
Decline in Size-at-Maturity of European Hake in Relation to Environmental Regimes: A Case in the Eastern Ionian Sea
 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
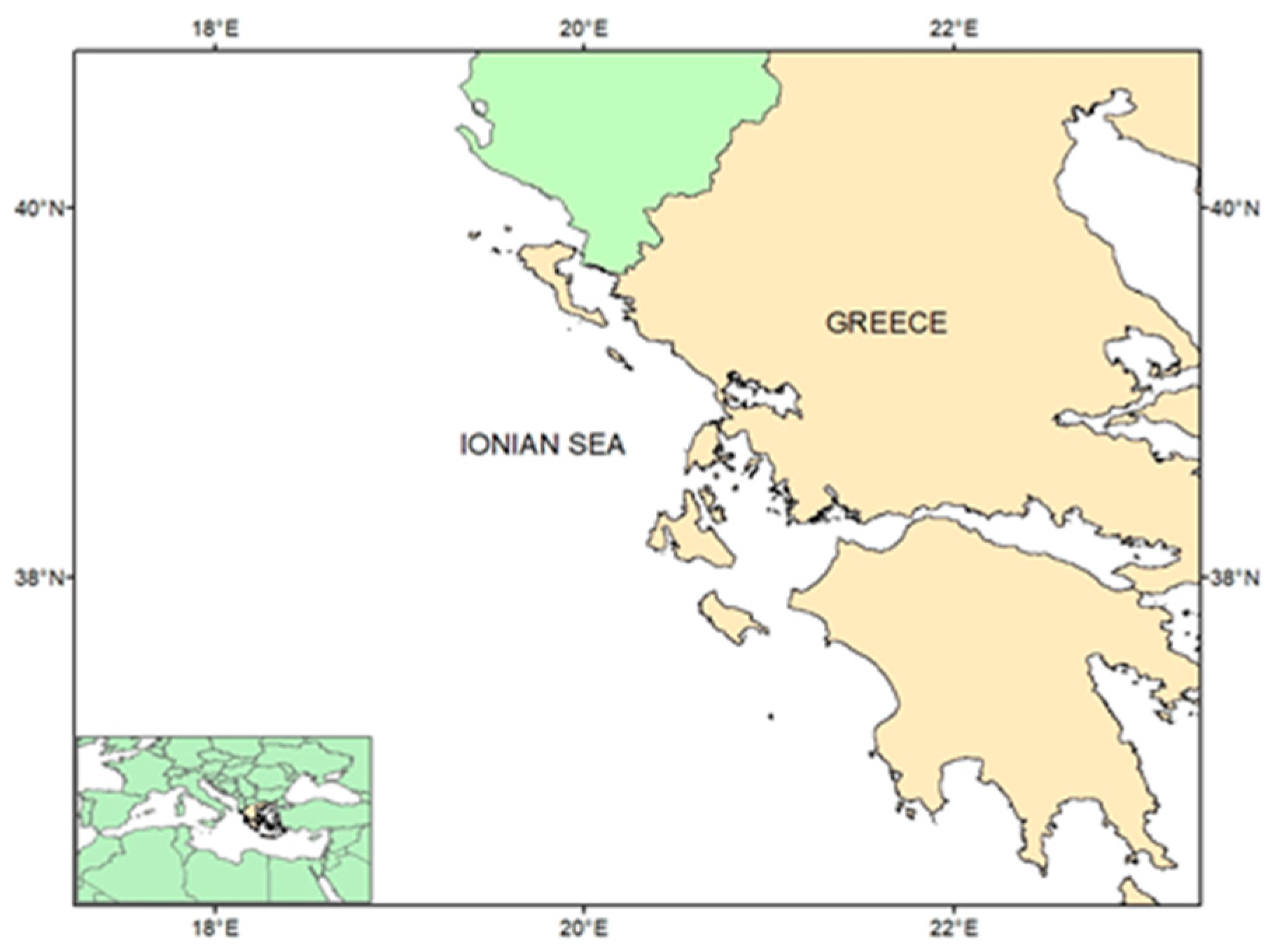
2.1. Study Area
2.2. Data
2.2.1. Biological Data (Size-at-Maturity)
2.2.2. Environmental Data
2.3. Statistical Analysis
2.3.1. Biological Data Analysis
2.3.2. Environmental Data Analysis
3. Results
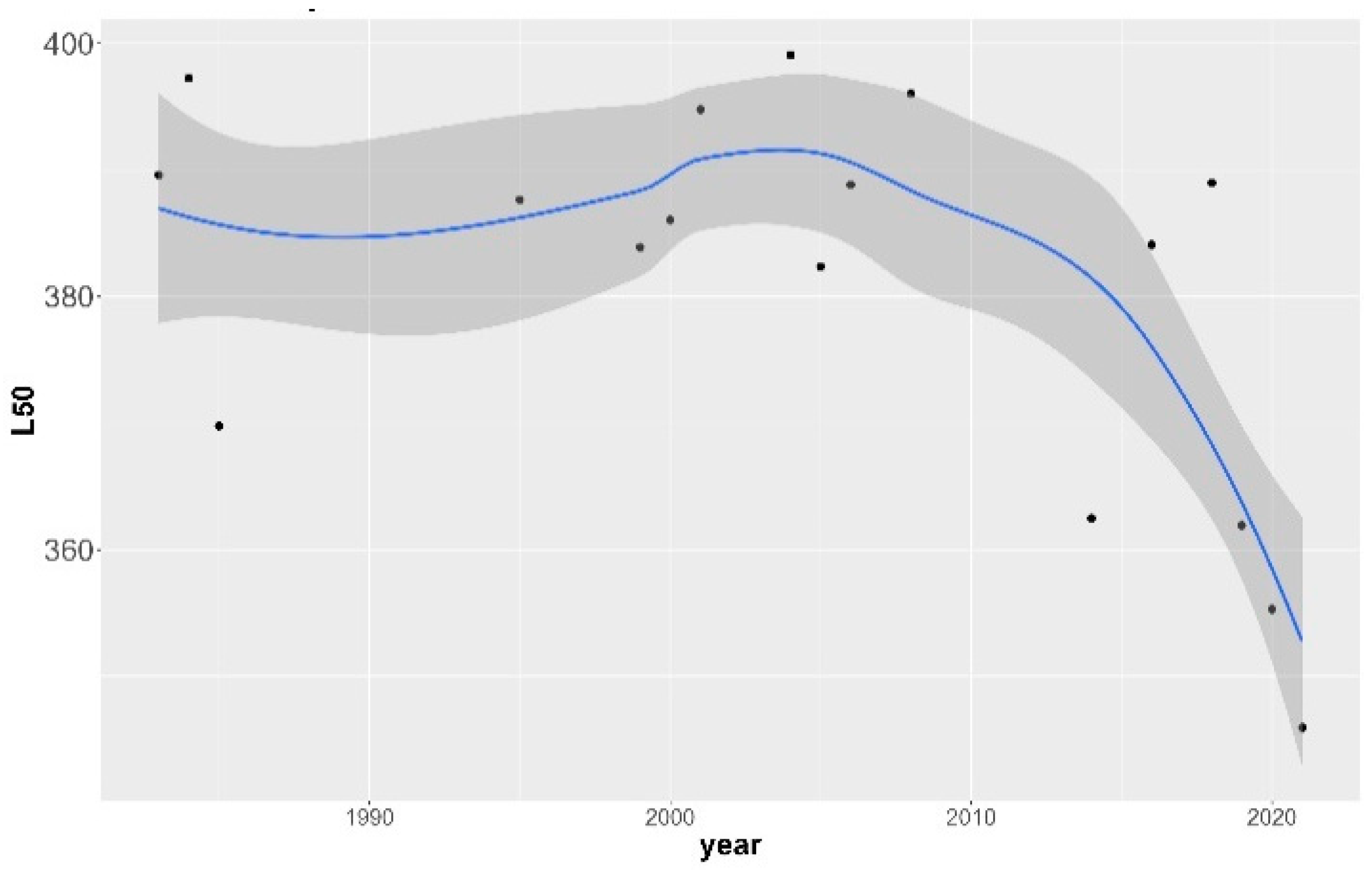
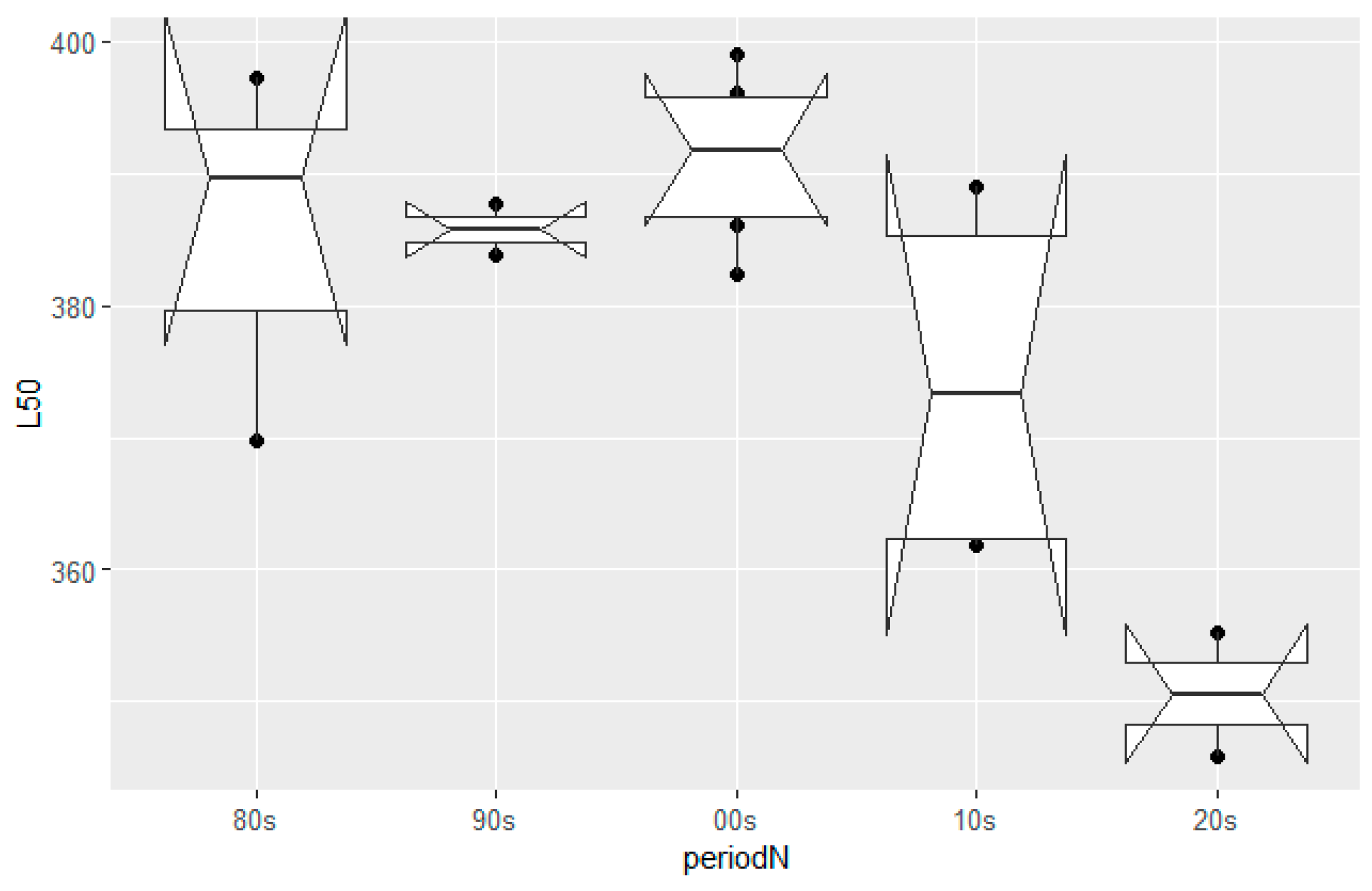
3.1. Size-at-Maturity
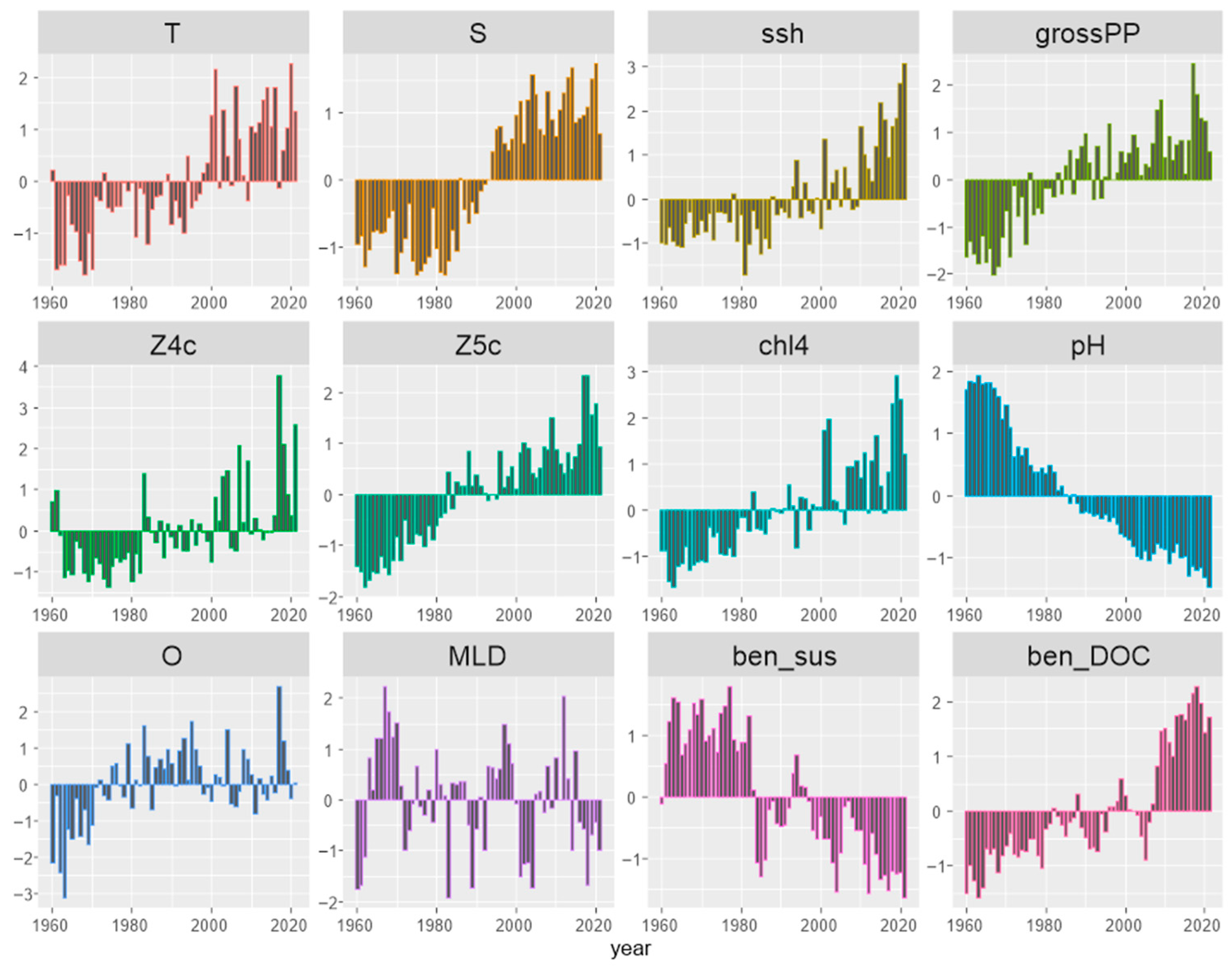
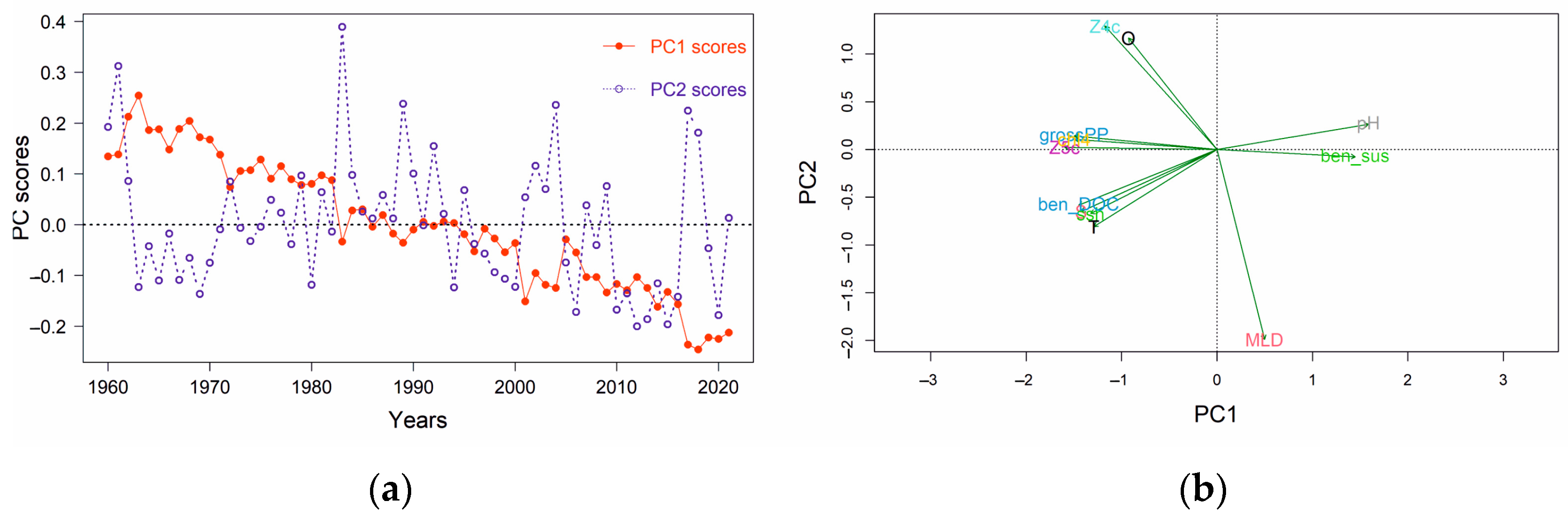
3.2. Environmental ‘Stressors’
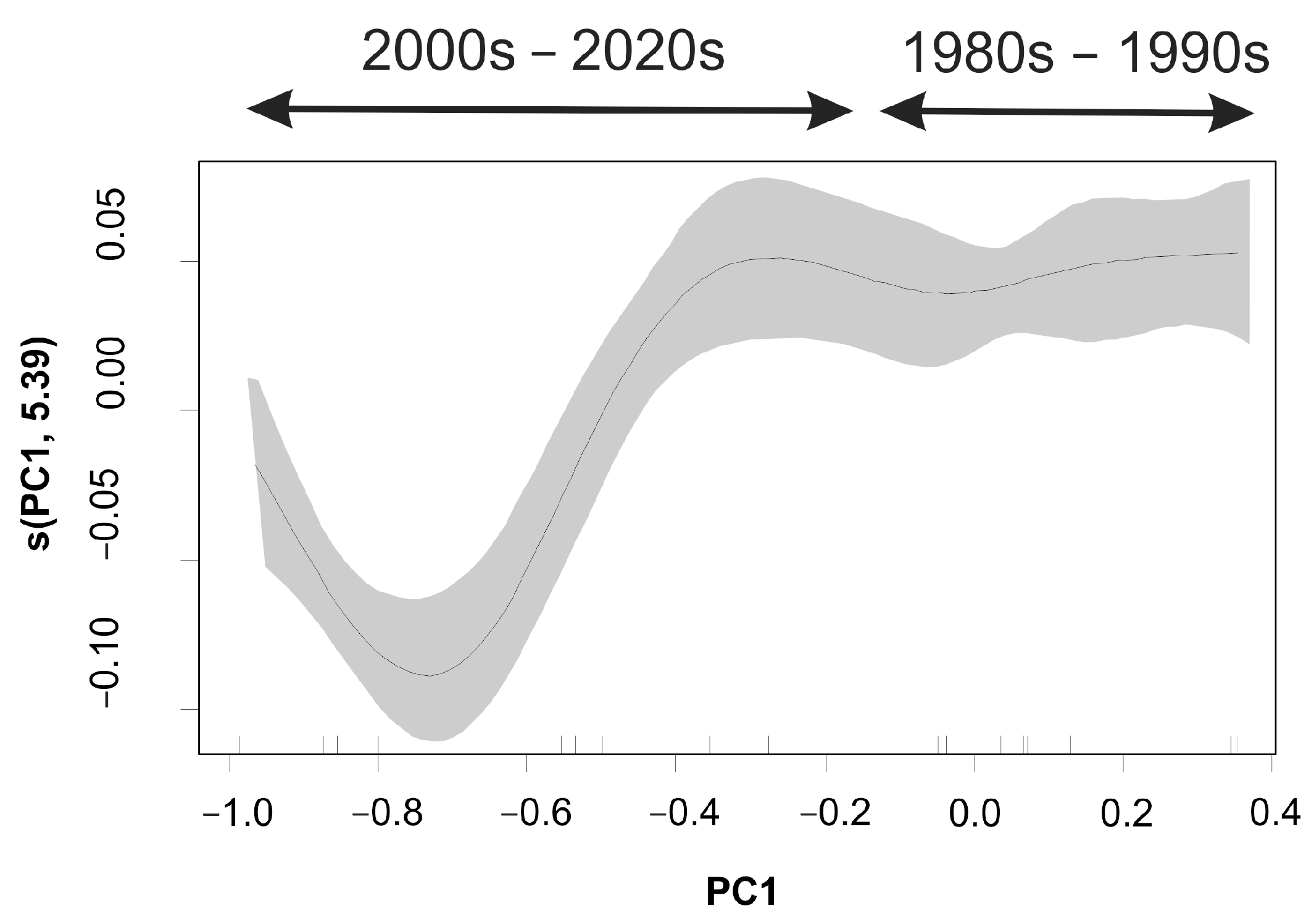
3.3. Maturity–Environmental Stressor Relationship
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jørgensen, C.; Enberg, K.; Dunlop, E.S.; Arlinghaus, R.; Boukal, D.S.; Brander, K.; Ernande, B.; Gårdmark, A.; Johnston, F.; Matsumura, S.; et al. Managing evolving fish stocks. Science 2007, 318, 1247–1248. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Nicieza, A.; Wootton, R.J. Compensatory growth in fishes: A response to growth depression. Fish Fish. 2003, 4, 147–190. [Google Scholar] [CrossRef]
- McNamara, J.M.; Barta, Z.; Klaassen, M.; Bauer, S. Cues and the optimal timing of activities under environmental changes. Ecol. Lett. 2011, 14, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Basilone, G.; Guisande, C.; Patti, B.; Mazzola, S.; Cuttitta, A.; Bonnano, A.; Vergara, A.R.; Maneiro, I. Effect of habitat conditions on reproduction of the European anchovy (Engraulis encrasicolus) in the Strait of Sicily. Fish. Oceanogr. 2006, 15, 271–280. [Google Scholar] [CrossRef]
- Rijnsdorp, A.D. Fisheries as a large-scale experiment on life-history evolution: Disentangling phenotypic and genetic effects in changes in maturation and reproduction of North Sea plaice, Pleuronectes platessa L. Oecologia 1993, 96, 391–401. [Google Scholar] [CrossRef]
- Trippel, E.A. Age at maturity as a stress indicator in fisheries. Bioscience 1995, 45, 759–771. [Google Scholar] [CrossRef]
- Cardinale, M.; Modin, J. Changes in size-at-maturity of Baltic cod (Gadus morhua) during a period of large variations in stock size and environmental conditions. Fish. Res. 1999, 41, 285–295. [Google Scholar] [CrossRef]
- Olsen, E.M.; Heino, M.; Lilly, G.R.; Morgan, M.J.; Brattey, J.; Ernande, B.; Dieckmann, U. Maturation trends indicative of rapid evolution preceded the collapse of northern cod. Nature 2004, 428, 932–935. [Google Scholar] [CrossRef]
- Enberg, K.; Jørgensen, C.; Dunlop, E.S.; Varpe, Ø.; Boukal, D.S.; Baulier, L.; Eliassen, S.; Heino, M. Fishing-induced evolution of growth: Concepts, mechanisms, and the empirical evidence. Mar. Ecol. 2012, 33, 1–25. [Google Scholar] [CrossRef]
- Lojo, D.; Cousido-Rocha, M.; Cerviño, S.; Dominguez-Petit, R.; Sainza, M.; Pennino, M.G. Assessing changes in size at maturity for the European hake (Merluccius merluccius) in Atlantic Iberian waters. Sci. Mar. 2022, 86, e046. [Google Scholar] [CrossRef]
- Koenigstein, S.; Mark, F.C.; Gößling-Reisemann, S.; Reuter, H.; Poertner, H.O. Modelling climate change impacts on marine fish populations: Process-based integration of ocean warming, acidification and other environmental drivers. Fish Fish. 2016, 17, 972–1004. [Google Scholar] [CrossRef]
- Hollins, J.; Thambithurai, D.; Koeck, B.; Crespel, A.; Bailey, D.M.; Cooke, S.J.; Lindström, J.; Parsons, K.J.; Killen, S.S. A physiological perspective on fisheries-induced evolution. Evol. Appl. 2018, 11, 561–576. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M.; Carpenter, S.R. Catastrophic regime shifts in ecosystems: Linking theory to observation. Trends Ecol. Evol. 2003, 18, 648–656. [Google Scholar] [CrossRef]
- Scheffer, M. Critical Transitions in Nature and Society; Princeton University Press: Princeton, NJ, USA, 2009; pp. 1–400. [Google Scholar]
- Palumbi, S.R.; McLeod, K.L.; Grünbaum, D. Ecosystems in action: Lessons from marine ecology about recovery, resistance, and reversibility. BioScience 2008, 58, 33–42. [Google Scholar] [CrossRef]
- Tzanatos, E.; Raitsos, D.E.; Triantafyllou, G.; Somarakis, S.; Tsonis, A.A. Indications of a climate effect on Mediterranean fisheries. Clim. Chang. 2014, 122, 41–54. [Google Scholar] [CrossRef]
- Damalas, D.; Sgardeli, V.; Vasilakopoulos, P.; Tserpes, G.; Maravelias, C. Evidence of climate-driven regime shifts in the Aegean Sea’s demersal resources: A study spanning six decades. Ecol. Evol. 2021, 11, 16951–16971. [Google Scholar] [CrossRef]
- Cohen, D.M.; Inada, T.; Iwamoto, T.; Scialabba, N.; FAO Species Catalogue. Gadiform fishes of the world (Order Gadiformes). An annotated and illustrated catalogue of cods, hakes, grenadiers and other gadiform fishes known to date. FAO Fish. Synop. 1990, 10, 1–442. [Google Scholar]
- Sion, L.; Zupa, W.; Calculli, C.; Garofalo, G.; Hidalgo, M.; Jadaud, A.; Lefkaditou, E.; Ligas, A.; Peristeraki, P.; Bitetto, I.; et al. Spatial distribution pattern of European hake, Merluccius merluccius (Pisces: Merlucciidae), in the Mediterranean Sea. Sci. Mar. 2019, 83, 21–32. [Google Scholar] [CrossRef]
- Recasens, L.; Chiericoni, V.; Belcari, P. Spawning pattern and batch fecundity of the European hake (Merluccius merluccius (Linnaeus, 1758)) in the western Mediterranean. Sci. Mar. 2008, 72, 721–732. [Google Scholar] [CrossRef]
- Carbonara, P.; Porcu, C.; Donnaloia, M.; Pesci, P.; Sion, L.; Spedicato, M.T.; Zupa, W.; Vitale, F.; Follesa, M.C. The spawning strategy of European hake (Merluccius merluccius, L. 1758) across the Western and Central Mediterranean Sea. Fish. Res. 2019, 219, 105333. [Google Scholar] [CrossRef]
- Al-Absawy, M.A. The reproductive biology and the histological and ultrastructural characteristics in ovaries of the female gadidae fish Merluccius merluccius from the Egyptian Mediterranean water. Afr. J. Biotechnol. 2010, 9, 2544–2559. [Google Scholar]
- Khoufi, W.; Ferreri, R.; Jaziri, H.; El Fehri, S.; Gargano, A.; Mangano, S.; Ben Meriem, S.; Romdhane, M.S.; Bonanno, A.; Aronica, S.; et al. Reproductive traits and seasonal variability of Merluccius merluccius from the Tunisian coast. J. Mar. Biol. Assoc. UK 2014, 94, 1545–1556. [Google Scholar] [CrossRef]
- Soykan, O.; İlkyaz, A.T.; Metin, G.; Kınacıgil, H.T. Age, growth and reproduction of European hake (Merluccius merluccius (Linn., 1758)) in the Central Aegean Sea, Turkey. J. Mar. Biol. Assoc. UK 2015, 95, 829–837. [Google Scholar] [CrossRef]
- Belhoucine, F.; Pierson, P.M.; Alioua, A. Characterization of the reproduction of European hake (Merluccius merluccius Linnaeus, 1758) in Oran bay. Appl. Ecol. Environ. Res. 2022, 20, 947–965. [Google Scholar] [CrossRef]
- Biagi, F.; Cesarini, A.; Sbrana, M.; Viva, C. Reproductive biology and fecundity of Merluccius merluccius (Linnaeus, 1758) in the northern Tyrrhenian Sea. Rapp. Comm. Int. Mer Médit. 1995, 34, 237. [Google Scholar]
- Scientific Technical and Economic Committee for Fisheries (STECF). Stock Assessments in the Mediterranean Sea—Adriatic, Ionian and Aegean Seas (STECF-20-15); EUR 28359 EN; Publications Office of the European Union: Luxembourg, 2020; Volume JRC122994, ISBN 978-92-76-27168-0. [CrossRef]
- General Fisheries Commission for the Mediterranean. The State of Mediterranean and Black Sea Fisheries 2022; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- General Fisheries Commission for the Mediterranean (GFCM); Working Group on Stock Assessment of Demersal Species (WGSAD); Scientific Advisory Committee on Fisheries (SAC). Rome, Italy, 12–17 December 2022; 125p. Available online: https://gfcm.sharepoint.com/:b:/g/EG/EWDhdeu5Sc5Djm0HatRs4-UBfVshM38Xhx4DlMlIXsioZg (accessed on 23 May 2023).
- Souvermezoglou, E.; Hatzigeorgiou, E.I.; Pampidis, I.; Siapsali, K. Distribution and seasonal variability of nutrients and dissolved oxygen in the northeastern Ionian Sea. Oceanol. Acta 1992, 15, 585–594. [Google Scholar]
- Theocharis, A.; Klein, B.; Nittis, K.; Roether, W. Evolution and status of the Eastern Mediterranean Transient (1997–1999). J. Mar. Syst. 2002, 33–34, 91–116. [Google Scholar] [CrossRef]
- Kavadas, S.; Damalas, D.; Georgakarakos, S.; Maravelias, C.; Tserpes, G.; Papaconstantinou, C.; Bazigos, G. IMAS-Fish: Integrated MAnagement System to Support the Sustainability of Greek Fisheries Resources. A Multidisciplinary Web-Based Database Management System: Implementation, Capabilities, Utilization and Future Prospects for Fisheries Stakeholders. Mediterr. Mar. Sci. 2013, 14, 109–118. [Google Scholar] [CrossRef]
- Nikolsky, G.V. The Ecology of Fishes; Academic Press: London, UK, 1963. [Google Scholar]
- Anon. Campagne internationale de chalutage démersal en Méditerranée (MEDITS): Manuel des protocoles. Biol. Mar. Medit. 1998, 5, 515–572. [Google Scholar]
- CERES Project. Available online: http://ceresproject.eu/ (accessed on 10 May 2023).
- Holt, J.T.; James, I.D.; Jones, J.E. An s coordinate density evolving model of the northwest European continental shelf 1, Model description and density structure. J. Geophys. Res. Ocean. 2001, 106, 14015–14034. [Google Scholar] [CrossRef]
- Baretta, J.W.; Ebenhöh, W.; Ruardij, P. The European Regional Seas Ecosystem Model, A Complex Marine Ecosystem Model. Neth. J. Sea Res. 1995, 33, 233–246. [Google Scholar] [CrossRef]
- Butenschön, M.; Clark, J.; Aldridge, J.N.; Allen, J.I.; Artioli, Y.; Blackford, J.; Bruggeman, J.; Cazenave, P.; Ciavatta, S.; Kay, S.; et al. ERSEM 15.06: A generic model for marine biogeochemistry and the ecosystem dynamics of the lower trophic levels. Geosci. Model Dev. 2016, 9, 1293–1339. [Google Scholar] [CrossRef]
- Holt, J.; Butenschön, M.; Wakelin, S.L.; Artioli, Y.; Allen, J.I. Oceanic controls on the primary production of the northwest European continental shelf: Model experiments under recent past conditions and a potential future scenario. Biogeosciences 2012, 9, 97–117. [Google Scholar] [CrossRef]
- Kay, S.; Butenschön, M. Projections of change in key ecosystem indicators for planning and management of marine protected areas: An example study for European seas. Estuar. Coast. Shelf Sci. 2016, 201, 172–184. [Google Scholar] [CrossRef]
- CERES. Deliverable D1.3. Projections of Physical and Biogeochemical Parameters and Habitat Indicators for European Seas, Including Synthesis of Sea Level Rise and Storminess. EU H2020 CERES Project. Climate Change and European Aquatic RESources. 2018. 64p. Available online: https://ec.europa.eu/research/participants/documents/downloadPublic?documentIds=080166e5b9fdf8fb&appId=PPGMS (accessed on 10 May 2023).
- Alemany, D.; Acha, E.M.; Iribarne, O.O. Marine fronts are important fishing areas for demersal species at the Argentine Sea (Southwest Atlantic Ocean). J. Sea Res. 2014, 87, 56–67. [Google Scholar] [CrossRef]
- Druon, J.N.; Fiorentino, F.; Murenu, M.; Knittweis, L.; Colloca, F.; Osio, C.; Mérigot, B.; Garofalo, G.; Mannini, A.; Jadaud, A.; et al. Modelling of European hake nurseries in the Mediterranean Sea: An ecological niche approach. Prog. Oceanogr. 2015, 130, 188–204. [Google Scholar] [CrossRef]
- Cheung, W.W.L.; Watson, R.; Pauly, D. Signature of ocean warming in global fisheries catch. Nature 2013, 497, 365–368. [Google Scholar] [CrossRef]
- Möllmann, C.; Diekmann, R. Marine ecosystem regime shifts induced by climate and overfishing: A review for the northern hemisphere. Adv. Ecol. Res. 2012, 47, 303–346. [Google Scholar] [CrossRef]
- Szekely, G.J.; Rizzo, M.L.; Bakirov, N.K. Measuring and Testing Dependence by Correlation of Distances. Ann. Stat. 2007, 35, 2769–2794. [Google Scholar] [CrossRef]
- ICES. Report of the Workshop on Maturity Ogive Estimation for Stock Assessment (WKMOG). In Proceedings of the Workshop on Maturity Ogive Estimation for Stock Assessment (WKMOG), Lisbon, Portugal, 3–6 June 2008; p. 72. [Google Scholar]
- Pattoura, P.; Mytilineou, C.; Anastasopoulou, A.; Adamidou, A.; Dogrammatzi, A.; Georgiadis, K. Aspects of reproductive biology of European hake (Merluccius merluccius) in the Aegean and Ionian Sea. In Proceedings of the 16th Hellenic Conference of Ichthyologists, Kavala, Greece, 6–9 October 2016; pp. 149–152. [Google Scholar]
- Domínguez-Petit, R.; Korta, M.; Saborido-Rey, F.; Murua, H.; Sainza, M.; Piñeiro, C. Changes in size at maturity of European hake Atlantic populations in relation with stock structure and environmental regimes. J. Mar. Syst. 2008, 71, 260–278. [Google Scholar] [CrossRef]
- Rao, C. The use and interpretation of principal component analysis in applied research. Sankhyā Indian J. Stat. 1964, 26, 329–358. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical ecology. In Developments in Environmental Modelling, 2nd ed.; Elsevier Science BV: Amsterdam, The Netherlands, 1998; Volume 20. [Google Scholar]
- Choi, J.; Frank, K.; Petrie, B.; Leggett, W. Integrated assessment of a large marine ecosystem: A case study of the devolution of the eastern Scotian Shelf, Canada. Oceanogr. Mar. Biol. Annu. Rev. 2005, 43, 67–93. [Google Scholar]
- Kenny, A.; Skjoldal, H.; Engelhard, G.; Kershaw, P.; Reid, J. An integrated approach for assessing the relative significance of human pressures and environmental forcing on the status of large marine ecosystems. Prog. Oceanogr. 2009, 81, 132–148. [Google Scholar] [CrossRef]
- Möllmann, C.; Diekmann, R.; Müller-Karulis, B.; Kornilovs, G.; Plikshs, M.; Axe, P. Reorganization of a large marine ecosystem due to atmospheric and anthropogenic pressure: A discontinuous regime shift in the central Baltic Sea. Glob. Chang. Biol. 2009, 15, 1377–1393. [Google Scholar] [CrossRef]
- Hastie, T. Gam: Generalized Additive Models. R Package Version 1.14-4. 2017. Available online: https://CRAN.R-project.org/package=gam (accessed on 18 May 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 20 May 2023).
- Rizzo, M.L.; Szekely, G.J. Energy: E-Statistics: Multivariate Inference via the Energy of Data. R Package Version 1.7-4. 2018. Available online: https://CRAN.R-project.org/package=energy (accessed on 22 May 2023).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-2. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 2 June 2023).
- Wood, S.N. Generalized Additive Models and Introduction with R, 2nd ed.; Chapman and Hall/CRC: London, UK, 2017. [Google Scholar]
- Lappalainen, A.; Saks, L.; Sustar, M.; Heikinheimo, O.; Jürgens, K.; Kokkonen, E.; Kurkilahti, M.; Verliin, A.; Vetema, M. Length at maturity as a potential indicator of fishing pressure effects on coastal pikeperch (Sander lucioperca) stocks in the northern Baltic Sea. Fish. Res. 2016, 174, 47–57. [Google Scholar] [CrossRef]
- Reznick, D.N.; Shaw, F.H.; Rodd, F.H.; Shaw, R.G. Evaluation of the rate of evolution in natural populations of guppies (Poecilia reticulata). Science 1997, 275, 1934–1937. [Google Scholar] [CrossRef] [PubMed]
- Rutterford, L.A.; Genner, M.J.; Engelhard, G.H.; Simpson, S.D.; Hunter, E. Fishing impacts on age structure may conceal environmental drivers of body size in exploited fish populations. ICES J. Mar. Sci. 2023, 80, 848–860. [Google Scholar] [CrossRef]
- Habouz, H.; Recasens, L.; Kifani, S.; Moukrim, A.; Bouhaimi, A.; Ayoubi, S. Maturity and batch fecundity of the European hake (Merluccius merluccius, Linnaeus, 1758) in the eastern central Atlantic. Sci. Mar. 2011, 75, 447–454. [Google Scholar] [CrossRef]
- El Habouz, H. Etude de la Biologie et la Dynamique des Populations du merlu blanc (Merluccius merluccius, Linnaeus 1758) Débarqué par les Chalutiers Côtiers au port d’Agadir. Master’s Thesis, Option Océanographie Biologique, University Ibn Zohr, Agadir, Morocco, 1995. [Google Scholar]
- Marty, L.; Rochet, M.J.; Ernande, B. Temporal trends in age and size at maturation of four North Sea gadid species: Cod, haddock, whiting and Norway pout. Mar. Ecol. Prog. Ser. 2014, 497, 179–197. [Google Scholar] [CrossRef]
- Kokkonen, E.; Vainikka, A.; Heikinheimo, O. Probabilistic maturation reaction norm trends reveal decreased size and age at maturation in an intensively harvested stock of pikeperch Sander lucioperca. Fish. Res. 2015, 167, 1–12. [Google Scholar] [CrossRef]
- Arula, T.; Shpilev, H.; Raid, T.; Vetemaa, M.; Albert, A. Maturation at a young age and small size of European smelt (Osmerus eperlanus): A consequence of population overexploitation or climate change? Helgol. Mar. Res. 2017, 71, 7. [Google Scholar] [CrossRef]
- Shapiro Goldberg, D.; van Rijn, I.; Kiflawi, M.; Belmaker, J. Decreases in Length at Maturation of Mediterranean Fishes Associated with Higher Sea Temperatures. ICES J. Mar. Sci. 2019, 76, 946–959. [Google Scholar] [CrossRef]
- Zervakis, V.; Georgopoulos, D.; Drakopoulos, P.G. The role of the north Aegean in triggering the recent eastern Mediterranean climatic changes. J. Geophys. Res. Ocean. 2000, 105, 26103–26116. [Google Scholar] [CrossRef]
- Drinkwater, K.F.; Beaugrand, G.; Kaeriyama, M.; Kim, S.; Ottersen, G.; Perry, R.I.; Pörtner, H.O.; Polovina, J.J.; Takasuka, A. On the processes linking climate to ecosystem changes. J. Mar. Syst. 2010, 79, 374–388. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, S.; Niu, X. Effect of water temperature on the dynamic behavior of phytoplankton–zooplankton model. Appl. Math. Comput. 2020, 378, 125211. [Google Scholar] [CrossRef]
- Paradinas, I.; Chust, G.; Garcia, D.; Ibaibarriaga, L. Large scale spatio-temporal study on European hake depicts sensitivity to sea bottom temperature, sea bottom oxygen concentration and sea surface temperature. bioRxiv 2023, Preprint. [Google Scholar] [CrossRef]
- Niu, J.; Huss, M.; Vasemägi, A.; Gårdmark, A. Decades of Warming Alters Maturation and Reproductive Investment in Fish. Ecosphere 2023, 14, e4381. [Google Scholar] [CrossRef]
- Chittaro, P.; Grandin, C.; Pacunski, R.; Zabel, R. Five decades of change in somatic growth of Pacific hake from Puget Sound and Strait of Georgia. PeerJ 2022, 10, e13577. [Google Scholar] [CrossRef]
- Boeuf, G.; Payan, P. How should salinity influence fish growth? Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2001, 130, 411–423. [Google Scholar] [CrossRef]
- Panfili, J.; Thior, D.; Ecoutin, J.M.; Ndiaye, P.; Albaret, J.J. Influence of salinity on the size at maturity for fish species reproducing in contrasting West African estuaries. J. Fish Biol. 2006, 69, 95–113. [Google Scholar] [CrossRef]
- Esbaugh, A.J. Physiological implications of ocean acidification for marine fish: Emerging patterns and new insights. J. Comp. Physiol. B. 2018, 188, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nagelkerken, I.; Alemany, T.; Anquetin, J.M.; Ferreira, C.M.; Ludwig, K.E.; Sasaki, M.; Connell, S.D. Ocean acidification boosts reproduction in fish via indirect effects. PLoS Biol. 2021, 19, e3001033. [Google Scholar] [CrossRef] [PubMed]
- Kuparinen, A.; Merilä, J. Detecting and managing fisheries-induced evolution. Trends Ecol. Evol. 2007, 22, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Flynn, E.E.; Bjelde, B.E.; Miller, N.A.; Todgham, A.E. Ocean acidification exerts negative effects under warming conditions in a developing Antarctic fish. Conserv. Physiol. 2015, 3, cov033. [Google Scholar] [CrossRef]
- Ferrer-Maza, D.; Lloret, J.; Munoz, M.; Faliex, E.; Vila, S.; Sasal, P. Parasitism, condition and reproduction of the European hake (Merluccius merluccius) in the northwestern Mediterranean Sea. ICES J. Mar. Sci. 2014, 71, 1088–1099. [Google Scholar] [CrossRef]
- Law, R. Fisheries-induced evolution: Present status and future directions. Mar. Ecol. Prog. Ser. 2007, 335, 271–277. [Google Scholar] [CrossRef]
- Ministry of Agricultural Development & Food-General Directorate of Fisheries—Greek Republic. Annual Report of the Fishing Fleet for the Reference Year 2022 According to Article 22 of Regulation (EC) no. 1380/2013 of the European Parliament and the Council (In Greek). 37p. 2023. Available online: http://www.alieia.minagric.gr/sites/default/files/basicPageFiles/%CE%95%CE%A4%CE%97%CE%A3%CE%99%CE%91%20%CE%95%CE%9A%CE%98%CE%95%CE%A3%CE%97%20%CE%A3%CE%A4%CE%9F%CE%9B%CE%9F%CE%A5%202022.pdf (accessed on 15 June 2023).
- Rouyer, T.; Fromentin, J.-M.; Hidalgo, M.; Stenseth, N.C. Combined effects of exploitation and temperature on fish stocks in the Northeast Atlantic. ICES J. Mar. Sci. 2014, 71, 1554–1562. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Units | Description |
|---|---|---|
| T | °C | Sea surface temperature |
| S | psu | Salinity |
| ssh | m | Sea surface height |
| grossPP | tons C/day | Gross primary production |
| Z4c | mg C/m3 | Mesozooplankton C |
| Z5c | mg C/m3 | Microzooplankton C |
| chl4 | mg/m3 | Microphytoplankton Chl |
| pH | −log[H+] | Degree of acidity/alkalinity |
| O | mmol/m3 | Dissolved oxygen |
| ben_sus | mg/m2 | Suspension/filter feeders, macrobenthos |
| ben_DOC | mg/m2 | Benthic dissolved organic carbon |
| MLD | m | Mixed layer depth |
| Variable | % PC1 Contribution |
|---|---|
| Z5c | 10.2 |
| pH | 10.1 |
| grossPP | 9.5 |
| chl4 | 9.5 |
| ben_sus | 9.2 |
| ben_DOC | 9.2 |
| S | 9.1 |
| ssh | 8.4 |
| T | 8.2 |
| Z4c | 7.5 |
| Family: Gamma | Link Function: log | |||
|---|---|---|---|---|
| Formula: L50 ~ s(PC1, k = 7) | ||||
| Parameter | Edf | Ref.df | F | p-Value |
| s(PC1) | 5.392 | 5.859 | 6.907 | 0.00313 |
| R-sq.(adj) = 0.696 | ||||
| Deviance explained = 80.2% | ||||
| GCV = 0.00085265 | ||||
| Scale est. = 0.00053114, n = 17 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legaki, A.; Chatzispyrou, A.; Damalas, D.; Sgardeli, V.; Lefkaditou, E.; Anastasopoulou, A.; Dogrammatzi, A.; Charalampous, K.; Stamouli, C.; Vassilopoulou, V.; et al. Decline in Size-at-Maturity of European Hake in Relation to Environmental Regimes: A Case in the Eastern Ionian Sea. Animals 2024, 14, 61. https://doi.org/10.3390/ani14010061
Legaki A, Chatzispyrou A, Damalas D, Sgardeli V, Lefkaditou E, Anastasopoulou A, Dogrammatzi A, Charalampous K, Stamouli C, Vassilopoulou V, et al. Decline in Size-at-Maturity of European Hake in Relation to Environmental Regimes: A Case in the Eastern Ionian Sea. Animals. 2024; 14(1):61. https://doi.org/10.3390/ani14010061
Chicago/Turabian StyleLegaki, Aglaia, Archontia Chatzispyrou, Dimitrios Damalas, Vasiliki Sgardeli, Evgenia Lefkaditou, Aikaterini Anastasopoulou, Aikaterini Dogrammatzi, Konstantinos Charalampous, Caterina Stamouli, Vassiliki Vassilopoulou, and et al. 2024. "Decline in Size-at-Maturity of European Hake in Relation to Environmental Regimes: A Case in the Eastern Ionian Sea" Animals 14, no. 1: 61. https://doi.org/10.3390/ani14010061
APA StyleLegaki, A., Chatzispyrou, A., Damalas, D., Sgardeli, V., Lefkaditou, E., Anastasopoulou, A., Dogrammatzi, A., Charalampous, K., Stamouli, C., Vassilopoulou, V., Tserpes, G., & Mytilineou, C. (2024). Decline in Size-at-Maturity of European Hake in Relation to Environmental Regimes: A Case in the Eastern Ionian Sea. Animals, 14(1), 61. https://doi.org/10.3390/ani14010061