

First Records of Picobiine Mites Associated with Birds-of-Paradise: Can Interspecific Sexual Behaviour of Hosts Play a Role in the Distribution of Quill Mite Parasites? †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Systematics
3.1.1. Descriptions
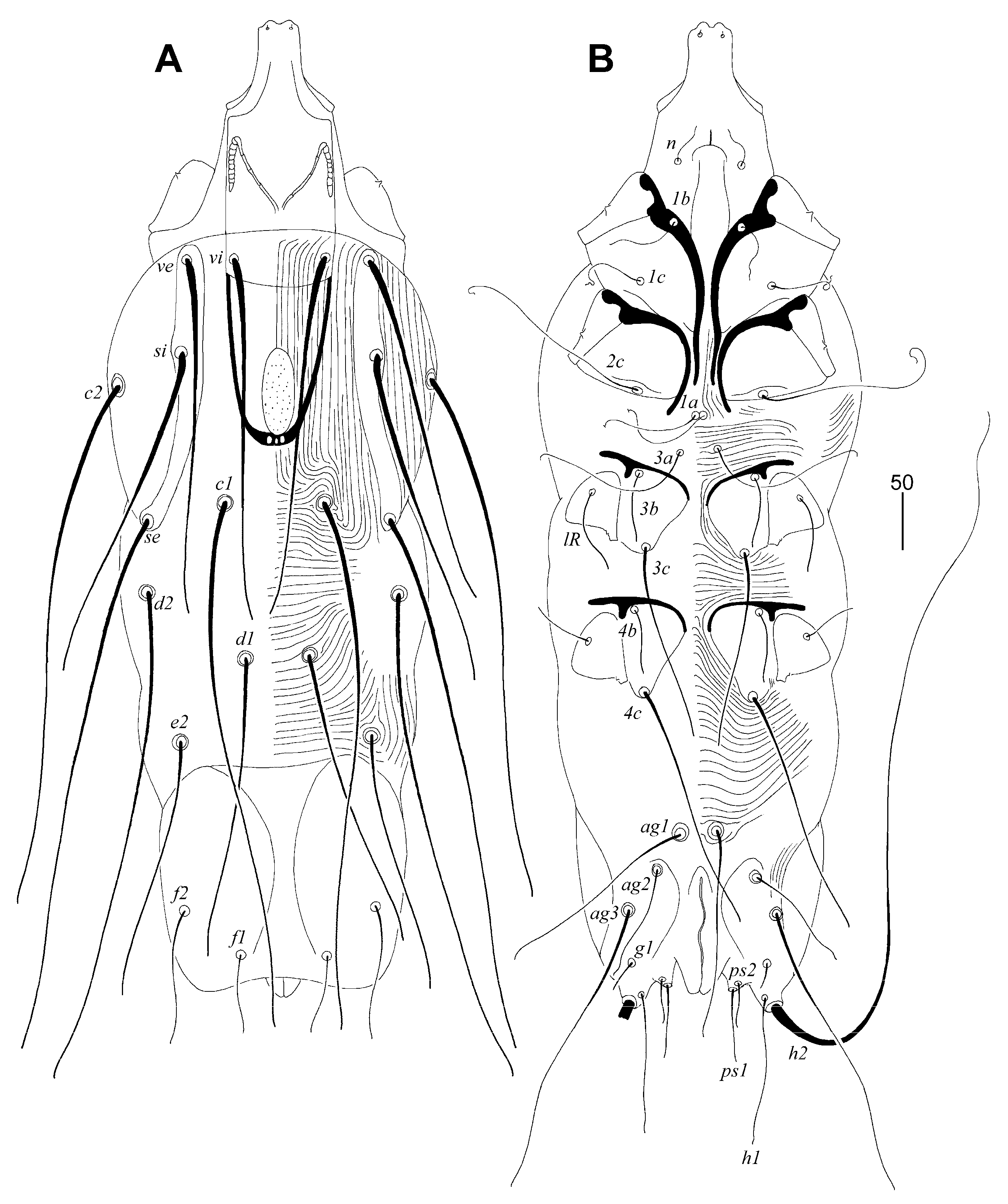
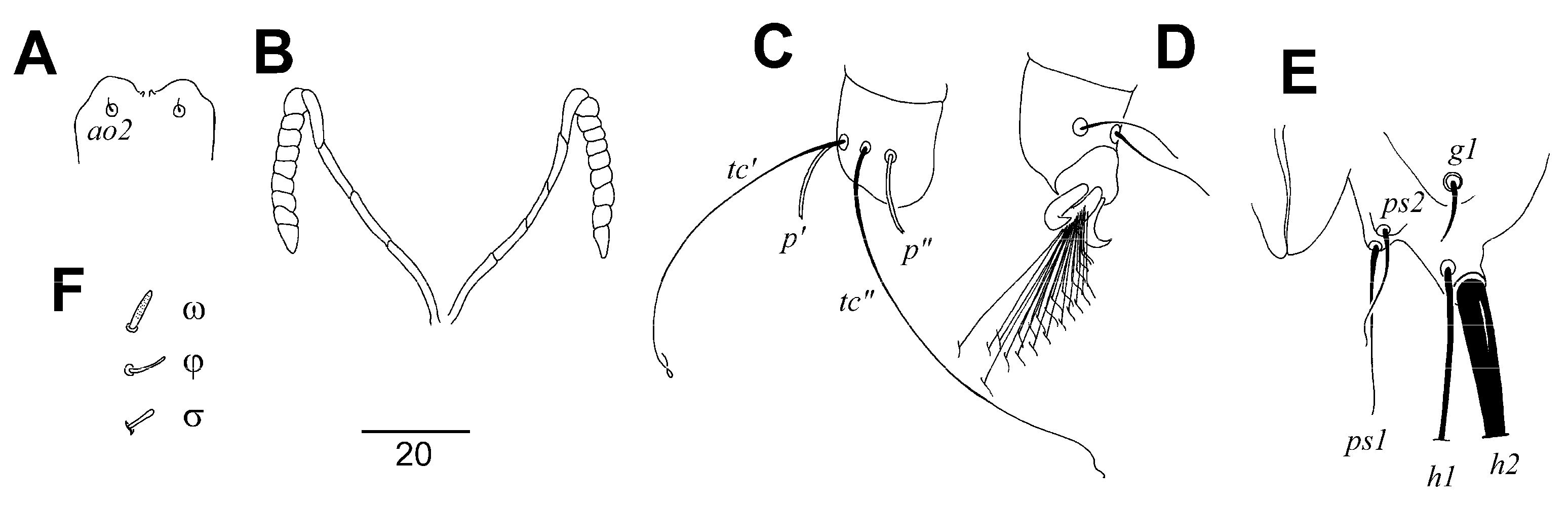
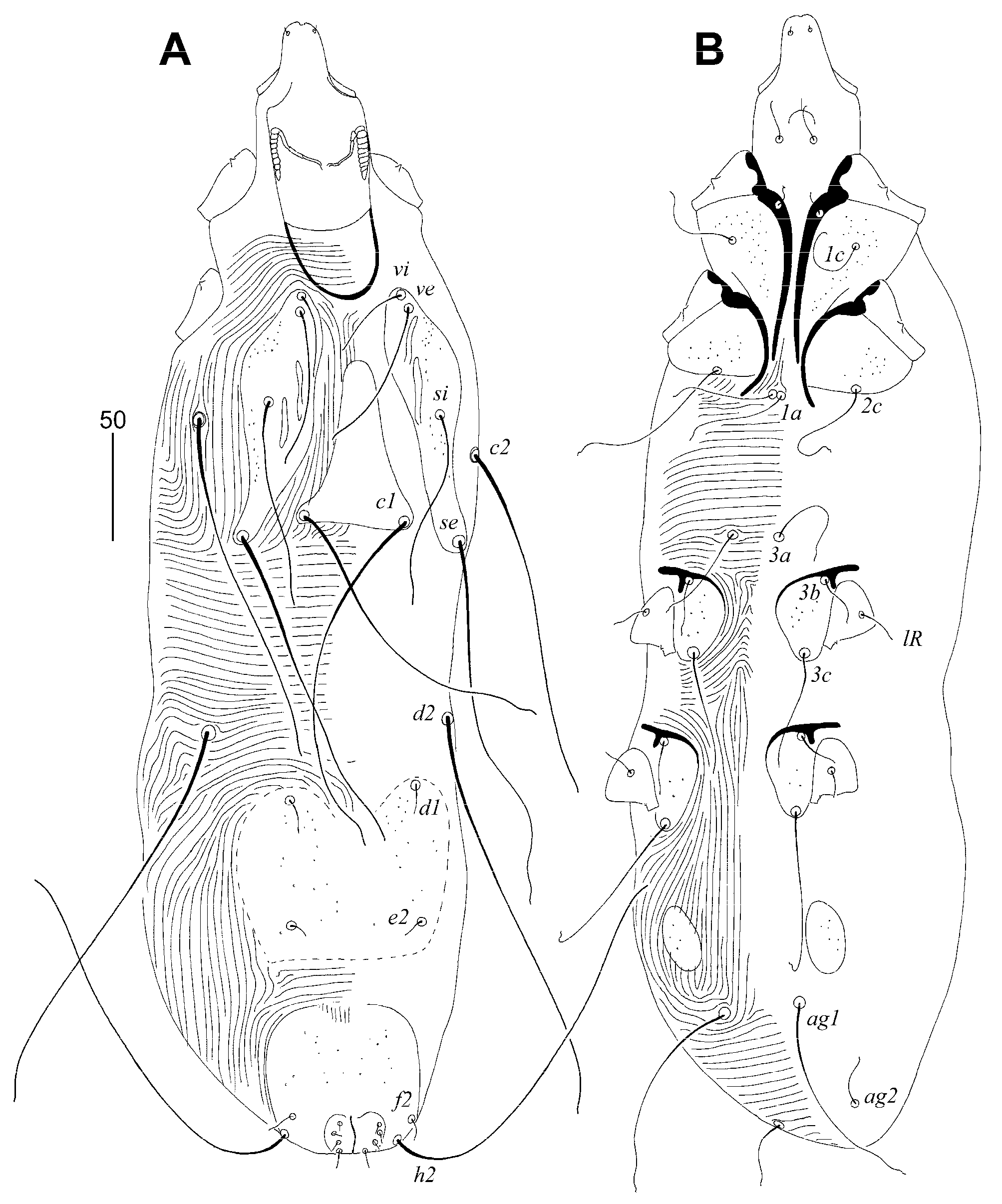
Picobia frankei sp. n. (Figure 1, Figure 2 and Figure 3)
Hosts and Distribution
Type Material
Type Material Deposition
Additional Material
Differential Diagnosis
Etymology
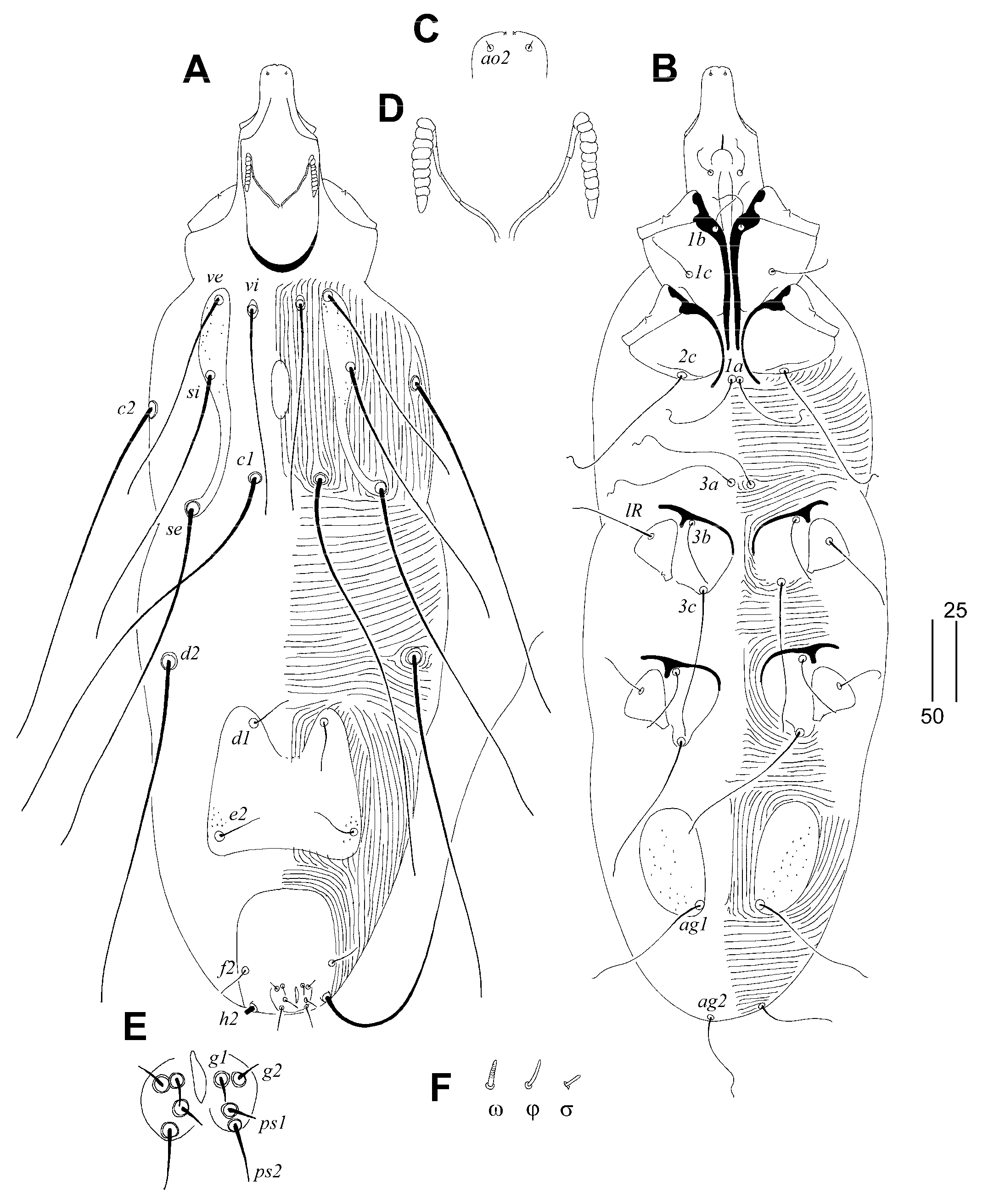
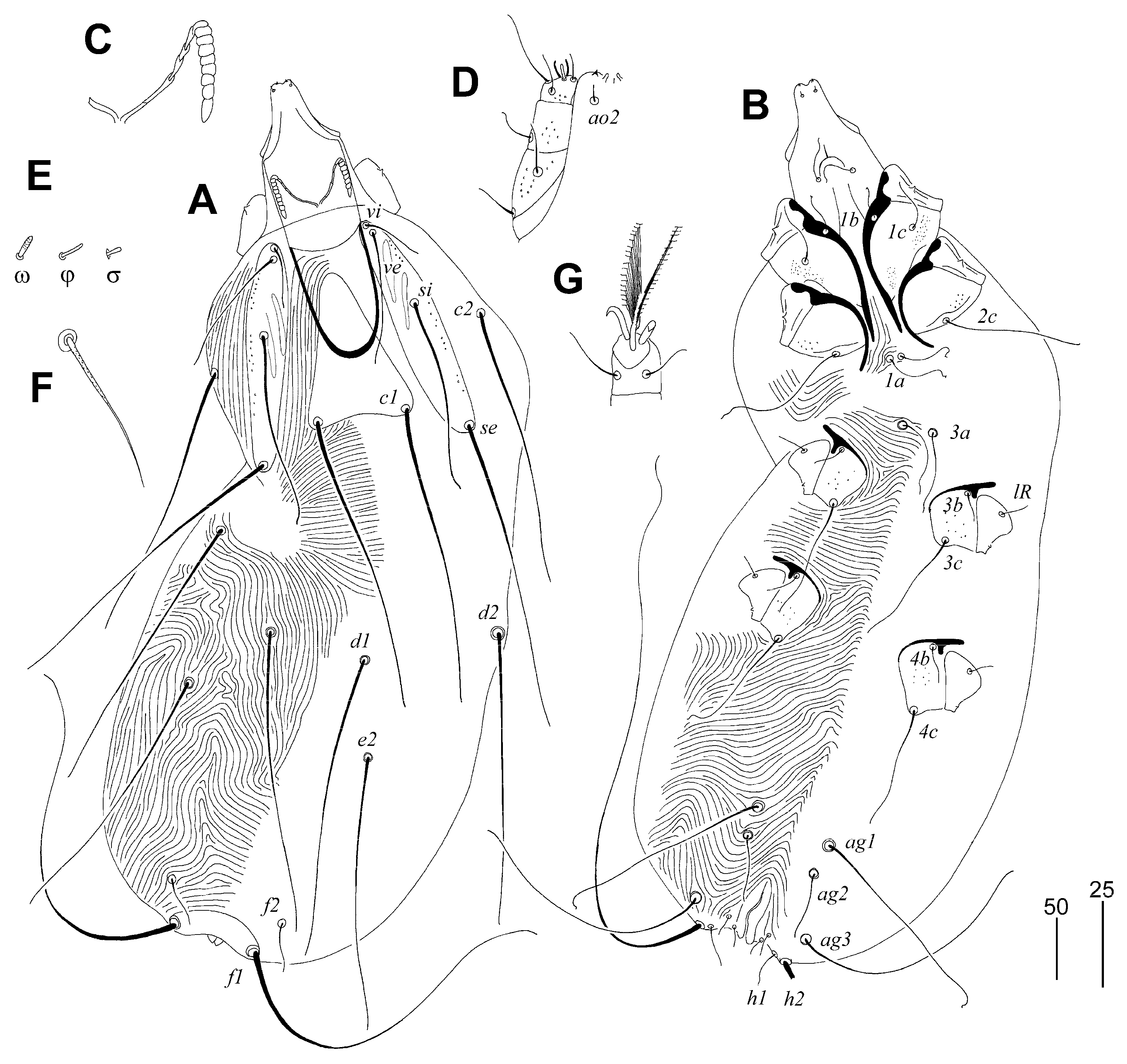
Gunabopicobia garylarsoni sp. n. (Figure 5 and Figure 6)
Hosts and Distribution
Type Material
Type Material Deposition
Additional Material
Differential Diagnosis
Etymology
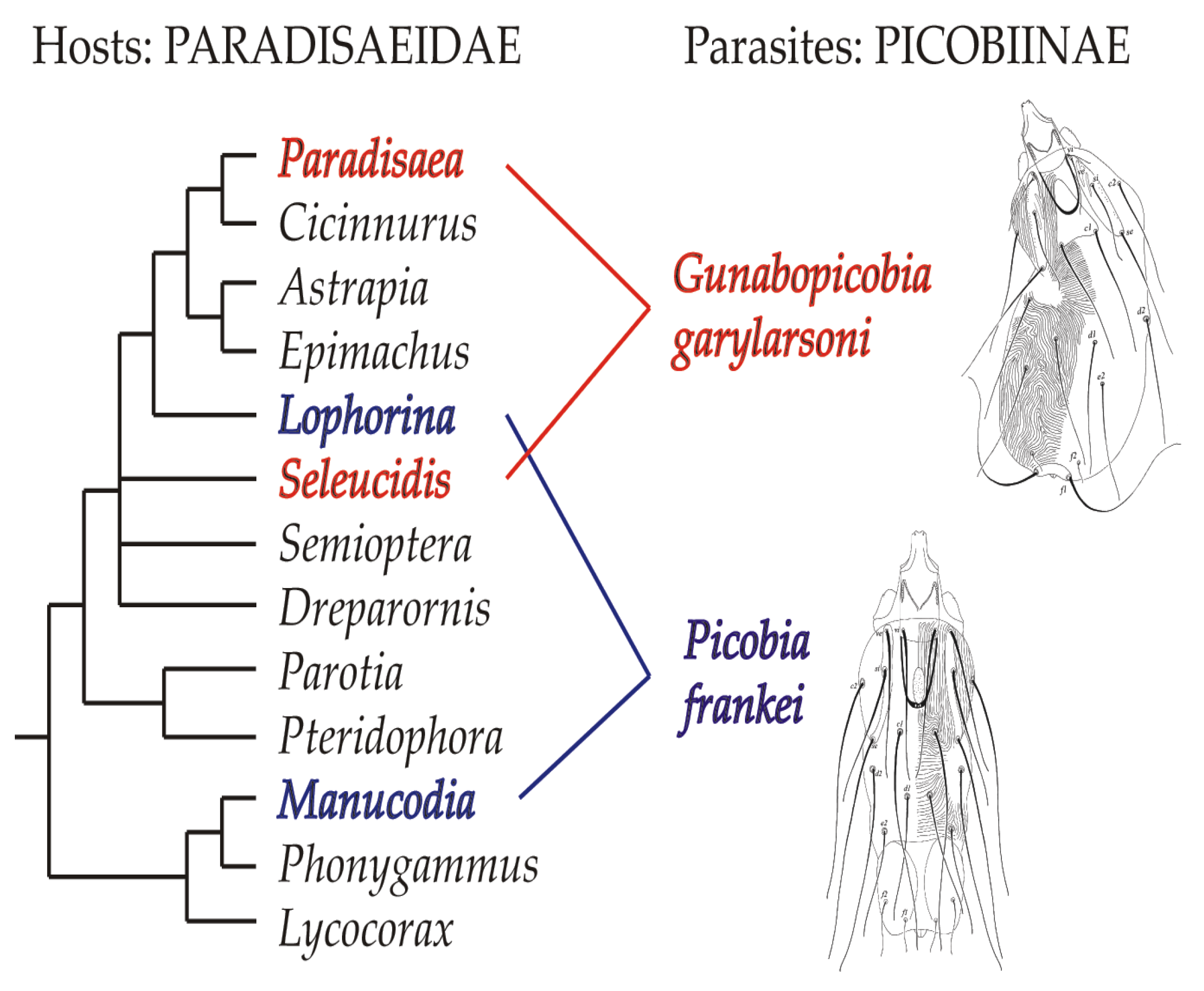
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Bird Families of the World; Lynx Edicions: Barcelona, Spain, 2015; pp. 339–391. [Google Scholar]
- del Hoyo, J. (Ed.) All the Birds of the World; Lynx Edicions: Barcelona, Spain, 2020; pp. 544–566. [Google Scholar]
- Jønsson, K.A.; Christidis, L.; Cibois, A.; Fuchs, J.; Irestedt, M.; Kennedy, J.D.; Fjeldså, J. Cohort Corvides: The crow-like passerines. In The Largest Avian Radiation. The Evolution of Perching Birds, or the Order Passeriformes; Fjeldså, J., Christidis, L., Ericson, P.G.P., Eds.; Lynx Edicions: Barcelona, Spain, 2020; Chapter 8; pp. 131–168. [Google Scholar]
- Kethley, J.B. A revision of the family Syringophilidae (Prostigmata: Acarina). Contrib. Am. Entomol. Inst. 1970, 6, 1–76. [Google Scholar]
- Johnston, D.E.; Kethley, J.B. A numerical phenetic study of the quill mites of the family Syringophilidae (Acari). J. Parasitol. 1973, 59, 520–530. [Google Scholar] [CrossRef]
- Skoracki, M. Quill mites (Acari: Syringophilidae) of the Palaearctic region. Zootaxa 2011, 2840, 1–414. [Google Scholar] [CrossRef]
- Skoracki, M.; Sikora, B.; Spicer, G.S. A review of the subfamily Picobiinae Johnston and Kethley, 1973 (Acariformes: Prostigmata: Syringophilidae). Zootaxa 2016, 4113, 1–95. [Google Scholar] [CrossRef] [PubMed]
- Skoracki, M.; Sikora, B.; Jerzak, L.; Hromada, M. Tanopicobia gen. nov., a new genus of quill mites, its phylogenetic placement in the subfamily Picobiinae (Acariformes: Syringophilidae) and picobiine relationships with avian hosts. PLoS ONE 2020, 15, e0225982. [Google Scholar] [CrossRef] [PubMed]
- Zmudzinski, M.; Skoracki, M.; Sikora, B. An Updated Checklist of Quill Mites of the Family Syringophilidae (Acariformes: Prostigmata). 2021. Available online: https://figshare.com/articles/dataset/An_updated_checklist_of_quill_mites_of_the_family_Syringophilidae_Acariformes_Prostigmata_/16529574/1 (accessed on 1 March 2023).
- Grandjean, F. Les segments post-larvaires de l’hystérosoma chez les Oribates (Acariens). Bull. Soc. Zool. Fr. 1939, 64, 273–284. [Google Scholar]
- Kethley, J.B. Acarina: Prostigmata (Actinedida). In Soil Biology Guide; Dindal, D.L., Ed.; John Wiley & Sons: New York, NY, USA, 1990; pp. 667–756. [Google Scholar]
- Grandjean, F. Observations sur les Acariens de la famille des Stigmaeidae. Arch. Sci. Phys. Nat. 1944, 26, 103–131. [Google Scholar]
- Klimovičová, M.; Skoracki, M.; Wamiti, W.; Hromada, M. Quill mites of the subfamily Picobiinae (Acari: Syringophilidae) parasitising African birds, with description of two new species. Folia Parasitol. 2014, 61, 394–400. [Google Scholar] [CrossRef]
- Kaszewska, K.; Skoracki, M.; Hromada, M. A review of the quill mites of the genus Gunabopicobia Skoracki and Hromada (Acariformes: Prostigmata: Syringophilidae) associated with birds of the order Columbiformes. Int. J. Acarol. 2018, 44, 288–299. [Google Scholar] [CrossRef]
- Clayton, D.H. Host specificity of Strigiphilus owl lice (Ischnocera: Philopteridae), with the description of new species and host associations. J. Med. Entomol. 1990, 27, 257–265. [Google Scholar] [CrossRef]
- Irestedt, M.; Jønsson, K.A.; Fjeldså, J.; Chistidis, L.; Ericson, P.G.P. An unexpectedly long history of sexual selection in birds-of-paradise. BMC Evol. Biol. 2009, 9, 235–242. [Google Scholar] [CrossRef]
- Frith, C.B.; Beehler, B.M. The Birds of Paradise. Bird Families of the World; Oxford University Press: Oxford, NY, USA; Tokyo, Japan, 1998; 613p. [Google Scholar]
- Fuller, E. The Lost Birds of Paradise; Swan Hill Press: Shrewsbury, UK, 1995; 160p. [Google Scholar]
- Frith, C.B.; Frith, D.W. Description of the unique Parotia lawesii x Paradisaea rudolphi hybrid bird of paradise (Aves: Passeriformes: Paradisaeidae). Rec. Aust. Mus. 1996, 48, 111–116. [Google Scholar] [CrossRef]
- Frith, C.B.; Frith, D.W. The unique type specimen of the bird of paradise Lophorina superba pseudoparotia Stresemann 1934 (Paradisaeidae): A hybrid of Lophorina superba × Parotia carolae. J. Fur Ornithol. 1996, 137, 515–521. [Google Scholar] [CrossRef]
- Fuller, E. Hybridization amongst the Paradisaeidae. Bull. Brit. Ornithol. Club 1979, 99, 145–152. [Google Scholar]
- Martin, P.R. The paradox of the Birds-of-Paradise: Persistent hybridization as a signature of historical reinforcement. Ideas Ecol. Evol. 2015, 8, 58–66. [Google Scholar] [CrossRef]
- Mc Charty, E.M. Handbook of Avian Hybrids of the World; Oxford University Press: Oxford, NY, USA; Tokyo, Japan, 2006; 608p. [Google Scholar]
- Koch, A. Discovery of a rare hybrid specimen known as Maria’s bird of paradise at the Staatliches Naturhistorisches Museum in Braunschweig. Zoosyst. Evol. 2018, 94, 315–324. [Google Scholar] [CrossRef]
- Kaszewska-Gilas, K.; Kosicki, J.Z.; Hromada, M.; Skoracki, M. Global studies of the host-parasite relationships between ectoparasitic mites of the family Syringophilidae and birds of the order Columbiformes. Animals 2021, 11, 3392. [Google Scholar] [CrossRef]
- Skoracki, M.; Hromada, M. A review of picobiine mites (Acari: Syringophilidae: Picobiinae) parasitising African birds. Folia Parasitol. 2013, 60, 192–212. [Google Scholar] [CrossRef]
- Jarvis, E.D.; Mirarab, S.; Aberer, A.J.; Li, B.; Houde, P.; Li, C.; Ho, S.Y.; Faircloth, B.C.; Nabholz, B.; Howard, J.T.; et al. Whole-genome analyses resolve early branches in the tree of life of modern birds. Science 2014, 346, 1320–1331. [Google Scholar] [CrossRef]
- Prum, R.O.; Jacob, S.B.; Dornburg, A.; Field, D.J.; Townsend, J.P.; Lemmon, E.M.; Lemmon, A.R. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nat. Lett. 2015, 15697, 569–573. [Google Scholar] [CrossRef]
- Nattress, B. Horizontal transmission of Syringophilopsis kirgizorum (Acari: Cheyletoidea: Syringophilidae). Acarina 2011, 19, 270. [Google Scholar]
- Skoracki, M.; Kosicki, J.Z.; Sikora, B.; Töpfer, T.; Hušek, J.; Unsöld, M.; Hromada, M. The occurrence of quill mites (Arachnida: Acariformes: Syringophilidae) on Bee-eaters (Aves: Coraciiformes: Meropidae: Merops) of two sister clades. Animals 2021, 11, 3500. [Google Scholar] [CrossRef] [PubMed]
- Hromada, M.; Klimovicova, M.; Unsoeld, M.; Skoracki, M. Host-parasite relationships in the system composed by cuckoos and quill mites. Syst. Appl. Acarol. 2016, 21, 528–536. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sikora, B.; Unsoeld, M.; Melzer, R.R.; Friedrich, S.; Skoracki, M. First Records of Picobiine Mites Associated with Birds-of-Paradise: Can Interspecific Sexual Behaviour of Hosts Play a Role in the Distribution of Quill Mite Parasites? Animals 2023, 13, 1509. https://doi.org/10.3390/ani13091509
Sikora B, Unsoeld M, Melzer RR, Friedrich S, Skoracki M. First Records of Picobiine Mites Associated with Birds-of-Paradise: Can Interspecific Sexual Behaviour of Hosts Play a Role in the Distribution of Quill Mite Parasites? Animals. 2023; 13(9):1509. https://doi.org/10.3390/ani13091509
Chicago/Turabian StyleSikora, Bozena, Markus Unsoeld, Roland R. Melzer, Stefan Friedrich, and Maciej Skoracki. 2023. "First Records of Picobiine Mites Associated with Birds-of-Paradise: Can Interspecific Sexual Behaviour of Hosts Play a Role in the Distribution of Quill Mite Parasites?" Animals 13, no. 9: 1509. https://doi.org/10.3390/ani13091509
APA StyleSikora, B., Unsoeld, M., Melzer, R. R., Friedrich, S., & Skoracki, M. (2023). First Records of Picobiine Mites Associated with Birds-of-Paradise: Can Interspecific Sexual Behaviour of Hosts Play a Role in the Distribution of Quill Mite Parasites? Animals, 13(9), 1509. https://doi.org/10.3390/ani13091509