Love Hug—Functional Validation of Nuptial Pad-Secreted Pheromone in Anurans

 , ,
, ,  ,
,  ,
,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Transcriptome Sequencing and Analysis
2.3. Protein Sequence Alignment and Amino Acid Characteristic Analysis
2.4. Recombinant Expression of Pheromone Protein
2.5. Behavioral Experiments and Data Analysis
3. Results
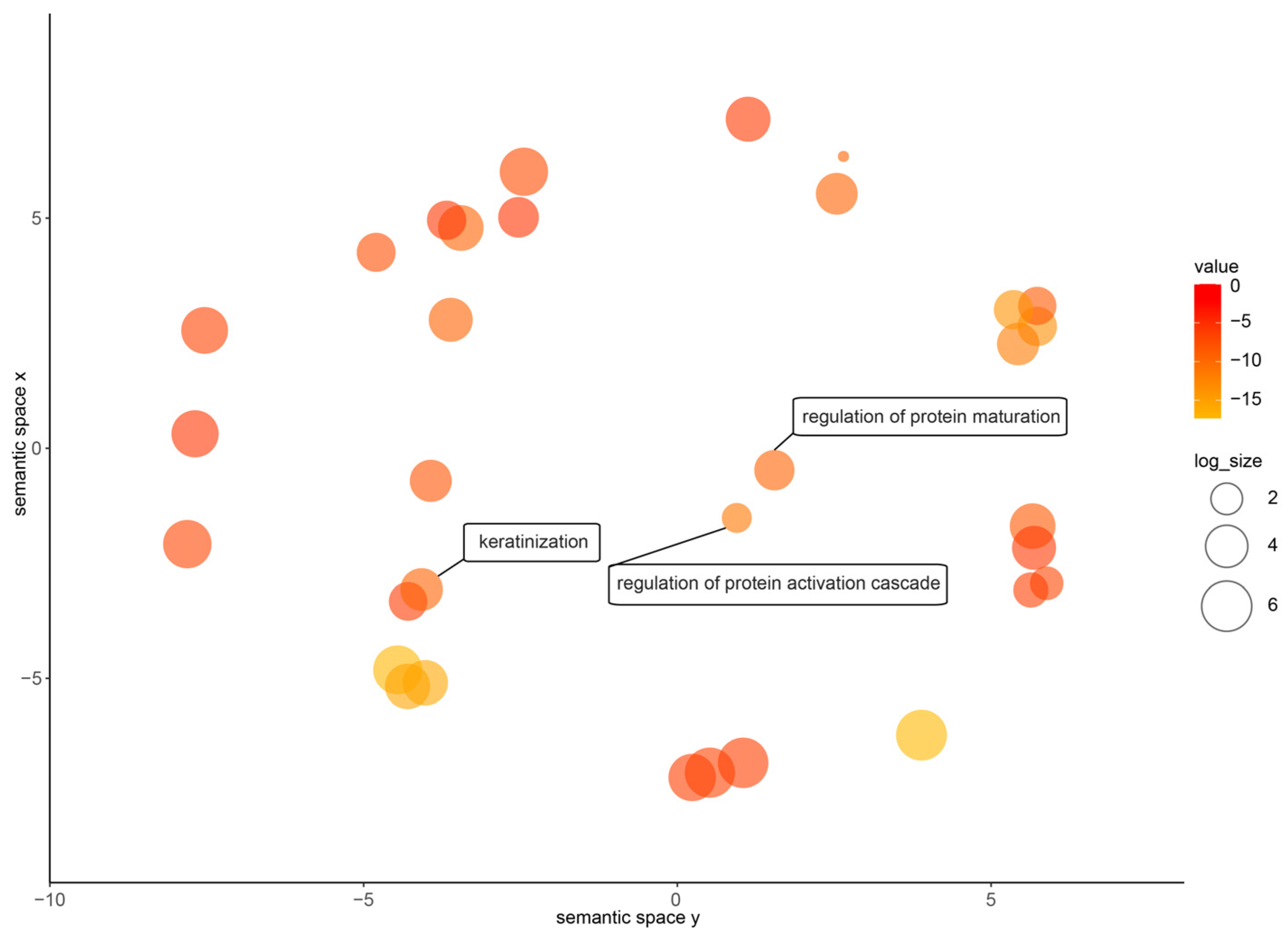
3.1. PMF Sequence Acquisition
3.2. Amino acid Sequence Characteristics of N. pleuraden PMF
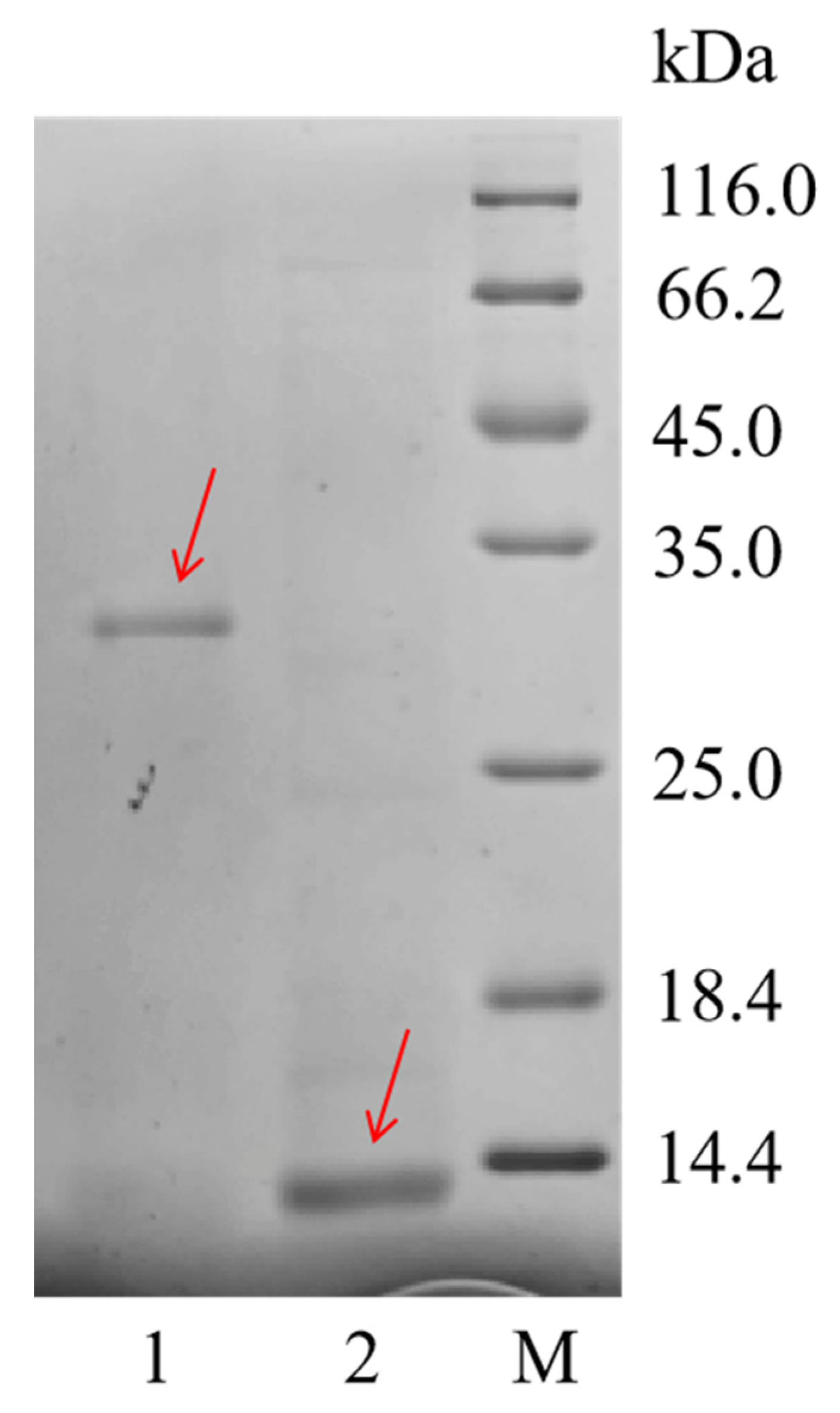
3.3. Recombinant Expression of N. pleuraden Amplexin
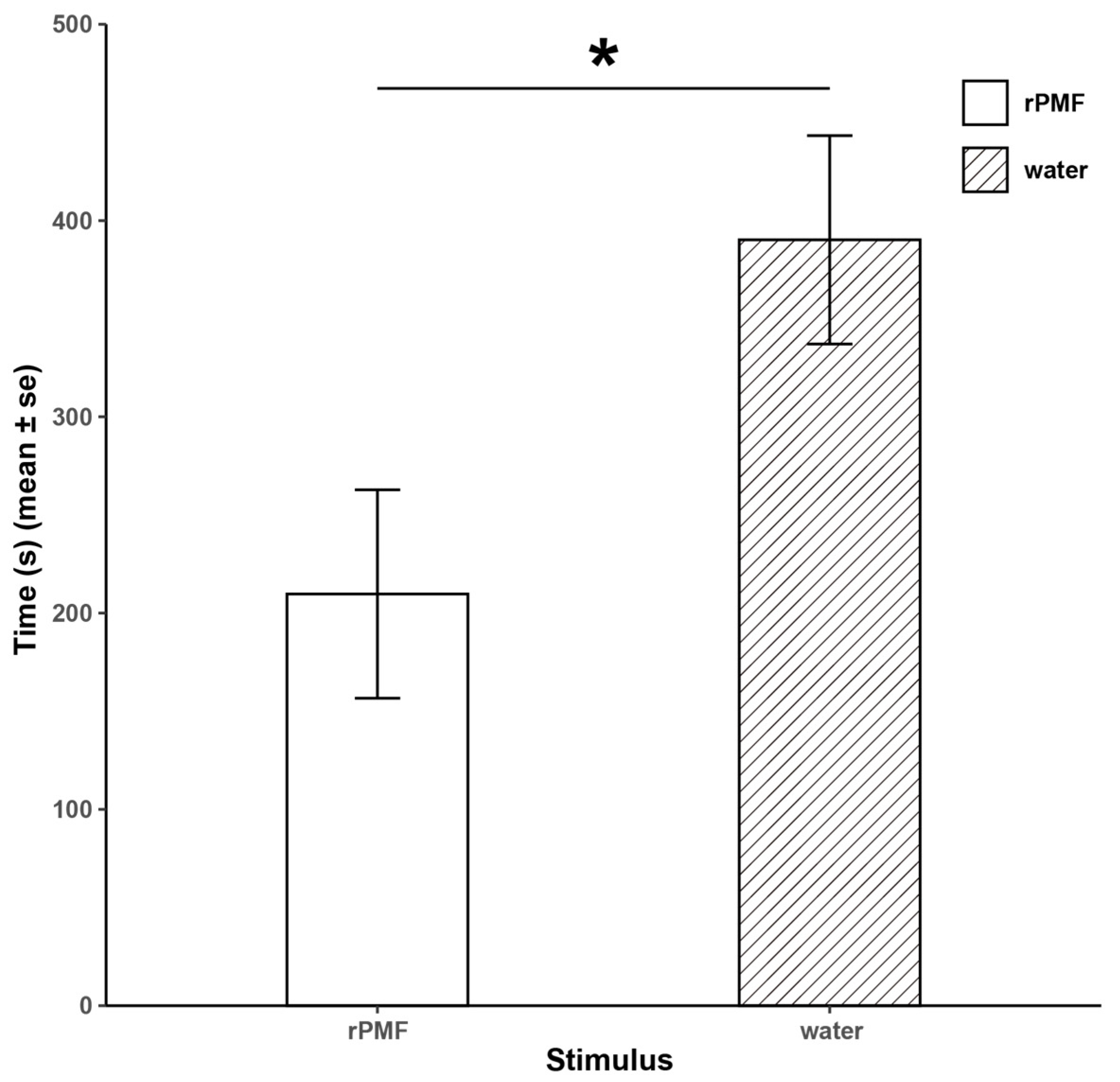
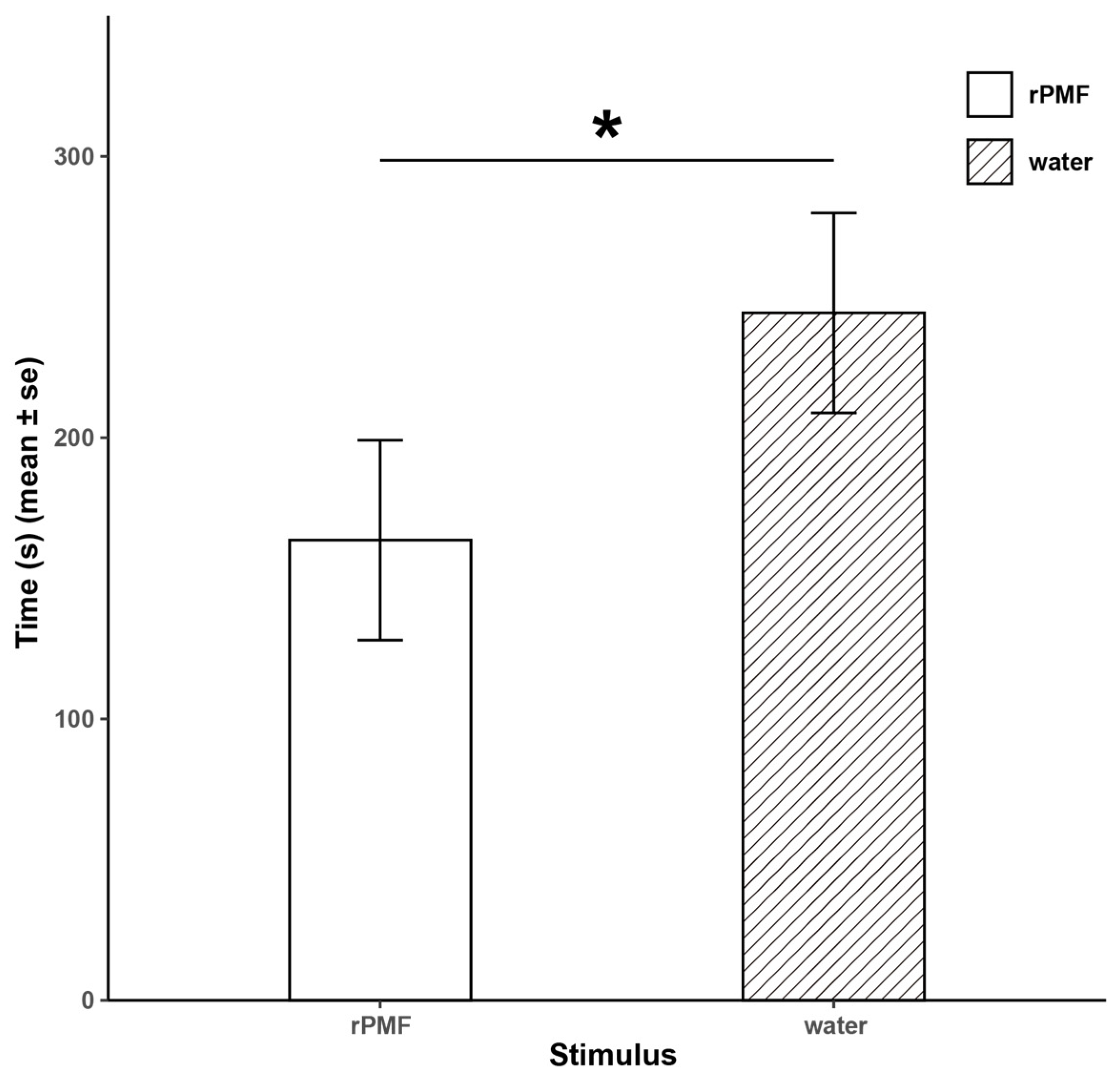
3.4. Functional Verification of N. pleuraden Amplexin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carvajal-Castro, J.D.; López-Aguirre, Y.; Ospina-L, A.M.; Santos, J.C.; Rojas, B.; Vargas-Salinas, F. Much more than a clasp: Evolutionary patterns of amplexus diversity in anurans. Biol. J. Linn. Soc. 2020, 129, 652–663. [Google Scholar] [CrossRef]
- Wells, K.D. The Ecology and Behavior of Amphibians; University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Duellman, W.E.; Trueb, L. Biology of Amphibians; McGraw-Hill: New York, NY, USA, 1986; p. 670. [Google Scholar]
- Kelehear, C.; Shine, R. Non-reproductive male cane toads (Rhinella marina) withhold sex-identifying information from their rivals. Biol. Lett. 2019, 15, 20190462. [Google Scholar] [CrossRef] [PubMed]
- Luna, M.C.; Mcdiarmid, R.W.; Faivovich, J. From erotic excrescences to pheromone shots: Structure and diversity of nuptial pads in anurans. Biol. J. Linn. Soc. 2018, 124, 403–446. [Google Scholar] [CrossRef]
- Willaert, B.; Bossuyt, F.; Janssenswillen, S.; Adriaens, D.; Baggerman, G.; Matthijs, S.; Pauwels, E.; Proost, P.; Raepsaet, A.; Schoofs, L.; et al. Frog nuptial pads secrete mating season-specific proteins related to salamander pheromones. J. Exp. Biol. 2013, 216, 4139–4143. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.O.; Tsang, L.; Licht, P. Comparative histochemistry of the sexually dimorphic skin glands of anuran amphibians. Copeia 1993, 1993, 133–143. [Google Scholar] [CrossRef]
- Houck, L.D.; Reagan, N.L. Male courtship pheromones increase female receptivity in a plethodontid salamander. Anim. Behav. 1990, 39, 729–734. [Google Scholar] [CrossRef]
- Kikuyama, S.; Toyoda, F.; Ohmiya, Y.; Matsuda, K.; Tanaka, S.; Hayashi, H. Sodefrin: A female-attracting peptide pheromone in newt cloacal glands. Science 1995, 267, 1643–1645. [Google Scholar] [CrossRef] [PubMed]
- Rollmann, S.M.; Houck, L.D.; Feldhoff, R.C. Proteinaceous pheromone affects female receptivity in a terrestrial salamander. Science 1999, 285, 1907–1909. [Google Scholar] [CrossRef] [PubMed]
- Maex, M.; Treer, D.; De Greve, H.; Proost, P.; Van Bocxlaer, I.; Bossuyt, F. Exaptation as a mechanism for functional reinforcement of an animal pheromone system. Curr. Biol. 2018, 28, 1–6. [Google Scholar] [CrossRef]
- Nakada, T.; Toyoda, F.; Matsuda, K.; Nakakura, T.; Hasunuma, I.; Yamamoto, K.; Onoue, S.; Yokosuka, M.; Kikuyama, S. Imorin: A sexual attractiveness pheromone in female red-bellied newts (Cynops pyrrhogaster). Sci. Rep. 2017, 7, 41334. [Google Scholar] [CrossRef]
- Yamamoto, K.; Kawai, Y.; Hayashi, T.; Ohe, Y.; Hayashi, H.; Toyoda, F.; Kawahara, G.; Iwata, T.; Kikuyama, S. Silefrin, a sodefrin-like pheromone in the abdominal gland of the sword-tailed newt, Cynops ensicauda. FEBS Lett. 2000, 472, 267–270. [Google Scholar] [CrossRef]
- Houck, L.D. Pheromone communication in amphibians and reptiles. Annu. Rev. Physiol. 2009, 71, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Wells, K.D. The social behaviour of anuran amphibians. Anim. Behav. 1977, 25, 666–693. [Google Scholar] [CrossRef]
- Bossuyt, F.; Schulte, L.M.; Maex, M.; Janssenswillen, S.; Novikova, P.Y.; Biju, S.D.; Peer, Y.V.D.; Matthijs, S.; Roelants, K.; Martel, A.; et al. Multiple independent recruitment of sodefrin precursor-like factors in Anuran sexually dimorphic glands. Mol. Biol. Evol. 2019, 36, 1921–1930. [Google Scholar] [CrossRef] [PubMed]
- Pearl, C.A.; Cervantes, M.; Chan, M.; Ho, U.; Shoji, R.; Thomas, E.O. Evidence for a mate-attracting chemosignal in the dwarf African clawed frog Hymenochirus. Horm. Behav. 2000, 38, 67–74. [Google Scholar] [CrossRef]
- Schulte, L.M.; Martel, A.; Cruz-Elizalde, R.; Ramírez-Bautista, A.; Bossuyt, F. Love bites: Male frogs (Plectrohyla, Hylidae) use teeth scratching to deliver sodefrin precursor-like factors to females during amplexus. Front. Zool. 2021, 18, 59. [Google Scholar] [CrossRef]
- Waldman, B.; Bishop, P.J. Chemical communication in an archaic anuran amphibian. Behav. Ecol. 2004, 15, 88–93. [Google Scholar] [CrossRef]
- Houck, L.D.; Palmer, C.A.; Watts, R.A.; Arnold, S.J.; Feldhoff, P.W.; Feldhoff, R.C. A new vertebrate courtship pheromone, PMF, affects female receptivity in a terrestrial salamander. Anim. Behav. 2007, 73, 315–320. [Google Scholar] [CrossRef]
- Janssenswillen, S.; Vandebergh, W.; Treer, D.; Willaert, B.; Maex, M.; Van Bocxlaer, I.; Bossuyt, F. Origin and diversification of a salamander sex pheromone system. Mol. Biol. Evol. 2015, 32, 472–480. [Google Scholar] [CrossRef]
- Love, M.; Anders, S.; Huber, W. Differential analysis of count data–the deseq2 package. Genome Biol. 2014, 15, 10–1186. [Google Scholar] [CrossRef]
- Bryant, D.M.; Johnson, K.; DiTommaso, T.; Tickle, T.; Couger, M.B.; Payzin-Dogru, D.; Lee, T.J.; Leigh, N.D.; Kuo, T.H.; Davis, F.G.; et al. A Tissue-Mapped Axolotl de Novo Transcriptome Enables Identification of Limb Regeneration Factors. Cell Rep. 2017, 18, 762–776. [Google Scholar] [CrossRef] [PubMed]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- KB, N. GeneDoc: Analysis and visualization of genetic variation. EMBnet News 1997, 4, 1–4. [Google Scholar] [CrossRef]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Houck, L.D.; Watts, R.A.; Arnold, S.J.; Bowen, K.E.; Kiemnec, K.M.; Godwin, H.A.; Feldhoff, P.W.; Feldhoff, R.C. A recombinant courtship pheromone affects sexual receptivity in a plethodontid salamander. Chem. Senses 2008, 33, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, L. ggplot2: Elegant Graphics for Data Analysis by WICKHAM, H. Biometrics 2011, 67, 678–679. [Google Scholar] [CrossRef]
- Forbes, M.S.; Dent, J.N.; Singhas, C.A. The developmental cytology of the nuptial pad in the red-spotted newt. Dev. Biol. 1975, 46, 56–78. [Google Scholar] [CrossRef]
- Parakkal, P.E.; Ellis, R.A. A cytochemical and electron microscopic study of the thumb pad in Rana pipiens. Exp. Cell Res. 1963, 32, 280–288. [Google Scholar] [CrossRef]
- Wilburn, D.B.; Bowen, K.E.; Gregg, R.G.; Cai, J.; Feldhoff, P.W.; Houck, L.D.; Feldhoff, R.C. Proteomic and utr analyses of a rapidly evolving hypervariable family of vertebrate pheromones. Evolution 2012, 66, 2227–2239. [Google Scholar] [CrossRef]
- Palmer, C.A.; Watts, R.A.; Hastings, A.P.; Houck, L.D.; Arnold, S.J. Rapid evolution of plethodontid modulating factor, a hypervariable salamander courtship pheromone, is driven by positive selection. J. Mol. Evol. 2010, 70, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Wilburn, D.B.; Bowen, K.E.; Doty, S.; Arumugam, S.; Lane, A.N.; Feldhoff, P.W.; Feldhoff, R.C. Structural insights into the evolution of a sexy protein: Novel topology and restricted backbone flexibility in a hypervariable vertebrate pheromone. PLoS ONE 2014, 9, e96975. [Google Scholar] [CrossRef] [PubMed]
- Galat, A.; Gross, G.; Drevet, P.; Ménez, A. Conserved structural determinants in three-fingered protein domains. FEBS J. 2008, 275, 3207–3225. [Google Scholar] [CrossRef] [PubMed]
- Doty, K.A.; Wilburn, D.B.; Bowen, K.E.; Feldhoff, P.W.; Feldhoff, R.C. Co-option and evolution of non-olfactory proteinaceous pheromones in a terrestrial lungless salamander. J. Proteom. 2016, 135, 101–111. [Google Scholar]
- Wilburn, D.B.; Bowen, K.E.; Feldhoff, P.W.; Feldhoff, R.C. Proteomic analyses of courtship pheromones in the redback salamander, Plethodon cinereus. J. Chem. Ecol. 2014, 40, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.A.; Watts, R.A.; Houck, L.D.; Picard, A.L.; Arnold, S.J. Evolutionary replacement of components in a salamander pheromone signaling complex: More evidence for phenotypic-molecular decoupling. Evolution 2007, 61, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Kiemnec-Tyburczy, K.M.; Watts, R.A.; Gregg, R.G.; von Borstel, D.; Arnold, S.J. Evolutionary shifts in courtship pheromone composition revealed by EST analysis of plethodontid salamander mental glands. Gene 2009, 432, 75–81. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A resource for timelines, timetrees, and divergence times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef]
- Zhang, P.; Zhou, H.; Chen, Y.Q.; Liu, Y.F.; Qu, L.H. Mitogenomic perspectives on the origin and phylogeny of living amphibians. Syst. Biol. 2005, 54, 391–400. [Google Scholar] [CrossRef]
- Wilburn, D.B.; Eddy, S.L.; Chouinard, A.J.; Arnold, S.J.; Feldhoff, R.C.; Houck, L.D. Pheromone isoform composition differentially affects female behaviour in the red-legged salamander, Plethodon shermani. Anim. Behav. 2015, 100, 1–7. [Google Scholar] [CrossRef]
- Wilburn, D.B.; Arnold, S.J.; Houck, L.D.; Feldhoff, P.W.; Feldhoff, R.C. Gene duplication, co-option, structural evolution, and phenotypic tango in the courtship pheromones of plethodontid salamanders. Herpetologica 2017, 73, 206–219. [Google Scholar] [CrossRef]
- Bossuyt, F.; Maex, M.; Treer, D.; Schulte, L.M.; Van Bocxlaer, I.; Janssenswillen, S. Chemistry between salamanders: Evolution of the SPF courtship pheromone system in Salamandridae. In Chemical Signals in Vertebrates 14; Buesching, C.D., Ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2019; pp. 205–220. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcript ID | Translated Protein Name | Expression Level (Average FPKM) | Accession Number |
|---|---|---|---|
| Np_f2p60_635_transcript36823_g2 | Npamplexin01 | 18.0 | PP583591 |
| Np_f6p60_691_transcript36243_g1 | Npamplexin02 | 110.0 | PP583592 |
| Np_f5p60_577_transcript37403_g1 | Npamplexin03 | 16.0 | PP583593 |
| Np_f2p60_624_transcript36920_g2 | Npamplexin04 | 50.3 | PP583594 |
| Np_f7p60_783_transcript35402_g2 | Npamplexin05 | 6093.0 | PP583595 |
| Np_f3p60_601_transcript37151_g1 | Npamplexin06 | 79.7 | PP583596 |
| Np_f2p33_731_transcript35890_g1 | Npamplexin07 | 33.7 | PP583597 |
| Np_f2p15_911_transcript34472_g1 | Npamplexin08 | 410.7 | PP623143 |
| Np_f2p53_997_transcript33771_g1 | Npamplexin09 | 126.3 | PP623144 |
| Np_f2p60_533_transcript37873_g1 | Npamplexin10 | 583.3 | PP623145 |
| Np_f4p60_809_transcript35147_g1 | Npamplexin11 | 620.3 | PP623146 |
| Np_f7p60_745_transcript35648_g1 | Npamplexin12 | 620.3 | PP623147 |
| Np_f9p60_755_transcript35756_g1 | Npamplexin13 | 1921.3 | PP623148 |
| Np_f11p60_818_transcript35677_g1 | Npamplexin14 | 2115.0 | PP623149 |
| Np_f2p16_866_transcript34815_g1 | Npamplexin15 | 1658.0 | PP623150 |
| Np_f2p60_865_transcript34782_g1 | Npamplexin16 | 45.7 | PP623151 |
| Np_f3p60_896_transcript34533_g2 | Npamplexin17 | 1996.0 | PP623152 |
| Np_f4p60_854_transcript34833_g1 | Npamplexin18 | 9.0 | PP623153 |
| Sequences | Npamplexin02 | Npamplexin05 | Npamplexin08 | Npamplexin14 | Npamplexin15 | Npamplexin17 | Npamplexin18 | amplexin1 | amplexin2 | amplexin3 | amplexin4 | amplexin5 | P. shermani | A. ferreus |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Npamplexin02 | ||||||||||||||
| Npamplexin05 | 47.8 | |||||||||||||
| Npamplexin08 | 43.4 | 47.4 | ||||||||||||
| Npamplexin14 | 22.7 | 22.5 | 21.1 | |||||||||||
| Npamplexin15 | 20.8 | 21.3 | 22.0 | 62.9 | ||||||||||
| Npamplexin17 | 20.8 | 21.3 | 22.0 | 61.2 | 98.3 | |||||||||
| Npamplexin18 | 17.2 | 13.9 | 15.2 | 46.4 | 42.4 | 42.4 | ||||||||
| amplexin1 | 28.3 | 30.1 | 31.2 | 26.4 | 23.7 | 24.4 | 19.2 | |||||||
| amplexin2 | 27.5 | 30.1 | 30.4 | 26.4 | 23.7 | 24.4 | 18.5 | 90.3 | ||||||
| amplexin3 | 28.3 | 30.9 | 31.2 | 27.1 | 24.4 | 25.1 | 19.2 | 87.5 | 97.1 | |||||
| amplexin4 | 23.3 | 25.3 | 26.4 | 21.4 | 19.4 | 20.1 | 17.8 | 83.6 | 83.6 | 81.7 | ||||
| amplexin5 | 23.3 | 26.1 | 26.4 | 20.7 | 18.7 | 19.4 | 17.8 | 78.8 | 86.5 | 84.6 | 94.7 | |||
| Plethodon shermani | 16.8 | 19.3 | 20.1 | 18.1 | 15.8 | 15.8 | 14.1 | 20.3 | 21.2 | 20.3 | 17.5 | 18.5 | ||
| Aneides ferreus | 17.6 | 17.6 | 17.6 | 18.1 | 16.6 | 15.8 | 15.7 | 22.2 | 23.1 | 23.1 | 20.3 | 20.3 | 38.8 |
| Transcript ID | Number of Amino Acids | Molecular Weight | Theoretical pI | Estimated Half-Life (Mammalian Reticulocytes, In Vitro) | Instability Index | Grand Average of Hydropathicity |
|---|---|---|---|---|---|---|
| Npamplexin01 | 71 | 8478.61 | 7.66 | 5.5 h | 24.74 | −0.682 |
| Npamplexin02 | 90 | 10,377.82 | 8.15 | 5.5 h | 45.27 | −0.891 |
| Npamplexin03 | 70 | 8139.16 | 5.73 | 5.5 h | 19.74 | −1.01 |
| Npamplexin04 | 96 | 11,008.44 | 4.46 | 5.5 h | 38.8 | −0.365 |
| Npamplexin05 | 96 | 11,008.44 | 4.46 | 5.5 h | 38.8 | −0.365 |
| Npamplexin06 | 108 | 12,360.1 | 4.38 | 5.5 h | 43.35 | −0.121 |
| Npamplexin07 | 106 | 11,824.42 | 4.98 | 5.5 h | 42.47 | −0.347 |
| Npamplexin08 | 96 | 11,016.67 | 7.65 | 5.5 h | 42.85 | −0.489 |
| Npamplexin09 | 96 | 1097.55 | 6.74 | 5.5 h | 44.56 | −0.531 |
| Npamplexin10 | 82 | 9268.46 | 4.59 | 5.5 h | 47.73 | −0.722 |
| Npamplexin11 | 117 | 12,912.49 | 4.56 | 5.5 h | 52.50 | −0.393 |
| Npamplexin12 | 72 | 8451.72 | 8.15 | 5.5 h | 39.52 | −0.578 |
| Npamplexin13 | 110 | 12,299.89 | 4.74 | 0.8 h | 63.05 | −0.610 |
| Npamplexin14 | 105 | 11,926.81 | 8.21 | 5.5 h | 52,26 | −0.549 |
| Npamplexin15 | 104 | 11,887.83 | 8.56 | 1 h | 54.75 | −0.560 |
| Npamplexin16 | 104 | 11,887.83 | 8.56 | 1 h | 54.75 | −0.560 |
| Npamplexin17 | 104 | 11,873.80 | 8.56 | 1 h | 57.01 | −0.561 |
| Npamplexin18 | 104 | 12,036.82 | 8.40 | 5.5 h | 64.80 | −0.930 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, P.; Gong, Y.; Wang, B.; Yu, H.; Huang, S.; Liao, X.; Jiang, J.; Ran, J.; Xie, F. Love Hug—Functional Validation of Nuptial Pad-Secreted Pheromone in Anurans. Animals 2024, 14, 1550. https://doi.org/10.3390/ani14111550
Zheng P, Gong Y, Wang B, Yu H, Huang S, Liao X, Jiang J, Ran J, Xie F. Love Hug—Functional Validation of Nuptial Pad-Secreted Pheromone in Anurans. Animals. 2024; 14(11):1550. https://doi.org/10.3390/ani14111550
Chicago/Turabian StyleZheng, Puyang, Yuzhou Gong, Bin Wang, Haoqi Yu, Sining Huang, Xun Liao, Jianping Jiang, Jianghong Ran, and Feng Xie. 2024. "Love Hug—Functional Validation of Nuptial Pad-Secreted Pheromone in Anurans" Animals 14, no. 11: 1550. https://doi.org/10.3390/ani14111550
APA StyleZheng, P., Gong, Y., Wang, B., Yu, H., Huang, S., Liao, X., Jiang, J., Ran, J., & Xie, F. (2024). Love Hug—Functional Validation of Nuptial Pad-Secreted Pheromone in Anurans. Animals, 14(11), 1550. https://doi.org/10.3390/ani14111550