko, A.; Zhou, H.; Hickford, J.G.H. Comparison of the myostatin (MSTN) gene in Russian Stavropol Merino sheep and New Zealand Merino sheep. Small Rumin. Res. 2018, 160, 103–106. [Google Scholar] [CrossRef]
- Sahu, A.R.; Jeichitra, V.; Rajendran, R.; Raja, A. Novel report on mutation in exon 3 of myostatin (MSTN) gene in Nilagiri sheep: An endangered breed of South India. Trop. Anim. Health Prod. 2019, 51, 1817–1822. [Google Scholar] [CrossRef]
- Farhadian, M.; Hashemi, A. Molecular Characterization and Phylogeny Based Analysis of Intron I Sequence of Myostatin (MSTN) Gene in Iranian Makuei Sheep Breed. Ann. Anim. Sci. 2016, 16, 1007–1018. [Google Scholar] [CrossRef]
- Boman, I.A.; Klemetsdal, G.; Nafstad, O.; Blichfeldt, T.; Våge, D.I. Impact of two myostatin (MSTN) mutations on weight gain and lamb carcass classification in Norwegian White Sheep (Ovis aries). Genet. Sel. Evol. 2010, 42, 4. [Google Scholar] [CrossRef]
- Masri, A.Y.; Lambe, N.R.; Macfarlane, J.M.; Brotherstone, S.; Haresign, W.; Bünger, L. Evaluating the effects of a single copy of a mutation in the myostatin gene (c.*1232G>A) on carcass traits in crossbred lambs. Meat Sci. 2011, 87, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Hadjipavlou, G.; Matika, O.; Clop, A.; Bishop, S.C. Two single nucleotide polymorphisms in the myostatin (GDF8) gene have significant association with muscle depth of commercial Charollais sheep. Anim. Genet. 2008, 39, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Hope, M.; Haynes, F.; Oddy, H.; Koohmaraie, M.; Al-Owaimer, A.; Geesink, G. The effects of the myostatin g+6723G>A mutation on carcass and meat quality of lamb. Meat Sci. 2013, 95, 118–122. [Google Scholar] [CrossRef] [PubMed]
- LAAA. Genealogy Programs. 2024. Available online: https://www.laaa.lv/lv/skirnes-saimniecibas/ciltsdarba-programmas/ (accessed on 5 August 2024).
- Eurostat. 2024. Available online: https://ec.europa.eu/eurostat/databrowser/view/APRO_MT_LSSHEEP/default/table?lang=en (accessed on 5 August 2024).
- Eurostat. 2024. Available online: https://ec.europa.eu/eurostat/databrowser/view/tag00045/default/table?lang=en (accessed on 5 August 2024).
- Berry, D.P.; Crowley, J.J. Residual intake and body weight gain: A new measure of efficiency in growing cattle. J. Anim. Sci. 2012, 90, 109–115. [Google Scholar] [CrossRef]
- Lima, N.L.L.; de Ribeiro, C.R.F.; de Sá, H.C.M.; Leopoldino Júnior, I.; Cavalcanti, L.F.L.; Santana, R.A.V.; Furusho-Garcia, I.F.; Pereira, I.G. Economic analysis, performance, and feed efficiency in feedlot lambs. Rev. Bras. de Zootec. 2017, 46, 821–829. [Google Scholar] [CrossRef]
- Hervás, G.; Toral, P.G.; Fernández-Díez, C.; Badia, A.D.; Frutos, P. Effect of Dietary Supplementation with Lipids of Different Unsaturation Degree on Feed Efficiency and Milk Fatty Acid Profile in Dairy Sheep. Animals 2021, 11, 2476. [Google Scholar] [CrossRef]
- Johnson, P.L.; Hickey, S.; Knowler, K.; Wing, J.; Bryson, B.; Hall, M.; Jonker, A.; Janssen, P.H.; Dodds, K.G.; McEwan, J.C.; et al. Genetic parameters for residual feed intake, methane emissions, and body composition in New Zealand maternal sheep. Front. Genet. 2022, 13, 911639. [Google Scholar] [CrossRef]
- Trapina, I.; Kairisa, D.; Paramonova, N. Analysing the Cost of Concentrated Feed and Income from Meat in Relation to Relative Growth Rate and Kleiber’s Ratio in Intensive Fattening of Latvian Dark-Headed Lambs. Agriculture 2024, 14, 593. [Google Scholar] [CrossRef]
- Wakchaure, R.; Ganguly, S.; Praveen, P.K.; Kumar, A.; Sharma, S.; Mahajan, T. Marker Assisted Selection (MAS) in Animal Breeding: A Review. J. Drug Metab. Toxicol. 2015, 6, e127. [Google Scholar] [CrossRef]
- Bārzdiņa, D.; Kairiša, D. Analysis of scrapie resistance and productivity of Latvian dark head breed old type sheep compliant to the conservation program of the genetic resources. Agron. Res. 2023, 21, 1058–1063. [Google Scholar] [CrossRef]
- Trapina, I.; Plavina, S.; Krasņevska, N.; Paramonovs, J.; Kairisa, D.; Paramonova, N. IGF1 and IGF2 gene polymorphisms are associated with the feed efficiency of fattened lambs in Latvian sheep breads. Agron. Res. 2024, 22, 1–15. [Google Scholar] [CrossRef]
- Trapina, I.; Kairisa, D.; Paramonova, N. Feed efficiency indicators and hormones related to nutrient metabolism in intensive fattened lambs of sire rams of different sheep breeds in Latvia. Agron. Res. 2023, 21 (Suppl. S2), 598–610. [Google Scholar] [CrossRef]
- LAAA. Instrukcija “Snieguma pārbaudei un ģenētiskās kvalitātes noteikšanai gaļas tipa šķirņu aitām” [Instruction “Performance Testing and Determination of Genetic Quality in Sheep of Meat-Type Breeds”] in Latvian. 2024. Available online: https://www.laaa.lv/data/uploads/ciltsdarba-programmas/2024/Instrukcija_15_04_2024_gala.doc (accessed on 5 August 2024).
- Šenfelde, L.; Kairiša, D.; Bārzdiņa, D. Effect of concentrate feeding technology on nutrient digestibility in Latvian Dark-Head lambs. Agron. Res. 2020, 18 (Suppl. S1), 1000–1009. [Google Scholar] [CrossRef]
- Johansen, J.; Aastveit, A.H.; Egelandsdal, B.; Kvaal, K.; Røe, M. Validation of the EUROP system for lamb classification in Norway; repeatability and accuracy of visual assessment and prediction of lamb carcass composition. Meat Sci. 2006, 74, 497–509. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Datasets. Mol. Biol. Evol 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows, Version 25.0; Released 2017; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- Bārzdiņa, D.; Kairiša, D. The production and quality analysis of Latvian darkhead breed lambs. In Proceedings of the Nordic View to Sustainable Rural Revelopment, Proceedings of the 25th NIF Congress, Riga, Latvia, 16–18 June 2015; pp. 361–366. [Google Scholar]
- Boareki, M.; Schenkel, F.; Kennedy, D.; Cánovas, A. Prediction of Genetic Resistance for Scrapie in Ungenotyped Sheep Using a Linear Animal Model. Genes 2021, 12, 1432. [Google Scholar] [CrossRef]
- Kijas, J.W.; McCulloch, R.; Edwards, J.E.H.; Oddy, V.H.; Lee, S.H.; van der Werf, J. Evidence for multiple alleles affecting muscling and fatness at the Ovine GDF8 locus. BMC Genet. 2007, 8, 80. [Google Scholar] [CrossRef]
- Tellam, R.; Cockett, N.; Vuocolo, T.; Bidwell, C. Genes Contributing to Genetic Variation of Muscling in Sheep. Front. Genet. 2012, 3, 164. [Google Scholar] [CrossRef]
- Bora, S. Applications of Genomic Selection in Animal Breeding; Challenges and Opportunities. Open Access J. Microbiol. Biotechnol. 2023, 48, 107–117. [Google Scholar] [CrossRef]
- Carracelas, B.; Navajas, E.A.; Vera, B.; Ciappesoni, G. SNP arrays evaluation as tools in genetic improvement in Corriedale sheep in Uruguay. Agrociencia Urug. 2022, 26, e998. [Google Scholar] [CrossRef]
- Kaseja, K.; Mucha, S.; Smith, E.; Yates, J.; Banos, G.; Conington, J. Including genotypic information in genetic evaluations increases the accuracy of sheep breeding values. J. Anim. Breed. Genet. 2023, 140, 462–471. [Google Scholar] [CrossRef]
- Nel, C.L.; van der Werf, J.H.J.; Rauw, W.M.; Cloete, S.W.P. Challenges and strategies for genetic selection of sheep better adapted to harsh environments. Anim. Front. Rev. Mag. Anim. Agric. 2023, 13, 43. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Parameter | Genotype of 3′UTR (c.*707DelT/c.*1232G>A) | p | ||||
|---|---|---|---|---|---|---|
| T/G | N/G | T/A | Rear * | |||
| Birth weight, kg | Mean ± SD | 4.34 ± 0.81 | 4.32 ± 0.56 | 5.76 ± 0.57 | 4.85 ± 0.13 | 4.76 × 10−4 |
| Median (IQR) | 4.30 (1.15) | 4.20 (1.00) | 5.60 (1.00) | 4.85 (0.25) | - | |
| Average daily gain, g | Mean ± SD | 333.35 ± 66.88 | 349.86 ± 66.88 | 261.30 ± 34.68 | 286.97 ± 96.57 | - |
| Median (IQR) | 321.74 (109.44) | 352.00 (82.61) | 272.73 (75.32) | 321.33 (174.28) | 3.72 × 10−2 | |
| Dry matter intake, kg | Mean ± SD | 1.54 ± 0.19 | 1.55 ± 0.16 | 1.27 ± 0.18 | 1.37 ± 0.28 | 2.35 × 10−3 |
| Median (IQR) | 1.53 (0.27) | 1.53 (0.26) | 1.24 (0.34) | 1.42 (0.53) | - | |
| Slaughter yield, % | Mean ± SD | 44.64 ± 2.30 | 44.92 ± 2.66 | 48.76 ± 2.08 | 46.55 ± 2.64 | 2.29 × 10−3 |
| Median (IQR) | 44.15 (3.38) | 44.90 (4.40) | 49.4 (3.40) | 46.05 (4.80) | - | |
| Muscle development | Mean ± SD | 2.84 ± 0.42 | 2.89 ± 0.27 | 1.90 ± 0.55 | 2.38 ± 0.75 | - |
| Median (IQR) | 3.00 (0.00) | 3.00 (0.00) | 2.00 (0.75) | 2.50 (1.38) | 1.00 × 10−4 | |
| Fat degree | Mean ± SD | 2.95 ± 0.38 | 2.97 ± 0.61 | 2.40 ± 0.22 | 2.50 ± 0.58 | - |
| Median (IQR) | 3.00 (0.00) | 3.00 (0.50) | 2.50 (0.25) | 2.50 (1.00) | 3.70 × 10−3 | |
| RGR | Mean ± SD | 0.35 ± 0.10 | 0.37 ± 0.08 | 0.24 ± 0.02 | 0.26 ± 0.09 | 2.02 × 10−2 |
| Median (IQR) | 0.33 (0.14) | 0.38 (0.13) | 0.25 (0.04) | 0.28 (0.16) | - | |
| KR | Mean ± SD | 18.50 ± 2.88 | 18.83 ± 2.58 | 15.02 ± 1.26 | 15.79 ± 4.78 | - |
| Median (IQR) | 18.67 (3.75) | 19.13 (3.94) | 15.72 (2.39) | 17.05 (8.54) | 2.03 × 10−2 | |
| RWG | Mean ± SD | 0.02 ± 0.14 | −0.01 ± 0.12 | −0.14 ± 0.20 | −0.06 ± 0.17 | 9.94 × 10−3 |
| Median (IQR) | 0.04 (0.18) | −0.03 (0.17) | −0.26 (0.37) | −0.03 (0.31) | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trapina, I.; Kairisa, D.; Plavina, S.; Krasnevska, N.; Paramonovs, J.; Senfelde, L.; Paramonova, N. The Multi-Loci Genotypes of the Myostatin Gene Associated with Growth Indicators of Intensively Fattened Lambs of Latvian Sheep. Animals 2024, 14, 3143. https://doi.org/10.3390/ani14213143
Trapina I, Kairisa D, Plavina S, Krasnevska N, Paramonovs J, Senfelde L, Paramonova N. The Multi-Loci Genotypes of the Myostatin Gene Associated with Growth Indicators of Intensively Fattened Lambs of Latvian Sheep. Animals. 2024; 14(21):3143. https://doi.org/10.3390/ani14213143
Chicago/Turabian StyleTrapina, Ilva, Daina Kairisa, Samanta Plavina, Nikole Krasnevska, Jegors Paramonovs, Liga Senfelde, and Natalia Paramonova. 2024. "The Multi-Loci Genotypes of the Myostatin Gene Associated with Growth Indicators of Intensively Fattened Lambs of Latvian Sheep" Animals 14, no. 21: 3143. https://doi.org/10.3390/ani14213143
APA StyleTrapina, I., Kairisa, D., Plavina, S., Krasnevska, N., Paramonovs, J., Senfelde, L., & Paramonova, N. (2024). The Multi-Loci Genotypes of the Myostatin Gene Associated with Growth Indicators of Intensively Fattened Lambs of Latvian Sheep. Animals, 14(21), 3143. https://doi.org/10.3390/ani14213143